
Occurrence and Characteristics of Staphylococcus aureus in a Hungarian Dairy Farm during a Control Program

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Place and Date of Samplings
2.2. Enumeration and Isolation of Staphylococcus aureus
2.3. Identification of Staphylococcus aureus with MALDI-TOF MS
2.4. Spa Typing
2.5. Haemolysis Testing
2.6. Antibiotic Susceptibility Testing
2.7. PCR Amplification of Staphylococcal Enterotoxin-Encoding Genes and Thermonuclease Gene
2.8. Statistical Analysis
3. Results
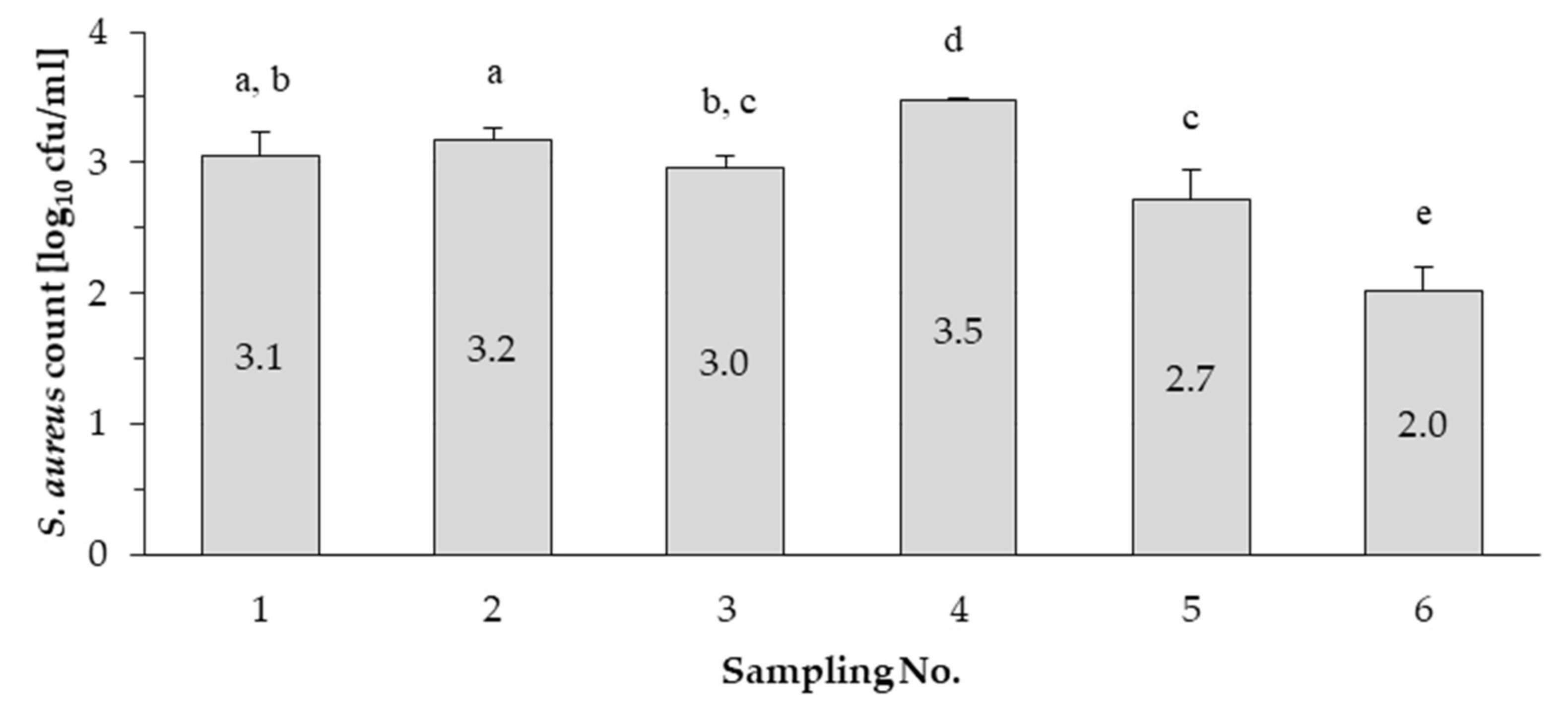
3.1. Enumeration and Isolation of Staphylococcus aureus
3.2. Identification of Staphylococcus aureus with MALDI-TOF MS
3.3. Spa Typing

{kind=link}
| Strain ID | Sampling No. | Best Score | Organism (Best Match) * | spa Type | spa Repeats |
|---|---|---|---|---|---|
| SA33 | 1. | 2.361 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA34 | 1. | 2.374 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA35A | 1. | 2.369 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA35B | 1. | 2.374 | 2 | t164 | r07r06r17r21r34r34r22r34 |
| SA39A | 2. | 2.342 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA39B | 2. | 2.312 | 3 | t164 | r07r06r17r21r34r34r22r34 |
| SA44 | 3. | 2.419 | 2 | t164 | r07r06r17r21r34r34r22r34 |
| SA45 | 3. | 2.359 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA53A | 4. | 2.318 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA53B | 4. | 2.410 | 1 | t164 | r07r06r17r21r34r34r22r34 |
| SA53D | 4. | 2.305 | 2 | t164 | r07r06r17r21r34r34r22r34 |
| SA54A | 5. | 2.302 | 2 | t164 | r07r06r17r21r34r34r22r34 |
| SA54B | 5. | 2.371 | 2 | t164 | r07r06r17r21r34r34r22r34 |
| SA54C | 5. | 2.500 | 1 | t1987 | r07r06r17r21r34 |
| SA57A | 6. | 2.333 | - | t164 | r07r06r17r21r34r34r22r34 |
