Identification of Echinococcus granulosus Genotypes G1 and G3 by SNPs Genotyping Assays

 , , ,
, , ,
Abstract
:1. Introduction
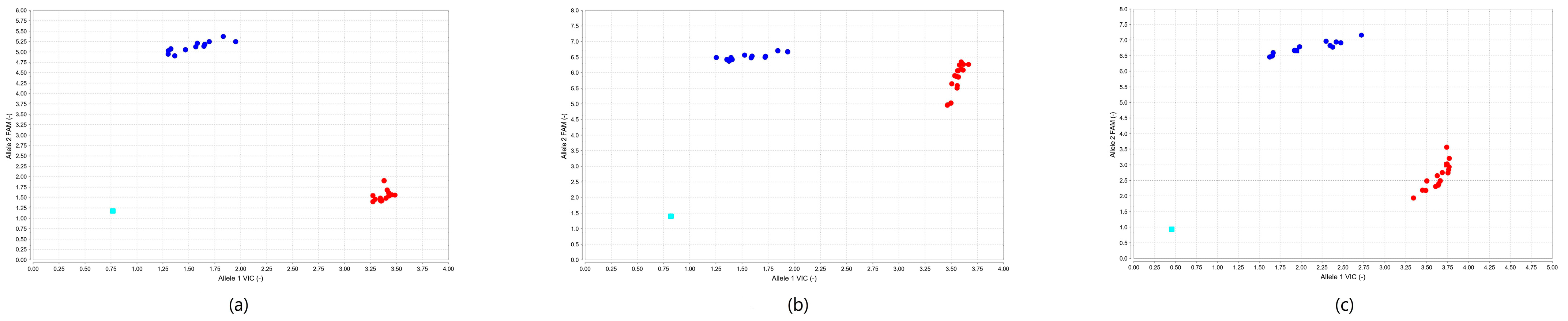
2. Results
2.1. Genotyping by DNA Sequencing
2.2. Genotyping by SYBR Green PCR
2.3. Genotyping by TaqMan PCR
3. Discussion
4. Materials and Methods
4.1. Samples Collection
4.2. Sequencing of the Mitochondrial nad5 Genes
4.3. SNP Genotyping
4.3.1. SYBR Green PCR with SNP Specific Primers
4.3.2. TaqMan PCR with SNP Specific Probes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| ID | Host | Isolate | Genotyping Method | Haplotype | Accession Number | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sanger sequencing | SYBR green PCR | TaqMan PCR | ||||||||
| nad5 (670 bp) | SNP 1123 | SNP1380 | SNP758 | SNP 1123 | SNP1380 | |||||
| 1 | ovine | 16762 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 2 | ovine | 16767 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR2 | MT993963 |
| 3 | caprine | 30487 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR3 | MT993964 |
| 4 | ovine | 40243 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR4 | MT993965 |
| 5 | bovine | 50508 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 6 | bovine | 51056 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR7 | MT993968 |
| 7 | bovine | 53965 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 8 | bovine | 60008 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR8 | MT993969 |
| 9 | ovine | 70802 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR5 | MT993966 |
| 10 | bovine | 74248 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR4 | MT993965 |
| 11 | bovine | 76558 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR6 | MT993967 |
| 12 | ovine | 76728 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 13 | ovine | 78417 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 14 | bovine | 78434 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 15 | bovine | 82760 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR1 | MT993962 |
| 16 | human | 82942 | G1 | G1 | G1 | G1 | G1 | G1 | nd5SAR9 | MT993970 |
| 17 | ovine | 35378 | G3 | G3 | G3 | G3 | G3 | G3 | ndSAR11 | MT993972 |
| 18 | bovine | 42450 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 19 | bovine | 50501 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR4 | MT993965 |
| 20 | bovine | 53437 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 21 | ovine | 57455 | G3 | G3 | G1/G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 22 | ovine | 68923 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 23 | ovine | 70804 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 24 | human | 75588 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 25 | ovine | 77580 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 26 | human | 84912 | G3 | G3 | G3 | G3 | G3 | G3 | nd5SAR9 | MT993970 |
| 27 | caprine | 95256 | G3 | G3 | G3 | G3 | G3 | G3 | ndSAR10 | MT993971 |
| 28 | human | 103212 | G3 | G3 | G3 | G3 | G3 | G3 | ndSAR12 | MT993973 |
References
- WHO|Working to Overcome the Global Impact of Neglected Tropical Diseases: First WHO Report on Neglected Tropical Diseases. Available online: https://www.who.int/neglected_diseases/resources/9789241564090/en/ (accessed on 9 December 2020).
