
FLO11, a Developmental Gene Conferring Impressive Adaptive Plasticity to the Yeast Saccharomyces cerevisiae


Abstract
:1. Introduction
2. Primary Sequence Analysis Distinguishes Flo11p from the Other Yeast Flocculins
3. Relationship between Sequence/Structure and Role in the Physiological Function of Flo11p
3.1. Strain Phenotypes Are Shaped by the Plasticity of the FLO11-Encoded Protein
3.2. Role of A, B, and C-Domains in the Physiological Function of Flo11p
3.3. Dual Role of the Amyloid-Forming Sequence in Flo11p-Dependent Cell–Cell Interactions
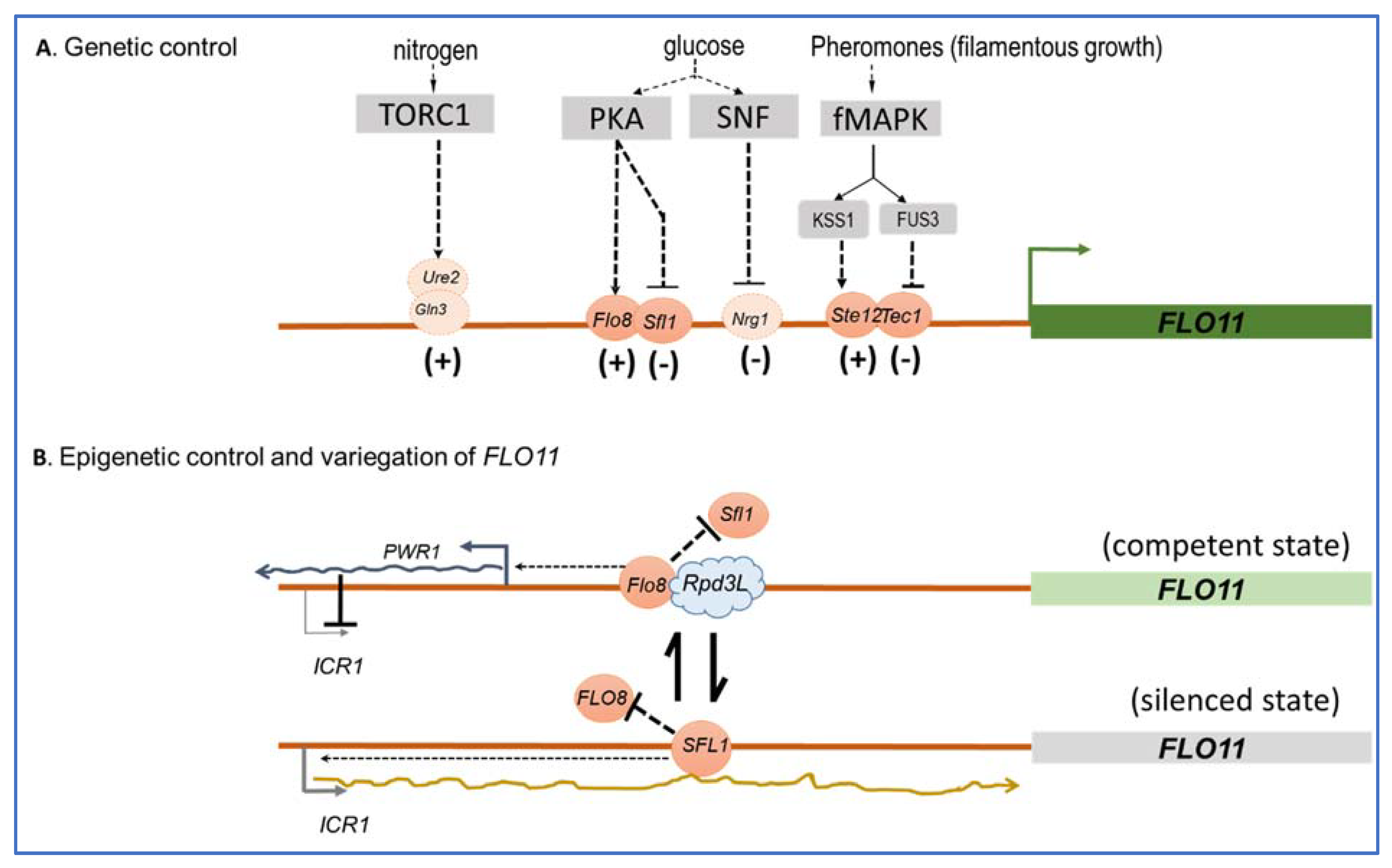
4. Intragenic Repeats Combined with Epigenetic and Conventional Genetic Regulation of FLO11 Confers an Impressive Evolutionary Plasticity to S. cerevisiae to Exploit New Niches and Resources
5. Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansen, E.H. Recherches sur la physiologie et la morphologie des ferments alcooliques. V. Methodes pour obtenior des cultures pures de saccharomyces et de microorganismes analogues. CR Trav. Lab. Carlsb. 1883, 2, 105. [Google Scholar]
- Stratford, M. Yeast flocculation: Reconciliation of physiological and genetic viewpoints. Yeast 1992, 8, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; van den Dool, A.H.; Jacobson, D.; Bauer, F.F. Comparative transcriptomic and proteomic profiling of industrial wine yeast strains. Appl. Environ. Microbiol. 2010, 76, 3911–3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smukalla, S.; Caldara, M.; Pochet, N.; Beauvais, A.; Guadagnini, S.; Yan, C.; Vinces, M.D.; Jansen, A.; Prevost, M.C.; Latge, J.P.; et al. Flo1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 2008, 135, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidalgo, M.; Barrales, R.R.; Ibeas, J.I.; Jimenez, J. Adaptive evolution by mutations in the flo11 gene. Proc. Natl. Acad. Sci. USA 2006, 103, 11228–11233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, H. Flor yeasts of Saccharomyces cerevisiae—Their ecology, genetics and metabolism. Int. J. Food Microbiol. 2013, 167, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, C.J.; Fink, G.R. The logic of cell division in the life cycle of yeast. Science 1992, 257, 626. [Google Scholar] [CrossRef]
- Kron, S.J. Filamentous growth in budding yeast. Trends Microbiol. 1997, 5, 450–454. [Google Scholar] [CrossRef]
- Roberts, R.L.; Fink, G.R. Elements of a single map kinase cascade in saccharomyces cerevisiae mediate two developmental programs in the same cell type: Mating and invasive growth. Genes Dev. 1994, 8, 2974–2985. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O.; Gancedo, J.M. Pseudohyphal growth is induced in Saccharomyces cerevisiae by a combination of stress and camp signalling. Antonie Van Leeuwenhoek 2000, 78, 187–194. [Google Scholar] [CrossRef]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: Regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar] [CrossRef]