| SA57B | 6. | 2.270 | - | t164 | r07r06r17r21r34r34r22r34 |
| SA57C | 6. | 2.417 | - | t164 | r07r06r17r21r34r34r22r34 |
3.4. Haemolysis Testing
3.5. Antibiotic Susceptibility Testing
3.6. Staphylococcal Enterotoxin-Encoding Genes
| Strain ID | Sampling No. | Characteristics | ||||
|---|---|---|---|---|---|---|
| Tellurite Reduction | Lecithinase Activity | Hemolysis | Antibiotic Resistance * | Detected Enterotoxin Genes | ||
| SA33 | 1. | dark grey | + | β | R (P) | sei |
| SA34 | 1. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA35A | 1. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA35B | 1. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA39A | 2. | dark grey | + | β | R (P) | sei, selm, seln, selo |
| SA39B | 2. | dark grey | + | β | R (P) | sei, selm, seln, selo |
| SA44 | 3. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA45 | 3. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA53A | 4. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA53B | 4. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA53D | 4. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA54A | 5. | dark grey | + | α + β | R (P) | seg, sei, selm, seln, selo |
| SA54B | 5. | dark grey | + | α + β | R (P) | sei, selm, seln, selo |
| SA54C | 5. | dark grey | + | weak β | R (P) | sei |
| SA57A | 6. | dark grey | + | β | R (P) | seg, sei, selm, seln, selo |
| SA57B | 6. | black | + | β | R (P) | sei, selm, seln, selo |
| SA57C | 6. | black | + | β | R (P) | seg, sei, selm, seln, selo |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DSMZ (German Collection of Microorganisms and Cell Cultures) Genus Staphylococcus. Available online: https://www.bacterio.net/genus/staphylococcus (accessed on 12 May 2020).
