
Community Parameters and Genome-Wide RAD-Seq Loci of Ceratothoa oestroides Imply Its Transfer between Farmed European Sea Bass and Wild Farm-Aggregating Fish

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Parasite Epidemiology
2.2. ddRAD Sequencing Output
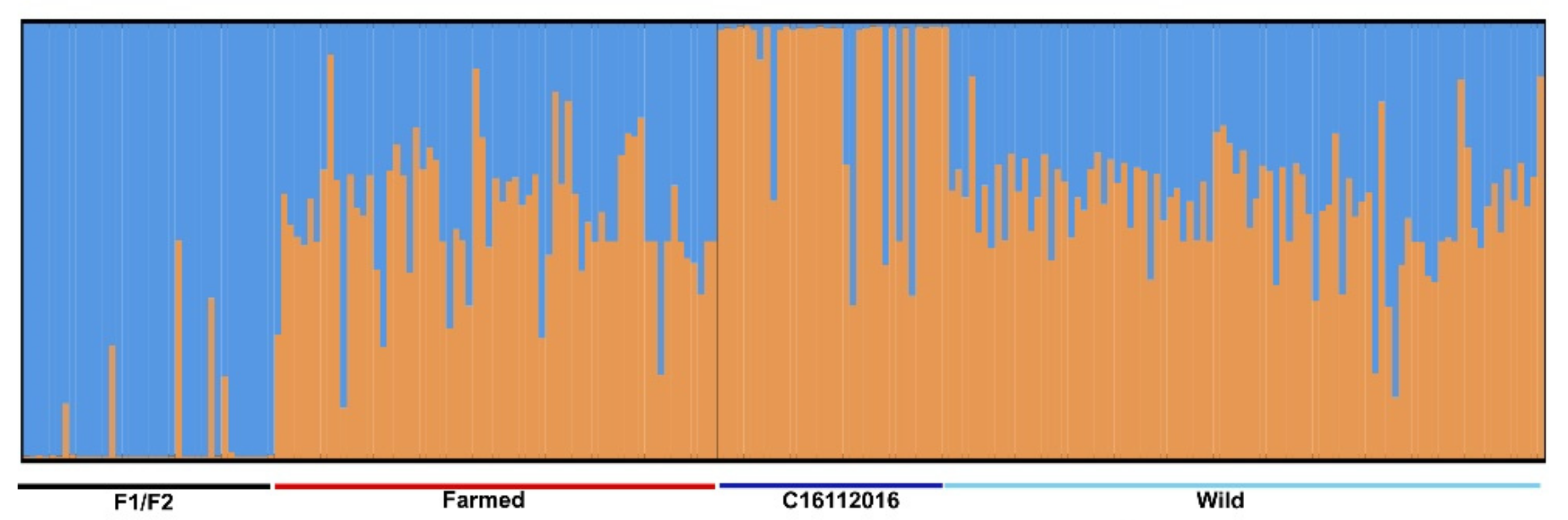
2.3. Population Genetic Structure and Relatedness in C. oestroides
3. Discussion
4. Materials and Methods
4.1. Parasite Sampling and Collection of Epidemiological Data
4.2. Development of C. oestroides Filial Generations
4.3. DNA Isolation and Construction of Double-Digest Restriction-Site-Associated DNA (ddRAD) Libraries
4.4. Data Analysis of ddRAD Library
4.5. C. oestroides Genetic Structure
4.6. Data Deposition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fioravanti, M.L.; Mladineo, I.; Palenzuela, O.; Beraldo, P.; Massimo, M.; Gustinelli, A.; Sitjà-Bobadilla, A. Guide 4—Fish Farmer’s Guide to Combating Parasitic Infections in European Sea Bass and Gilthead Sea Bream Aquaculture; 833-20-104-5; A Series of ParaFishControl Guides to Combating Fish Parasite Infections in Aquaculture; Sitjà-Bobadilla, A., Bello-Gómez, E., Eds.; Springer: Cham, Switzerland, 2020; p. 29. [Google Scholar]
- Romestand, B.; Thuet, P.; Trilles, J.-P. Quelques aspects des mécanismes nutritionnels chez l’isopode cymothoidae: Ceratothoa oestroides (Risso, 1826). Ann. Parasitol. Hum. Comparée 1982, 57, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Nagler, C.; Haug, J.T. Functional morphology of parasitic isopods: Understanding morphological adaptations of attachment and feeding structures in Nerocila as a pre-requisite for reconstructing the evolution of Cymothoidae. Peer J. 2016, 4, e2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittington, I.D.; Chisholm, L.A. Chapter 13: Diseases caused by Monogenea. In Fish Diseases; Eiras, J.C., Segner, H., Wahli, T., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2008; pp. 683–816. [Google Scholar]
- Charfi-Cheikhrouha, F.; Zghidi, W.; Ould Yarba, L.; Trilles, J.P. Les Cymothoidae (Isopodes parasites de poissons) des côtes tunisiennes: Ecologie et indices parasitologiques. Syst. Parasitol. 2000, 46, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Trilles, J.-P.; Radujković, B.; Romestand, B. Parasites des poissons marins du Monténégro: Isopodes. Acta Adriat. 1989, 30, 279–306. [Google Scholar]
- Arechavala-Lopez, P.; Fernandez-Jover, D.; Black, K.D.; Ladoukakis, E.; Bayle-Sempere, J.T.; Sanchez-Jerez, P.; Dempster, T. Differentiating the wild or farmed origin of Mediterranean fish: A review of tools for sea bream and sea bass. Rev. Aquac. 2013, 5, 137–157. [Google Scholar] [CrossRef]
- Peeler, E.; Thrush, M.; Paisley, L.; Rodgers, C. An assessment of the risk of spreading the fish parasite Gyrodactylus salaris to uninfected territories in the European Union with the movement of live Atlantic salmon (Salmo salar) from coastal waters. Aquaculture 2006, 258, 187–197. [Google Scholar] [CrossRef]
