
Hitching a Ride: Examining the Ability of a Specialist Baculovirus to Translocate through Its Insect Host’s Food Plant

 , , , and
, , , and
Abstract
:1. Introduction
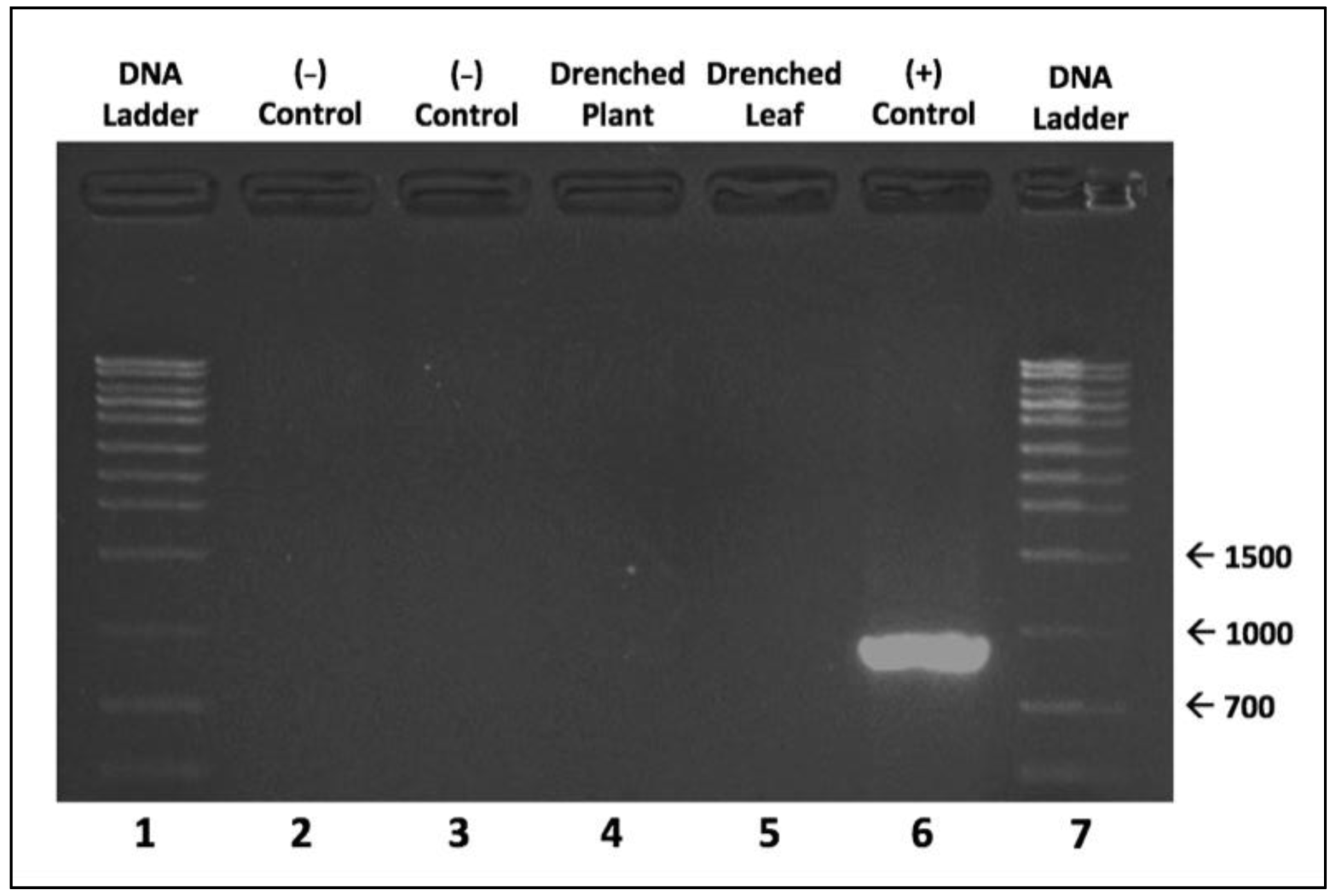
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monnerat, R.G.; Soares, C.M.; Capdeville, G.; Jones, G.; Martins, É.S.; Praça, L.; Cordeiro, B.A.; Braz, S.V.; Dos Santos, R.C.; Berry, C. Translocation and insecticidal activity of Bacillus thuringiensis living inside of plants. Microb. Biotechnol. 2009, 2, 512–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Suárez, R.; Verduzco-Rosas, L.A.; Del Rincón-Castro, M.C.; Délano-Frier, J.P.; Ibarra, J.E. Translocation of Bacillus thuringiensis in Phaseolus vulgaris tissues and vertical transmission in Arab. Thaliana. J. Appl. Microbiol. 2017, 122, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant-endophyte-herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Cory, J.S.; Myers, J.H. The Ecology and Evolution of Insect Baculoviruses. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 239–272. [Google Scholar] [CrossRef] [Green Version]
- Pusztai, M.; Fast, P.; Gringorten, L.; Kaplan, H.; Lessard, T.; Carey, P.R. The mechanism of sunlight-mediated inactivation of Bacillus thuringiensis crystals. Biochem. J. 1991, 273, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruska, A.J.; Gould, F. Fall Armyworm (Lepidoptera: Noctuidae) and Diatraea lineolata (Lepidoptera: Pyralidae): Impact of Larval Population Level and Temporal Occurrence on Maize Yield in Nicaragua. J. Econ. Entomol. 1997, 90, 611–622. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Elderd, B.D. Developing Models of Disease Transmission: Insights from Ecological Studies of Insects and Their Baculoviruses. PLoS Pathog. 2013, 9, e1003372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reagan, B.C.; Burch-Smith, T.M. Viruses Reveal the Secrets of Plasmodesmal Cell Biology. Mol. Plant Microbe Interact. 