- Akindolire, M.A.; Babalola, O.O.; Ateba, C.N. Detection of Antibiotic Resistant Staphylococcus aureus from Milk: A Public Health Implication. Int. J. Environ. Res. Public Health 2015, 12, 10254–10275. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.; Ostyn, A.; Guillier, F.; Herbin, S.; Prufer, A.; Dragacci, S. How should Staphylococcal food poisoning outbreaks be characterized? Toxins 2010, 2, 2106–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergonier, D.; De Crémoux, R.; Rupp, R.; Lagriffoul, G.; Berthelot, X. Mastitis of dairy small ruminants. Vet. Res. 2003, 34, 689–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennekinne, J.; De Buyser, M.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2011, 36, 815–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loir, Y.L.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar] [PubMed]
- Benkerroum, N. Staphylococcal enterotoxins and enterotoxin-like toxins with special reference to dairy products: An overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 1943–1970. [Google Scholar] [CrossRef]
- Lina, G.; Bohach, G.A.; Nair, S.P.; Hiramatsu, K.; Jouvin-Marche, E.; Mariuzza, R. Standard nomenclature for the superantigens expressed by Staphylococcus. J. Infect. Dis. 2004, 189, 2334–2336. [Google Scholar] [CrossRef] [Green Version]
- Schelin, J.; Wallin-Carlquist, N.; Cohn, M.T.; Lindqvist, R.; Barker, G.C. The formation of Staphylococcus aureus enterotoxin in food environments and advances in risk assessment. Virulence 2011, 2, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Viçosa, G.N.; Moraes, P.M.; Yamazi, A.K.; Nero, L.A. Enumeration of coagulase and thermonuclease-positive Staphylococcus spp. in raw milk and fresh soft cheese: An evaluation of Baird-Parker agar, Rabbit Plasma Fibrinogen agar and the PetrifilmTM Staph Express count system. Food Microbiol. 2010, 27, 447–452. [Google Scholar] [CrossRef]
- Denny, C.B.; Tan, P.L.; Bohrer, C.W. Heat inactivation of staphylococcus enterotoxin A. J. Food Sci. 1966, 31, 762–767. [Google Scholar] [CrossRef]
- Genigeorgis, C.A. Present state of knowledge on staphylococcal intoxication. Int. J. Food Microbiol. 1989, 9, 327–360. [Google Scholar] [CrossRef]
- Gunvig, A.; Andresen, M.S.; Jacobsen, T.; Borggaard, C. Staphtox predictor—A dynamic mathematical model to predict formation of Staphylococcus enterotoxin during heating and fermentation of meat products. Int. J. Food Microbiol. 2018, 285, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Hnasko, R.; Lin, A.V.; McGarvey, J.A. Rapid Detection of Staphylococcal Enterotoxin-B by Lateral Flow Assay. Monoclon. Antib. Immunodiagn. Immunother. 2019, 38, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Umeda, K.; Nakamura, H.; Yamamoto, K.; Nishina, N.; Yasufuku, K.; Hirai, Y.; Hirayama, T.; Goto, K.; Hase, A.; Ogasawara, J. Molecular and epidemiological characterization of staphylococcal foodborne outbreak of Staphylococcus aureus harboring seg, sei, sem, sen, seo, and selu genes without production of classical enterotoxins. Int. J. Food. Microbiol. 2017, 256, 30–35. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and foodborne outbreaks in 2018. EFSA J. 2019, 17, 5926. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting using a novel software for spa-repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [Green Version]
- Mellmann, A.; Weniger, T.; Berssenbrügge, C.; Keckevoet, U.; Friedrich, A.W.; Harmsen, D.; Grundmann, H. Characterization of clonal relatedness among the natural population of Staphylococcus aureus strains by using spa sequence typing and the BURP (based upon repeat patterns) algorithm. J. Clin. Microbiol. 2008, 46, 2805–2808. [Google Scholar] [CrossRef] [Green Version]
- Vann, J.M.; Proctor, R.A. Cytotoxic effects of ingested Staphylococcus aureus on bovine endothelial cells: Role of S. aureus α-hemolysin. Microb. Pathog. 1988, 4, 443–453. [Google Scholar] [CrossRef]
- Medveczky, I.; Rusvai, M.; Varga, J.; Tuboly, S. Állatorvosi Járványtan I.—Állatorvosi Mikrobiológia, Bakteriológia, Virológia, Immunológia; Mezőgazda Kiadó: Budapest, Hungary, 1999; p. 612. [Google Scholar]