- Mladineo, I.; Šegvić, T.; Grubišić, L. Molecular evidence for the lack of transmission of the monogenean Sparicotyle chrysophrii (Monogenea, Polyopisthocotylea) and isopod Ceratothoa oestroides (Crustacea, Cymothoidae) between wild bogue (Boops boops) and cage-reared sea b. Aquaculture 2009, 295, 160–167. [Google Scholar] [CrossRef]
- Mladineo, I.; Šegvić-Bubić, T.; Stanić, R.; Desdevises, Y. Morphological plasticity and phylogeny in a monogenean parasite transferring between wild and reared fish populations. PLoS ONE 2013, 8, e62011. [Google Scholar] [CrossRef] [Green Version]
- Davey, J.W.; Hohenlohe, P.A.; Etter, P.D.; Boone, J.Q.; Catchen, J.M.; Blaxter, M.L. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet. 2011, 12, 499–510. [Google Scholar] [CrossRef]
- Hendricks, S.; Anderson, E.C.; Antao, T.; Bernatchez, L.; Forester, B.R.; Garner, B.; Hand, B.K.; Hohenlohe, P.A.; Kardos, M.; Koop, B.; et al. Recent advances in conservation and population genomics data analysis. Evol. Appl. 2018, 11, 1197–1211. [Google Scholar] [CrossRef]
- Robledo, D.; Palaiokostas, C.; Bargelloni, L.; Martínez, P.; Houston, R. Applications of genotyping by sequencing in aquaculture breeding and genetics. Rev. Aquac. 2018, 10, 670–682. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Meyer, E.; Mckay, J.K.; Matz, M.V. 2b-RAD: A simple and flexible method for genome-wide genotyping. Nat. Methods 2012, 9, 808–810. [Google Scholar] [CrossRef] [PubMed]
- Pante, E.; Abdelkrim, J.; Viricel, A.; Gey, D.; France, S.C.; Boisselier, M.C.; Samadi, S. Use of RAD sequencing for delimiting species. Heredity 2015, 114, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavey, S.A.; Gaudin, J.; Normandeau, E.; Dionne, M.; Castonguay, M.; Audet, C.; Bernatchez, L. RAD sequencing highlights polygenic discrimination of habitat ecotypes in the panmictic american eel. Curr. Biol. 2015, 25, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Crawford, D.L.; Oleksiak, M.F. Ecological population genomics in the marine environment. Brief. Funct. Genom. 2016, 15, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, J.B.; Payton, A.C.; Bjorndal, K.A.; Bolten, A.B.; McDaniel, S.F. Hitchhiking the high seas: Global genomics of rafting crabs. Ecol. Evol. 2019, 9, 957–974. [Google Scholar] [CrossRef] [Green Version]
- Vendrami, D.L.J.; de Noia, M.; Telesca, L.; Handal, W.; Charrier, G.; Boudry, P.; Eberhart-Phillips, L.; Hoffman, J.I. RAD sequencing sheds new light on the genetic structure and local adaptation of European scallops and resolves their demographic histories. Sci. Rep. 2019, 9, 7455. [Google Scholar] [CrossRef]