- Guo, B.; Styles, C.A.; Feng, Q.; Fink, G.R. A saccharomyces gene family involved in invasive growth, cell-cell adhesion, and mating. Proc. Natl. Acad. Sci. USA 2000, 97, 12158–12163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, T.B. Going with the flo: The role of flo11-dependent and independent interactions in yeast mat formation. J. Fungi 2018, 4, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, T.B.; Fink, G.R. Bakers’ yeast, a model for fungal biofilm formation. Science 2001, 291, 878–881. [Google Scholar] [CrossRef]
- Bruckner, S.; Mosch, H.U. Choosing the right lifestyle: Adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2012, 36, 25–58. [Google Scholar] [CrossRef] [Green Version]
- Blankenship, J.R.; Mitchell, A.P. How to build a biofilm: A fungal perspective. Curr. Opin. Microbiol. 2006, 9, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Dranginis, A.M.; Rauceo, J.M.; Coronado, J.E.; Lipke, P.N. A biochemical guide to yeast adhesins: Glycoproteins for social and antisocial occasions. Microbiol. Mol. Biol. Rev. 2007, 71, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [Green Version]
- Lemesle-Varloot, L.; Henrissat, B.; Gaboriaud, C.; Bissery, V.; Morgat, A.; Mornon, J.P. Hydrophobic cluster analysis: Procedures to derive structural and functional information from 2-d-representation of protein sequences. Biochimie 1990, 72, 555–574. [Google Scholar] [CrossRef]
- Goossens, K.V.; Stassen, C.; Stals, I.; Donohue, D.S.; Devreese, B.; De Greve, H.; Willaert, R.G. The n-terminal domain of the flo1 flocculation protein from saccharomyces cerevisiae binds specifically to mannose carbohydrates. Eukaryot. Cell 2011, 10, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Veelders, M.; Bruckner, S.; Ott, D.; Unverzagt, C.; Mosch, H.U.; Essen, L.O. Structural basis of flocculin-mediated social behavior in yeast. Proc. Natl. Acad. Sci. USA 2010, 107, 22511–22516. [Google Scholar] [CrossRef] [Green Version]
- Willaert, R.G. Adhesins of yeasts: Protein structure and interactions. J. Fungi 2018, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Kraushaar, T.; Bruckner, S.; Veelders, M.; Rhinow, D.; Schreiner, F.; Birke, R.; Pagenstecher, A.; Mosch, H.U.; Essen, L.O. Interactions by the fungal flo11 adhesin depend on a fibronectin type iii-like adhesin domain girdled by aromatic bands. Structure 2015, 23, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P. Description of komagataella phaffii sp. Nov. And the transfer of Pichia pseudopastoris to the methylotrophic yeast genus komagataella. Int. J. Syst Evol. Microbiol. 2005, 55, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Essen, L.O.; Vogt, M.S.; Mosch, H.U. Diversity of GPI-anchored fungal adhesins. Biol. Chem. 2020, 401, 1389–1405. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Escamilla, A.M.; Rousseau, F.; Schymkowitz, J.; Serrano, L. Prediction of sequence-dependent and mutational effects on the aggregation of peptides and proteins. Nat. Biotechnol. 2004, 22, 1302–1306. [Google Scholar] [CrossRef] [PubMed]