- Barcs, I. Az antibiotikum-érzékenység és-rezisztencia. In A Semmelweis Egyetem Egészségtudományi Kar Népegészségtani Intézetének kiadványa; Barcs, I., Ed.; Semmelweis Egyetem, Egészségtudományi Kar, Népegészségtani Intézet: Budapest, Hungary, 2009; p. 40. [Google Scholar]
- Angulo, F.J.; Nargund, V.N.; Chiller, T.C. Evidence of an association between use of anti-microbial agents in food animals and anti-microbial resistance among bacteria isolated from humans and the human health consequences of such resistance. J. Vet. Med. B 2004, 51, 374–379. [Google Scholar] [CrossRef]
- Carmeli, Y.; Troillet, N.; Karchmer, A.W.; Samore, M.H. Health and economic outcomes of antibiotic resistance in Pseudomonas aeruginosa. Arch. Intern. Med. 1999, 159, 1127–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, S.E. The relationship between antimicrobial resistance and patient outcomes: Mortality, length of hospital stay, and health care costs. Clin. Infect. Dis. 2006, 42, S82–S89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCallum, N.; Berger-Bachi, B.; Senn, M.M. Regulation of antibiotic resistance in Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Petróczki, F.M.; Tonamo, T.A.; Béri, B.; Peles, F. The effect of breed and stage of lactation on the microbiological status of raw milk. Acta Agrar. Debr. 2019, 1, 37–45. [Google Scholar] [CrossRef]
- MSZ EN ISO 6887-1. Élelmiszerek és Takarmányok Mikrobiológiája. A Vizsgálati Minták, az Alapszuszpenzió és a Decimális Hígítások Elkészítése Mikrobiológiai Vizsgálathoz. 1. Rész: Az Alapszuszpenzió és a Decimális Hígítások Elkészítésének Általános Szabályai; ISO: Geneva, Switzerland, 2017. [Google Scholar]
- MSZ EN ISO 6888-1. Élelmiszerek és Takarmányok Mikrobiológiája. Horizontális Módszer a Koagulázpozitív Sztafilokokkuszok (Staphylococcus Aureus és más Fajok) Számának Meghatározása. 1. Rész: Baird-Parker-agar Táptalajos Eljárás; ISO: Geneva, Switzerland, 2008. [Google Scholar]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [Green Version]
- SeqNet (European Network of Laboratories for Sequence Based Typing of Microbial Pathogens) DNA Sequencing of the Spa Gene. 2004. Available online: https://www.seqnet.org/downloads.html (accessed on 1 October 2020).
- Pereira, V.; Lopes, C.; Castro, A.; Silva, J.; Gibbs, P.; Teixeira, P. Characterization for enterotoxin production, virulence factors, and antibiotic susceptibility of Staphylococcus aureus isolates from various foods in Portugal. Food Microbiol. 2009, 26, 278–282. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. In M100, 27th ed.; Replaces M100-S26; CLSI: Wayne, NJ, USA, 2017; 282p. [Google Scholar]
- Bianchi, D.M.; Gallina, S.; Bellio, A.; Chiesa, F.; Civera, T.; Decastelli, L. Enterotoxin gene profiles of Staphylococcus aureus isolated from milk and dairy products in Italy. Lett. Appl. Microbiol. 2014, 58, 190–196. [Google Scholar] [CrossRef]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for Detection of Genes for Staphylococcus aureus Enterotoxins, Exfoliative Toxins, Toxic Shock Syndrome Toxin 1, and Methicillin Resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.K.; Rees, C.E.D.; Dodd, C.E.R. Development of a single-reaction multiplex PCR toxin typing assay for Staphylococcus aureus strains. Appl. Environ. Microbiol. 2000, 66, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Bania, J.; Dabrowska, A.; Bystron, J.; Korzekwa, K.; Chrzanowska, J.; Molenda, J. Distribution of newly described enterotoxin-like genes in Staphylococcus aureus from food. Int. J. Food Microbiol. 2006, 108, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-C.; Liao, W.-W.; Fan, C.-M.; Pai, W.-Y.; Chiou, C.-S.; Tsen, H.-Y. PCR detection of Staphylococcal enterotoxins (SEs) N, O, P, Q, R, U, and survey of SE types in Staphylococcus aureus isolates from food-poisoning cases in Taiwan. Int. J. Food Microbiol. 2008, 121, 66–73. [Google Scholar] [CrossRef] [PubMed]
- SPSS 220 for Windows; SPSS Inc.: Chicago, IL, USA, 2013.