- Hupalo, D.N.; Bradic, M.; Carlton, J.M. The impact of genomics on population genetics of parasitic diseases. Curr. Opin. Microbiol. 2015, 23, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Barría, A.; Christensen, K.A.; Yoshida, G.M.; Correa, K.; Jedlicki, A.; Lhorente, J.P.; Davidson, W.S.; Yáñez, J.M. Genomic predictions and genome-wide association study of resistance against Piscirickettsia salmonis in coho salmon (Oncorhynchus kisutch) using ddRAD sequencing. G3 Genes Genomes Genet. 2018, 8, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Barría, A.; Marín-Nahuelpi, R.; Cáceres, P.; López, M.E.; Bassini, L.N.; Lhorente, J.P.; Yáñez, J.M. Single-step genome-wide association study for resistance to Piscirickettsia salmonis in rainbow trout (Oncorhynchus mykiss). G3 Genes Genomes Genet. 2019, 9, 3833–3841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, S.N.; Bekaert, M.; Taggart, J.B.; Christie, H.R.L.; Bassett, D.I.; Bron, J.E.; Skuce, P.J.; Gharbi, K.; Skern-Mauritzen, R.; Sturm, A. Identification of a sex-linked SNP marker in the salmon louse (Lepeophtheirus salmonis) using RAD sequencing. PLoS ONE 2013, 8, e77832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, A.; de Noia, M.; Praebel, K.; Kanstad-Hanssen, Ø.; Paterno, M.; Jackson, D.; McGinnity, P.; Sturm, A.; Elmer, K.R.; Llewellyn, M.S. Genetic fingerprinting of salmon louse (Lepeophtheirus salmonis) populations in the North-East Atlantic using a random forest classification approach. Sci. Rep. 2018, 8, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagianou, S.; Athanassopoulou, F.; Ragias, V.; di Cave, D.; Leontides, L.; Golomazou, E. Prevalence and pathology of ectoparasites of mediterranean sea bream and sea bass reared under different environmental and aquaculture conditions. Isr. J. Aquac. Bamidgeh 2006, 58, 78–88. [Google Scholar]
- Horton, T.; Okamura, B. Post-haemorrhagic anaemia in sea bass, Dicentrarchus labrax (L.), caused by blood feeding of Ceratothoa oestroides (Isopoda: Cymothoidae). J. Fish Dis. 2003, 26, 401–406. [Google Scholar] [CrossRef]
- Šarušić, G. Preliminary report of infestation by isopod Ceratothoa oestroides (Risso, 1826), in marine cultured fis. Bull. Eur. Assoc. Fish Pathol. 1999, 19, 110–112. [Google Scholar]
- Horton, T.; Okamura, B. Cymothoid isopod parasites in aquaculture: A review and case study of a Turkish sea bass (Dicentrarchus labrax) and sea bream (Sparus auratus) farm. Dis. Aquat. Organ. 2001, 46, 181–188. [Google Scholar] [CrossRef]
- Mladineo, I.; Hrabar, J.; Vidjak, O.; Bočina, I.; Čolak, S.; Katharios, P.; Cascarano, M.C.; Keklikoglou, K.; Volpatti, D.; Beraldo, P. Host-parasite interaction between parasitic cymothoid Ceratothoa oestroides and its host, farmed European sea bass (Dicentrarchus labrax). Pathogens 2020, 9, 230. [Google Scholar] [CrossRef] [Green Version]