- Ramsook, C.B.; Tan, C.; Garcia, M.C.; Fung, R.; Soybelman, G.; Henry, R.; Litewka, A.; O’Meally, S.; Otoo, H.N.; Khalaf, R.A.; et al. Yeast cell adhesion molecules have functional amyloid-forming sequences. Eukaryot. Cell 2010, 9, 393–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, L.M.; Li, L.; Yang, Y.; Dranginis, A.M. Expression and characterization of the flocculin flo11/muc1, a Saccharomyces cerevisiae mannoprotein with homotypic properties of adhesion. Eukaryot. Cell 2007, 6, 2214–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.X.; El-Kirat-Chatel, S.; Joseph, I.G.; Jackson, D.N.; Ramsook, C.B.; Dufrene, Y.F.; Lipke, P.N. Force sensitivity in saccharomyces cerevisiae flocculins. mSphere 2016, 1, e00128-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, W.S.; Dranginis, A.M. Flo11, a yeast gene related to the sta genes, encodes a novel cell surface flocculin. J. Bacteriol. 1996, 178, 7144–7151. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, M.G.; Bauer, F.F.; Marmur, J.; Pretorius, I.S. Muc1, a mucin-like protein that is regulated by mss10, is critical for pseudohyphal differentiation in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 8419–8424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretorius, I.S.; Lambrechts, M.G.; Marmur, J. The glucoamylase multigene family in Saccharomyces cerevisiae var. Diastaticus: An overview. Crit. Rev. Biochem. Mol. Biol. 1991, 26, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, M.; Nakagawa, Y.; Hayakawa, M.; Iimura, Y. Flo11 is the primary factor in flor formation caused by cell surface hydrophobicity in wild-type flor yeast. Biosci. Biotechnol. Biochem. 2006, 70, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Zara, S.; Bakalinsky, A.T.; Zara, G.; Pirino, G.; Demontis, M.A.; Budroni, M. Flo11-based model for air-liquid interfacial biofilm formation by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 2934–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voordeckers, K.; De Maeyer, D.; van der Zande, E.; Vinces, M.D.; Meert, W.; Cloots, L.; Ryan, O.; Marchal, K.; Verstrepen, K.J. Identification of a complex genetic network underlying Saccharomyces cerevisiae colony morphology. Mol. Microbiol. 2012, 86, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Bouyx, C.; Schiavone, M.; Teste, M.A.; Dague, E.; Sieczkowski, N.; Julien, A.; Francois, J.M. The dual role of amyloid-beta-sheet sequences in the cell surface properties of flo11-encoded flocculins in Saccharomyces cerevisiae. elife 2021, 10, e68592. [Google Scholar] [CrossRef]
- Purevdorj-Gage, B.; Orr, M.E.; Stoodley, P.; Sheehan, K.B.; Hyman, L.E. The role of flo11 in Saccharomyces cerevisiae biofilm development in a laboratory based flow-cell system. FEMS Yeast Res. 2007, 7, 372–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mulders, S.E.; Christianen, E.; Saerens, S.M.; Daenen, L.; Verbelen, P.J.; Willaert, R.; Verstrepen, K.J.; Delvaux, F.R. Phenotypic diversity of flo protein family-mediated adhesion in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 178–190. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Styles, C.A.; Fink, G.R. Saccharomyces cerevisiae s288c has a mutation in flo8, a gene required for filamentous growth. Genetics 1996, 144, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, T.B.; Jansen, A.; Peng, X.; Fink, G.R. Mat formation in Saccharomyces cerevisiae requires nutrient and ph gradients. Eukaryot. Cell 2008, 7, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Bayly, J.C.; Douglas, L.M.; Pretorius, I.S.; Bauer, F.F.; Dranginis, A.M. Characteristics of flo11-dependent flocculation in saccharomyces cerevisiae. FEMS Yeast Res. 2005, 5, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barua, S.; Li, L.; Lipke, P.N.; Dranginis, A.M. Molecular basis for strain variation in the saccharomyces cerevisiae adhesin flo11p. mSphere 2016, 1, e00129-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, A.; Forment, J.; Serrano, R. Involvement of nst1p/ynl091w and msl1p, a u2b” splicing factor, in Saccharomyces cerevisiae salt tolerance. Yeast 2002, 19, 193–202. [Google Scholar] [CrossRef]
- Bruckner, S.; Schubert, R.; Kraushaar, T.; Hartmann, R.; Hoffmann, D.; Jelli, E.; Drescher, K.; Muller, D.J.; Oliver Essen, L.; Mosch, H.U. Kin discrimination in social yeast is mediated by cell surface receptors of the flo11 adhesin family. elife 2020, 9, e55587. [Google Scholar] [CrossRef]
- Goossens, K.V.; Willaert, R.G. The n-terminal domain of the flo11 protein from Saccharomyces cerevisiae is an adhesin without mannose-binding activity. FEMS Yeast Res. 2012, 12, 78–87. [Google Scholar] [CrossRef] [Green Version]
- West, S.A.; Griffin, A.S.; Gardner, A.; Diggle, S.P. Social evolution theory for microorganisms. Nat. Rev. Microbiol. 2006, 4, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caro, L.H.; Tettelin, H.; Vossen, J.H.; Ram, A.F.; van den, E.H.; Klis, F.M. In silicio identification of glycosyl-phosphatidylinositol-anchored plasma-membrane and cell wall proteins of Saccharomyces cerevisiae. Yeast 1997, 13, 1477–1489. [Google Scholar] [CrossRef]
- Kitagaki, H.; Wu, H.; Shimoi, H.; Ito, K. Two homologous genes, dcw1 (ykl046c) and dfg5, are essential for cell growth and encode glycosylphosphatidylinositol (GPI)-anchored membrane proteins required for cell wall biogenesis in Saccharomyces cerevisiae. Mol. Microbiol. 2002, 46, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.S.; Schmitz, G.F.; Varon Silva, D.; Mosch, H.U.; Essen, L.O. Structural base for the transfer of GPI-anchored glycoproteins into fungal cell walls. Proc. Natl. Acad. Sci. USA 2020, 117, 22061–22067. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M. Review: Cell wall assembly in yeast. Yeast 1994, 10, 851–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieman, M.B.; Cormack, B.P. Multiple sequence signals determine the distribution of glycosylphosphatidylinositol proteins between the plasma membrane and cell wall in Saccharomyces cerevisiae. Microbiology 2004, 150, 3105–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunanithi, S.; Vadaie, N.; Chavel, C.A.; Birkaya, B.; Joshi, J.; Grell, L.; Cullen, P.J. Shedding of the mucin-like flocculin flo11p reveals a new aspect of fungal adhesion regulation. Curr. Biol. 2010, 20, 1389–1395. [Google Scholar] [CrossRef] [Green Version]
- Verbelen, P.J.; Dekoninck, T.M.; Saerens, S.M.; Van Mulders, S.E.; Thevelein, J.M.; Delvaux, F.R. Impact of pitching rate on yeast fermentation performance and beer flavour. Appl. Microbiol. Biotechnol. 2009, 82, 155–167. [Google Scholar] [CrossRef]