2020, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, M. Plant virus replication and movement. Virology 2015, 479-480, 657–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, C.; Sandig, V.; Jennings, G.; Rudolph, M.; Schlag, P.; Strauss, M. Efficient gene transfer into human hepatocytes by baculovirus vectors. Proc. Natl. Acad. Sci. USA 1995, 92, 10099–10103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condreay, J.P.; Witherspoon, S.M.; Clay, W.C.; Kost, T.A. Transient and stable gene expression in mammalian cells transduced with a recombinant baculovirus vector. Proc. Natl. Acad. Sci. USA 1999, 96, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elderd, B.D.; Reilly, J.R. Warmer temperatures increase disease transmission and outbreak intensity in a host-pathogen system. J. Anim. Ecol. 2014, 83, 838–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.L.; Puttler, B.; Popham, H.J.R. Genomic sequence analysis of a fast-killing isolate of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 775–790. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 11 November 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Term | Estimate | Standard Error | Z Value | Pr (>|z|) |
|---|---|---|---|---|
| Intercept | 1.447 | 0.556 | 2.604 | 0.009 |
| Negative Control | −39.320 | 1.036 × 107 | 0.000 | 1.000 |
| Drenched Plant | −38.690 | 8.523 × 106 | 0.000 | 1.000 |
| Drenched Leaf | −46.370 | 9.789 × 106 | 0.000 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Issa, P.P.; Garvey, M.; Grimmell, S.; Pantha, P.; Dassanayake, M.; Elderd, B.D. Hitching a Ride: Examining the Ability of a Specialist Baculovirus to Translocate through Its Insect Host’s Food Plant. Pathogens 2021, 10, 1500. https://doi.org/10.3390/pathogens10111500
Issa PP, Garvey M, Grimmell S, Pantha P, Dassanayake M, Elderd BD. Hitching a Ride: Examining the Ability of a Specialist Baculovirus to Translocate through Its Insect Host’s Food Plant. Pathogens. 2021; 10(11):1500. https://doi.org/10.3390/pathogens10111500
Chicago/Turabian StyleIssa, Peter P., Michael Garvey, Scott Grimmell, Pramod Pantha, Maheshi Dassanayake, and Bret D. Elderd. 2021. "Hitching a Ride: Examining the Ability of a Specialist Baculovirus to Translocate through Its Insect Host’s Food Plant" Pathogens 10, no. 11: 1500. https://doi.org/10.3390/pathogens10111500
APA StyleIssa, P. P., Garvey, M., Grimmell, S., Pantha, P., Dassanayake, M., & Elderd, B. D. (2021). Hitching a Ride: Examining the Ability of a Specialist Baculovirus to Translocate through Its Insect Host’s Food Plant. Pathogens, 10(11), 1500. https://doi.org/10.3390/pathogens10111500