- Čolak, S.; Kolega, M.; Mejdandžić, D.; Župan, I.; Šarić, T.; Piplović, E.; Mustać, B. Prevalence and effects of the cymothoid isopod (Ceratothoa oestroides, Risso 1816) on cultured meagre (Argyrosomus regius, Asso 1801) in the Eastern Adriatic Sea. Aquac. Res. 2018, 49, 1001–1007. [Google Scholar] [CrossRef]
- Mladineo, I. Prevalence of Ceratothoa oestroides (Risso, 1826), a cymothoid isopod parasite, in cultured sea bass (Dicentrarchus labrax, L.) on two farms in the middle of Adriatic Sea. Acta Adriat. 2002, 43, 97–102. [Google Scholar]
- Diamant, A.; Colorni, A.; Ucko, M. Parasite and disease transfer between cultured and wild coastal marine fish. CIESM Work. Monogr. 2007, 32, 49–53. [Google Scholar]
- Boxaspen, K. A review of the biology and genetics of sea lice. ICES J. Mar. Sci. 2006, 63, 1304–1316. [Google Scholar] [CrossRef] [Green Version]
- Mennerat, A.; Ugelvik, M.S.; Håkonsrud Jensen, C.; Skorping, A. Invest more and die faster: The life history of a parasite on intensive farms. Evol. Appl. 2017, 10, 890–896. [Google Scholar] [CrossRef]
- Todd, C.D.; Walker, A.M.; Ritchie, M.G.; Graves, J.A.; Walker, A.F. Population genetic differentiation of sea lice (Lepeophtheirus salmonis) parasitic on Atlantic and Pacific salmonids: Analyses of microsatellite DNA variation among wild and farmed hosts. Can. J. Fish. Aquat. Sci. 2004, 61, 1176–1190. [Google Scholar] [CrossRef]
- Besnier, F.; Kent, M.; Skern-Mauritzen, R.; Lien, S.; Malde, K.; Edvardsen, R.B.; Taylor, S.; Ljungfeldt, L.E.R.; Nilsen, F.; Glover, K.A. Human-induced evolution caught in action: SNP-array reveals rapid amphi-atlantic spread of pesticide resistance in the salmon ecotoparasite Lepeophtheirus salmonis. BMC Genom. 2014, 15, 937. [Google Scholar] [CrossRef] [Green Version]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fabian, I.; Rozsa, L. Biostatistics for parasitologists—A primer to Quantitative Parasitology. Trend. Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Manousaki, T.; Tsakogiannis, A.; Taggart, J.B.; Palaiokostas, C.; Tsaparis, D.; Lagnel, J.; Chatziplis, D.; Magoulas, A.; Papandroulakis, N.; Mylonas, C.C.; et al. Exploring a nonmodel teleost genome through rad sequencing-linkage mapping in common pandora, Pagellus erythrinus and comparative genomic analysis. G3 Genes Genomes Genet. 2016, 6, 509–519. [Google Scholar]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Fish Species | Season | N. of Fish | Mean Length ± SD (mm) | Prevalence of Infection (%) (Range) | Mean Intensity (Range) | Mean Abundance (Confidence Intervals) | v/m |
|---|---|---|---|---|---|---|---|
| Farmed fish Dicentrarchus labrax | total sample (N = 55,250) | 310.81 ± 8.53 * | 1.02 | 2.00 | 0.02 | ||
| I (2016) | summer (N = 14,000) | 320.11 ± 6.63 * | 0.7 (0.54–0.81) | 2.27 (2.14–2.43) | 0.01 | 2.50 | |
| II (2016) | autumn (N = 16,250) | 309.05 ± 4.15 * | 1.1 (0.96–1.29) | 2.00 | 0.02 | 1.98 | |