- Fidalgo, M.; Barrales, R.R.; Jimenez, J. Coding repeat instability in the flo11 gene of saccharomyces yeasts. Yeast 2008, 25, 879–889. [Google Scholar] [CrossRef]
- Meem, M.H.; Cullen, P.J. The impact of protein glycosylation on flo11-dependent adherence in saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipke, P.N.; Garcia, M.C.; Alsteens, D.; Ramsook, C.B.; Klotz, S.A.; Dufrene, Y.F. Strengthening relationships: Amyloids create adhesion nanodomains in yeasts. Trends Microbiol. 2012, 20, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Lipke, P.N.; Ovalle, R. Cell wall architecture in yeast: New structure and new challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.C.; Lee, J.T.; Ramsook, C.B.; Alsteens, D.; Dufrene, Y.F.; Lipke, P.N. A role for amyloid in cell aggregation and biofilm formation. PLoS ONE 2011, 6, e17632. [Google Scholar] [CrossRef]
- Chan, C.X.; Lipke, P.N. Role of force-sensitive amyloid-like interactions in fungal catch bonding and biofilms. Eukaryot. Cell 2014, 13, 1136–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsteens, D.; Garcia, M.C.; Lipke, P.N.; Dufrene, Y.F. Force-induced formation and propagation of adhesion nanodomains in living fungal cells. Proc. Natl. Acad. Sci. USA 2010, 107, 20744–20749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formosa, C.; Schiavone, M.; Boisrame, A.; Richard, M.L.; Duval, R.E.; Dague, E. Multiparametric imaging of adhesive nanodomains at the surface of Candida albicans by atomic force microscopy. Nanomedicine 2015, 11, 57–65. [Google Scholar] [CrossRef] [Green Version]
- de Groot, P.W.; Bader, O.; de Boer, A.D.; Weig, M.; Chauhan, N. Adhesins in human fungal pathogens: Glue with plenty of stick. Eukaryot. Cell 2013, 12, 470–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, V.; Herman-Bausier, P.; Shaw, C.; Conrad, K.A.; Garcia-Sherman, M.C.; Draghi, J.; Dufrene, Y.F.; Lipke, P.N.; Rauceo, J.M. An amyloid core sequence in the major candida albicans adhesin als1p mediates cell-cell adhesion. mBio 2019, 10, e01766-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipke, P.N.; Mathelie-Guinlet, M.; Viljoen, A.; Dufrene, Y.F. A new function for amyloid-like interactions: Cross-beta aggregates of adhesins form cell-to-cell bonds. Pathogens 2021, 10, 1013. [Google Scholar] [CrossRef]
- Schiavone, M.; Sieczkowski, N.; Castex, M.; Dague, E.; Marie Francois, J. Effects of the strain background and autolysis process on the composition and biophysical properties of the cell wall from two different industrial yeasts. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Dehullu, J.; Valotteau, C.; Herman-Bausier, P.; Garcia-Sherman, M.; Mittelviefhaus, M.; Vorholt, J.A.; Lipke, P.N.; Dufrene, Y.F. Fluidic force microscopy demonstrates that homophilic adhesion by candida albicans ALS proteins is mediated by amyloid bonds between cells. Nano Lett. 2019, 19, 3846–3853. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Jansen, A.; Lewitter, F.; Fink, G.R. Intragenic tandem repeats generate functional variability. Nat. Genet. 