- Peles, F.; Wagner, M.; Varga, L.; Hein, I.; Rieck, P.; Gutser, K.; Keresztúri, P.; Kardos, G.; Turcsányi, I.; Béri, B.; et al. Characterization of Staphylococcus aureus strains isolated from bovine milk in Hungary. Int. J. Food Microbiol. 2007, 118, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeinhom, M.M.A.; Aziz, R.A.L.; Mohammed, A.N.; Bernabucci, U. Impact of seasonal conditions on quality and pathogens content of milk in Friesian cows. Asian Australas. J. Anim. Sci. 2016, 29, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.Y.; Park, Y.K.; Koo, H.C.; Park, Y.H. spa typing and enterotoxin gene profile of Staphylococcus aureus isolated from bovine raw milk in Korea. J. Vet. Sci. 2010, 11, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Morandi, S.; Brasca, M.; Andrighetto, C.; Lombardi, A.; Lodi, R. Phenotypic and genotypic characterization of Staphylococcus aureus strains from Italian dairy products. Int. J. Microbiol. 2009, 2009, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Abo-Shama, U.H. Prevalence and antimicrobial susceptibility of Staphylococcus aureus isolated from cattle, buffalo, sheep and goats raws milk in Sohag governorate, Egypt. Assiut. Vet. Med. J. 2014, 60, 63–72. [Google Scholar]
- Visciano, P.; Pomilio, F.; Tofalo, R.; Sacchini, L.; Saletti, M.A.; Tieri, E.; Schirone, M.; Suzzi, G. Detection of methicillin-resistant Staphylococcus aureus in dairy cow farms. Food Control 2014, 46, 532–538. [Google Scholar] [CrossRef]
- André, M.C.D.P.B.; Campos, M.R.H.; Borges, L.J.; Kipnis, A.; Pimenta, F.C.; Serafini, Á.B. Comparison of Staphylococcus aureus isolates from food handlers, raw bovine milk and Minas Frescal cheese by antibiogram and pulsed-field gel electrophoresis following SmaI digestion. Food Control 2008, 19, 200–207. [Google Scholar] [CrossRef]
- Karahan, M.; Açik, M.N.; Cetinkaya, B. Investigation of toxin genes by polymerase chain reaction in Staphylococcus aureus strains isolated from bovine mastitis in Turkey. Foodborne Pathog. Dis. 2009, 6, 1029–1035. [Google Scholar] [CrossRef]
- Korpysa-Dzirba, W.; Osek, J. Identification of genes encoding classical staphylococcal enterotoxins in Staphylococcus aureus isolated from raw milk. Bull. Vet. Inst. Pulawy 2011, 55, 55–58. [Google Scholar]
- Dai, J.; Wu, S.; Huang, J.; Wu, Q.; Zhang, F.; Zhang, J.; Wang, J.; Ding, Y.; Zhang, S.; Yang, X.; et al. Prevalence and characterization of Staphylococcus aureus isolated from pasteurized milk in China. Front. Microbiol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cziszter, L.T.; Acatincăi, S.; Neciu, F.C.; Neamţ, R.I.; Ilie, D.E.; Costin, L.I.; Gavojdian, D.; Tripon, I. The influence of season on the cow milk quantity, quality and hygiene. Sci. Pap. Anim. Sci. Biotechnol. 2012, 45, 305–312. [Google Scholar]
- Petróczki, F.M.; Béri, B.; Peles, F. The effect of season on the microbiological status of raw milk. Acta Agrar. Debr. 2020, 1, 95–99. [Google Scholar] [CrossRef]