| III (2017) | winter (N = 11,250) | 296.74 ± 5.29 * | 1.2 (1.03–1.44) | 2.00 | 0.02 | 1.98 | |
| IV (2017) | spring (N = 13,750) | 300.23 ± 4.86 * | 1.1 (0.98–1.34) | 2.00 | 0.02 | 1.98 | |
| Total wild fish | total sample (N = 444) | 22.91 ± 7.62 | 8.1 (5.82–11.00) | 1.81 (1.34–2.53) | 0.15 (0.10–0.21) | 2.68 | |
| I (2016) | winter (N = 51) | 23.64 ± 6.75 | 9.8 (3.95–21.34) | 1.60 | 1.60 (0.04–0.31) | 1.62 | |
| II (2016) | spring (N = 187) | 19.37 ± 8.93 | 6.4 (3.65–10.89) | 1.58 (1.17–2.00) | 0.10 (0.05–0.17) | 1.86 | |
| III (2017) | summer (N = 48) | 27.44 ± 2.91 | 10.4 (4.20–22.67) | 1.40 | 0.15 (0.04–0.29) | 1.46 | |
| IV (2017) | autumn (N = 158) | 25.53 ± 4.90 | 8.9 (5.28–14.47) | 2.21 (1.57–3.79) | 0.20 (0.10–0.40) | 3.73 | |
| Boops boops | total sample (N = 268) | 27.72 ± 4.11 | 9.7 (6.67–13.94) | 2.04 | 0.20 (0.13–0.32) | 2.96 | |
| winter (N = 22) | 27.30 ± 3.04 | 9.1 (1.64–29.07) | 1.50 | 0.14 (0.00–0.36) | 1.60 | ||
| spring (N = 62) | 30.68 ± 2.83 | 9.7 (4.30–19.95) | 2.17 | 0.21 (0.08–0.42) | 2.21 | ||
| summer (N = 47) | 27.44 ± 2.905 | 10.6 (4.29–23.15) | 1.40 | 0.15 (0.04–0.30) | 1.45 | ||
| autumn (N = 137) | 26.53 ± 4.46 | 9.5 (5.32–15.60) | 2.31 (1.62–4.08) | 0.22 (0.11–0.43) | 3.81 | ||
| Sparus aurata | total sample (N = 20) | 11.87 ± 2.38 | 10.0 (1.81–31.99) | 1.50 | 0.15 (0.00–0.40) | 1.60 | |
| Dicentrarchus labrax | total sample (N = 56) | 11.48 ± 4.91 | 5.4 (1.48–14.90) | 1.00 | 0.05 (0.00–0.11) | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mladineo, I.; Hrabar, J.; Trumbić, Ž.; Manousaki, T.; Tsakogiannis, A.; Taggart, J.B.; Tsigenopoulos, C.S. Community Parameters and Genome-Wide RAD-Seq Loci of Ceratothoa oestroides Imply Its Transfer between Farmed European Sea Bass and Wild Farm-Aggregating Fish. Pathogens 2021, 10, 100. https://doi.org/10.3390/pathogens10020100
Mladineo I, Hrabar J, Trumbić Ž, Manousaki T, Tsakogiannis A, Taggart JB, Tsigenopoulos CS. Community Parameters and Genome-Wide RAD-Seq Loci of Ceratothoa oestroides Imply Its Transfer between Farmed European Sea Bass and Wild Farm-Aggregating Fish. Pathogens. 2021; 10(2):100. https://doi.org/10.3390/pathogens10020100
Chicago/Turabian StyleMladineo, Ivona, Jerko Hrabar, Željka Trumbić, Tereza Manousaki, Alexandros Tsakogiannis, John B. Taggart, and Costas S. Tsigenopoulos. 2021. "Community Parameters and Genome-Wide RAD-Seq Loci of Ceratothoa oestroides Imply Its Transfer between Farmed European Sea Bass and Wild Farm-Aggregating Fish" Pathogens 10, no. 2: 100. https://doi.org/10.3390/pathogens10020100
APA StyleMladineo, I., Hrabar, J., Trumbić, Ž., Manousaki, T., Tsakogiannis, A., Taggart, J. B., & Tsigenopoulos, C. S. (2021). Community Parameters and Genome-Wide RAD-Seq Loci of Ceratothoa oestroides Imply Its Transfer between Farmed European Sea Bass and Wild Farm-Aggregating Fish. Pathogens, 10(2), 100. https://doi.org/10.3390/pathogens10020100