2005, 37, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Rupp, S.; Summers, E.; Lo, H.J.; Madhani, H.; Fink, G. Map kinase and camp filamentation signaling pathways converge on the unusually large promoter of the yeast flo11 gene. EMBO J. 1999, 18, 1257–1269. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.; Starr, I.; Jamalzadeh, S.; Muniz, O.; Kumar, A.; Gokcumen, O.; Ferkey, D.M.; Cullen, P.J. Filamentation regulatory pathways control adhesion-dependent surface responses in yeast. Genetics 2019, 212, 667–690. [Google Scholar] [CrossRef] [PubMed]
- Pfyffer, G.E.; Rast, D.M. Accumulation of acyclic polyols and trehalose as related to growth form and carbohydrate source in the dimorphic fungi Mucor rouxii and Candida albicans. Mycopathologia 1989, 105, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.J.; Sprague, G.F., Jr. Glucose depletion causes haploid invasive growth in yeast. Proc. Natl. Acad. Sci. USA 2000, 97, 13619–13624. [Google Scholar] [CrossRef] [Green Version]
- Kuchin, S.; Vyas, V.K.; Carlson, M. Snf1 protein kinase and the repressors nrg1 and nrg2 regulate flo11, haploid invasive growth, and diploid pseudohyphal differentiation. Mol. Cell Biol. 2002, 22, 3994–4000. [Google Scholar] [CrossRef] [Green Version]
- Cutler, N.S.; Pan, X.; Heitman, J.; Cardenas, M.E. The tor signal transduction cascade controls cellular differentiation in response to nutrients. Mol. Biol. Cell 2001, 12, 4103–4113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, T.M.; Xu, W.; Diamond, A.; Mitchell, A.P. Alkaline response genes of Saccharomyces cerevisiae and their relationship to the rim101 pathway. J. Biol. Chem. 2001, 276, 1850–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penalva, M.A.; Tilburn, J.; Bignell, E.; Arst, H.N., Jr. Ambient PH gene regulation in fungi: Making connections. Trends Microbiol. 2008, 16, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halme, A.; Bumgarner, S.; Styles, C.; Fink, G.R. Genetic and epigenetic regulation of the FLO gene family generates cell-surface variation in yeast. Cell 2004, 116, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Barrales, R.R.; Jimenez, J.; Ibeas, J.I. Identification of novel activation mechanisms for flo11 regulation in saccharomyces cerevisiae. Genetics 2008, 178, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Octavio, L.M.; Gedeon, K.; Maheshri, N. Epigenetic and conventional regulation is distributed among activators of flo11 allowing tuning of population-level heterogeneity in its expression. PLoS Genet. 2009, 5, e1000673. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, H.; Shaban, K.; Foster, B.; Proteau, Y.; Yankulov, K. Histone chaperones and the rrm3p helicase regulate flocculation in S. cerevisiae. Epigenetics Chromatin 2019, 12, 56. [Google Scholar] [CrossRef]