- Hill, B.; Smythe, B.; Lindsay, D.; Shepherd, J. Microbiology of raw milk in New Zealand. Int. J. Food Microbiol. 2012, 157, 305–308. [Google Scholar] [CrossRef] [PubMed]

| Enterotoxin genes | Primers | Nucleotide Sequences (5′-3′) | Amplification Size (bp) | Reference |
|---|---|---|---|---|
| sea | GSEAR-1 | GGTTATCAATGTGCGGGTGG | 102 | [36] |
| GSEAR-2 | CGGCACTTTTTTCTCTTCGG | |||
| seb | GSEBR-1 | GTATGGTGGTGTAACTGAGC | 164 | [36] |
| GSEBR-2 | CCAAATAGTGACGAGTTAGG | |||
| sec | GSECR-1 | AGATGAAGTAGTTGATGTGTATGG | 451 | [36] |
| GSECR-2 | CACACTTTTAGAATCAACCG | |||
| sed | GSEDR-1 | CCAATAATAGGAGAAAATAAAAG | 278 | [36] |
| GSEDR-2 | ATTGGTATTTTTTTTCGTTC | |||
| see | SA-U | TGTATGTATGGAGGTGTAAC | 213 | [37] |
| SA-E rev | GCCAAAGCTGTCTGAG | |||
| seg | SEG-F | GTTAGAGGAGGTTTTATG | 198 | [38] |
| SEG-R | TTCCTTCAACAGGTGGAGA | |||
| seh | SEH-F | CAACTGCTGATTTAGCTCAG | 173 | [38] |
| SEH-R | CCCAAACATTAGCACCA | |||
| sei | SEI-F | GGCCACTTTATCAGGACA | 328 | [38] |
| SEI-R | AACTTACAGGCAGTCCA | |||
| sej | SEJ-F | GTTCTGGTGGTAAACCA | 131 | [38] |
| SEJ-R | GCGGAACAACAGTTCTGA | |||
| selm | SEM-F | CATATCGCAACCGCTGA | 148 | [38] |
| SEM-R | TCAGCTGTTACTGTCGA | |||
| seln | SEN-F | GGCAATTAGACGAGTCA | 237 | [38] |
| SEN-R | ATCGTAACTCCTCCGTA | |||
| selo | SEO-F | GTCAAGTGTAGACCCTA | 288 | [38] |
| SEO-R | TGTACAGGCAGTATCCA | |||
| ser | SER1-F | AGATGTGTTTGGAATACCCTAT | 123 | [39] |
| SER2-R | CTATCAGCTGTGGAGTGCAT | |||
| nuc | NUC-F | GCGATTGATGGTGATACGGTT | 270 | [31] |
| NUC-R | AGCCAAGCCTTGACGAACTAAAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petróczki, F.M.; Pásztor, Á.; Szűcs, K.D.; Pál, K.; Kardos, G.; Albert, E.; Horváth, B.; Ungvári, E.; Béri, B.; Peles, F. Occurrence and Characteristics of Staphylococcus aureus in a Hungarian Dairy Farm during a Control Program. Pathogens 2021, 10, 104. https://doi.org/10.3390/pathogens10020104
Petróczki FM, Pásztor Á, Szűcs KD, Pál K, Kardos G, Albert E, Horváth B, Ungvári E, Béri B, Peles F. Occurrence and Characteristics of Staphylococcus aureus in a Hungarian Dairy Farm during a Control Program. Pathogens. 2021; 10(2):104. https://doi.org/10.3390/pathogens10020104
Chicago/Turabian StylePetróczki, Flóra M., Ákos Pásztor, Kata D. Szűcs, Károly Pál, Gábor Kardos, Ervin Albert, Brigitta Horváth, Erika Ungvári, Béla Béri, and Ferenc Peles. 2021. "Occurrence and Characteristics of Staphylococcus aureus in a Hungarian Dairy Farm during a Control Program" Pathogens 10, no. 2: 104. https://doi.org/10.3390/pathogens10020104
APA StylePetróczki, F. M., Pásztor, Á., Szűcs, K. D., Pál, K., Kardos, G., Albert, E., Horváth, B., Ungvári, E., Béri, B., & Peles, F. (2021). Occurrence and Characteristics of Staphylococcus aureus in a Hungarian Dairy Farm during a Control Program. Pathogens, 10(2), 104. https://doi.org/10.3390/pathogens10020104