- Bumgarner, S.L.; Dowell, R.D.; Grisafi, P.; Gifford, D.K.; Fink, G.R. Toggle involving cis-interfering noncoding RNAs controls variegated gene expression in yeast. Proc. Natl. Acad. Sci. USA 2009, 106, 18321–18326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coi, A.L.; Bigey, F.; Mallet, S.; Marsit, S.; Zara, G.; Gladieux, P.; Galeote, V.; Budroni, M.; Dequin, S.; Legras, J.L. Genomic signatures of adaptation to wine biological ageing conditions in biofilm-forming flor yeasts. Mol. Ecol. 2017, 26, 2150–2166. [Google Scholar] [CrossRef] [PubMed]
- Fay, J.C.; McCullough, H.L.; Sniegowski, P.D.; Eisen, M.B. Population genetic variation in gene expression is associated with phenotypic variation in saccharomyces cerevisiae. Genome Biol. 2004, 5, R26. [Google Scholar] [CrossRef] [Green Version]
- Klingberg, T.D.; Lesnik, U.; Arneborg, N.; Raspor, P.; Jespersen, L. Comparison of Saccharomyces cerevisiae strains of clinical and nonclinical origin by molecular typing and determination of putative virulence traits. FEMS Yeast Res. 2008, 8, 631–640. [Google Scholar] [CrossRef] [Green Version]
- McCusker, J.H.; Clemons, K.V.; Stevens, D.A.; Davis, R.W. Genetic characterization of pathogenic Saccharomyces cerevisiae isolates. Genetics 1994, 136, 1261–1269. [Google Scholar] [CrossRef]
- de Llanos, R.; Fernandez-Espinar, M.T.; Querol, A. A comparison of clinical and food Saccharomyces cerevisiae isolates on the basis of potential virulence factors. Antonie Van Leeuwenhoek 2006, 90, 221–231. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Flo1p | Flo5p | Flo9p | Flo10p | Flo11p | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | AA Position | Motif | β-Aggregate (%) | AA Position | Motif | β-Aggregate (%) | AA Position | Motif | β-Aggregate (%) | AA Position | Motif | β-Aggregate (%) | AA Position | Motif | Mean β-Aggregate (%) |
| A | 7–21 | YMFLAVFTLLALTSV | 82.2 | 9–22 | IFVILAFLALINVA | 82 | 6–22 | YYCLLLAIVLLGLTNVV | 77.6 | 7–23 | YIFLTGLFLLSVANVAL | 81.7 | 5–16 | FLLAYVLSLLF | 96 |
| B | 308–312 | TVIVI | 87.9 | 207–215 | TVYMYAGYY | 30.5 | 118–122 | IIAYW | 72 | ||||||
| 353–357 | TIIVI | 87.3 | 308–312 | TVIVI | 73.2 | 207–215 | TVYMYAGFY | 50.9 | |||||||
| 398–402 | TIIVI | 87.3 | 353–357 | TVIVI | 87.8 | 308–312 | TVIVI | 87.9 | |||||||
| 443–447 | TIIVI | 87.3 | 398–402 | TVIVI | 87.8 | 353–357 | TIIVI | 87.3 | |||||||
| 488–492 | TIIVI | 87.3 | 443–447 | TVIVI | 87.8 | 398–402 | TIIVI | 87.3 | |||||||
| 533–537 | TIIVI | 87.2 | 488–492 | TVIVI | 87.8 | 443–447 | TIIVI | 87.3 | |||||||
| 578–562 | TIIVI | 87.2 | 533–537 | TVIVI | 87.9 | 498–492 | TIIVI | 87.3 | |||||||
| 623–627 | TIIVI | 87.2 | 578–582 | TVIVI | 87.9 | 533–537 | TIIVI | 87.2 | |||||||
| 667–672 | TIIVI | 87.2 | 623–627 | TVIVI | 87.9 | 578–582 | TIIVI | 87.2 | |||||||
| 713–717 | TIIVI | 87.2 | 783–788 | TLVTVT | 31.7 | 623–627 | TIIVI | 87.2 | |||||||
| 758–762 | TVIVI | 87.8 | 802–811 | AIVSTATVTV | 45.2 | 668–672 | TIIVI | 87.2 | |||||||
| 803–807 | TVIVI | 87.8 | 855–859 | TVVTI | 37.2 | 713–717 | TIIVI | 87.2 | |||||||
| 848–852 | TVIVI | 87.8 | 758–762 | TVIVI | 87.8 | ||||||||||
| 893–897 | TVIVI | 87.8 | 803–807 | TVIVI | 87.8 | ||||||||||
| 938–942 | TVIVI | 87.8 | 848–852 | TVIIV | 87.,7 | ||||||||||
| 983–987 | TVIVI | 87.8 | 1021–1026 | TLVTVT | 31.9 | ||||||||||
| C | 1028–1032 | TVIVI | 87.9 | 1040–1049 | AIVSTATVTV | 42.6 | |||||||||
| 1073–1077 | TVIVV | 88.5 | 1093–1097 | TVVTI | 37.4 | ||||||||||
| 1234–1239 | TLVTVT | 31.7 | 1028–1035 | VVTVYSTW | 48.5 | 1033–1042 | VTTVVSTTVV | 75.8 | |||||||
| 1254–1262 | IVSTATVTV | 45.9 | 1115–1119 | VLISV | 47 | 1056–1061 | ITTTFV | 56 | |||||||
| 1299–1303 | TVVTI | 37.2 | 906–911 | TLVTVT | 36.1 | 1144–1149 | TLVTVT | 37.7 | 1157–1168 | ISIFIASLLLAI | 89.8 | 1133–1144 | TLVTTAVTTTVV | 84.8 | |
| 1350–1355 | TLVTVT | 36 | 1063–1074 | LSVFIASLLLAI | 87 | 1175–1182 | VVTVYSTW | 82.6 | 1356–1362 | FMWLLLA | 85.3 | ||||
| 1525–1536 | LSVFIASLLLAI | 87 | 1310–1321 | LSVFIASLLLAI | 87 | ||||||||||
| Strain | ORF (bp) | Repetition Length (TR) | Score | Count | Repetition Start (nt. seq.) | Repetition Stop (nt. seq.) | Repetition Conservation (%) | Repeated Sequence (Consensus) |
|---|---|---|---|---|---|---|---|---|
| S288c | 4104 | 63 | 304 | 24 | 688 | 2388 | 60.8 | tctactacagcaaccacttcaaccaccgcaactactgcaaccacttctactactgaaaccact |
| 33 | 55 | 8 | 2832 | 3095 | 66.7 | ctctgcatgaacaaccactaccactacaactac | ||
| 45 | 48 | 3 | 2429 | 2563 | 84.4 | caaccccatcaagctctagcactgaaagctcttctgctccagtat | ||
| 72 | 24 | 4 | 3099 | 3386 | 66.7 | aactacagttttctccccaaacactgttactactacggtttcttctacaactacaactggtgcagacactac | ||
| Σ1278b | 3633 | 81 | 438 | 13 | 767 | 1819 | 74.6 | caaccagctctaccactgaaagctcttctgctccagctccaactccaaccagctctaccactgaaagctcttctgctccag |
| 45 | 94 | 5 | 1923 | 2147 | 80.9 | cactgaaagctcttctgctccagtaccaactccatccagctctag | ||
| 45 | 46 | 3 | 2158 | 2292 | 83.7 | ccagtaccaactccatccagctctagcactgaaagctcctctgct | ||
| 45 | 29 | 2 | 335 | 424 | 91.1 | gttgcgacgaaaatacctatttgattgacaacccaactgatttca | ||
| 133d | 4890 | 81 | 1708 | 49 | 827 | 4795 | 72.5 | cttcttctgctccagttacttcttctactactgaatcttcttctgctccagctcctactccttcttcttctactactgaat |
| L69 | 5166 | 63 | 323 | 30 | 658 | 2547 | 60.2 | acttcatctaccgctactactgcaaccacttctactactgcaaccacttctactactgcaaca |
| 45 | 95 | 4 | 2646 | 2825 | 88.9 | accagctccaactccatccagctctactactgaaagctcttctgc | ||
| 45 | 71 | 4 | 2958 | 3137 | 82.2 | atccagctctaccactgaaagctcttctgctccagtatcaacccc | ||
| 12 | 41 | 8 | 2836 | 2955 | 73.3 | agctctactgctcca |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouyx, C.; Schiavone, M.; François, J.M. FLO11, a Developmental Gene Conferring Impressive Adaptive Plasticity to the Yeast Saccharomyces cerevisiae. Pathogens 2021, 10, 1509. https://doi.org/10.3390/pathogens10111509
Bouyx C, Schiavone M, François JM. FLO11, a Developmental Gene Conferring Impressive Adaptive Plasticity to the Yeast Saccharomyces cerevisiae. Pathogens. 2021; 10(11):1509. https://doi.org/10.3390/pathogens10111509
Chicago/Turabian StyleBouyx, Clara, Marion Schiavone, and Jean Marie François. 2021. "FLO11, a Developmental Gene Conferring Impressive Adaptive Plasticity to the Yeast Saccharomyces cerevisiae" Pathogens 10, no. 11: 1509. https://doi.org/10.3390/pathogens10111509
APA StyleBouyx, C., Schiavone, M., & François, J. M. (2021). FLO11, a Developmental Gene Conferring Impressive Adaptive Plasticity to the Yeast Saccharomyces cerevisiae. Pathogens, 10(11), 1509. https://doi.org/10.3390/pathogens10111509