- Romig, T.; Ebi, D.; Wassermann, M. Taxonomy and molecular epidemiology of Echinococcus granulosus sensu lato. Vet. Parasitol. 2015, 213, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Hüttner, M.; Nakao, M.; Wassermann, T.; Siefert, L.; Boomker, J.D.F.; Dinkel, A.; Sako, Y.; Mackenstedt, U.; Romig, T.; Ito, A. Genetic characterization and phylogenetic position of Echinococcus felidis Ortlepp, 1937 (Cestoda: Taeniidae) from the African lion. Int. J. Parasitol. 2008, 38, 861–868. [Google Scholar] [CrossRef]
- Deplazes, P.; Rinaldi, L.; Alvarez Rojas, C.A.; Torgerson, P.R.; Harandi, M.F.; Romig, T.; Antolova, D.; Schurer, J.M.; Lahmar, S.; Cringoli, G.; et al. Global distribution of alveolar and cystic echinococcosis. Adv. Parasitol. 2017, 95, 315–493. [Google Scholar] [PubMed] [Green Version]
- Alvarez Rojas, C.A.; Romig, T.; Lightowlers, M.W. Echinococcus granulosus sensu lato genotypes infecting humans—Review of current knowledge. Int. J. Parasitol. 2014, 44, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.A. The taxonomy, phylogeny and transmission of Echinococcus. Exp. Parasitol. 2008, 119, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Eckert, J.; Gemmell, M.A.; Meslin, F.-X.; Pawłowski, Z.S. World Health Organization World Organisation for Animal Health WHO/OIE Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern; World Organisation for Animal Health: Paris, France, 2001. [Google Scholar]
- Kinkar, L.; Laurimäe, T.; Sharbatkhori, M.; Mirhendi, H.; Kia, E.B.; Ponce-Gordo, F.; Andresiuk, V.; Simsek, S.; Lavikainen, A.; Irshadullah, M.; et al. New mitogenome and nuclear evidence on the phylogeny and taxonomy of the highly zoonotic tapeworm Echinococcus granulosus sensu stricto. Infect. Genet. Evol. 2017, 52, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Casulli, A.; Interisano, M.; Sreter, T.; Chitimia, L.; Kirkova, Z.; La Rosa, G.; Pozio, E. Genetic variability of Echinococcus granulosus sensu stricto in Europe inferred by mitochondrial DNA sequences. Infect. Genet. Evol. 2012, 12, 377–383. [Google Scholar] [CrossRef]
- Yanagida, T.; Mohammadzadeh, T.; Kamhawi, S.; Nakao, M.; Sadjjadi, S.M.; Hijjawi, N.; Abdel-Hafez, S.K.; Sako, Y.; Okamoto, M.; Ito, A. Genetic polymorphisms of Echinococcus granulosus sensu stricto in the Middle East. Parasitol. Int. 2012, 61, 599–603. [Google Scholar] [CrossRef]
- Laatamna, A.E.; Ebi, D.; Brahimi, K.; Bediaf, K.; Wassermann, M.; Souttou, K.; Romig, T. Frequency and genetic diversity of Echinococcus granulosus sensu stricto in sheep and cattle from the steppe region of Djelfa, Algeria. Parasitol. Res. 2019, 118, 89–96. [Google Scholar] [CrossRef]
- Ohiolei, J.A.; Yan, H.B.; Li, L.; Magaji, A.A.; Luka, J.; Zhu, G.Q.; Isaac, C.; Odoya, M.E.; Wu, Y.T.; Alvi, M.A.; et al. Cystic echinococcosis in Nigeria: First insight into the genotypes of Echinococcus granulosus in animals. Parasites Vectors 2019, 12, 392. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Jian, Y.; Zhang, X.; Ma, L.; Zhu, W.; Cai, Q.; Wu, S.; Wang, X.; Shi, B. Genetic characterization of Echinococcus isolates from various intermediate hosts in the Qinghai-Tibetan Plateau Area, China. Parasitology 2019, 146, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Kinkar, L.; Laurimäe, T.; Simsek, S.; Balkaya, I.; Casulli, A.; Manfredi, M.T.; Ponce-Gordo, F.; Varcasia, A.; Lavikainen, A.; González, L.M.; et al. High-resolution phylogeography of zoonotic tapeworm Echinococcus granulosus sensu stricto genotype G1 with an emphasis on its distribution in Turkey, Italy and Spain. Parasitology 2016, 143, 1790–1801. [Google Scholar] [CrossRef] [Green Version]
- Laurimäe, T.; Kinkar, L.; Andresiuk, V.; Haag, K.L.; Ponce-Gordo, F.; Acosta-Jamett, G.; Garate, T.; Gonzàlez, L.M.; Saarma, U. Genetic diversity and phylogeography of highly zoonotic Echinococcus granulosus genotype G1 in the Americas (Argentina, Brazil, Chile and Mexico) based on 8279 bp of mtDNA. Infect. Genet. Evol. 2016, 45, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Kinkar, L.; Laurimäe, T.; Acosta-Jamett, G.; Andresiuk, V.; Balkaya, I.; Casulli, A.; Gasser, R.B.; González, L.M.; Haag, K.L.; Zait, H.; et al. Distinguishing Echinococcus granulosus sensu stricto genotypes G1 and G3 with confidence: A practical guide. Infect. Genet. Evol. 2018, 64, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Brookes, A.J. The essence of SNPs. Gene 1999, 234, 177–186. [Google Scholar] [CrossRef]
- Kim, S.; Misra, A. SNP Genotyping: Technologies and Biomedical Applications. Annu. Rev. Biomed. Eng. 2007, 9, 289–320. [Google Scholar] [CrossRef]
- Syvänen, A.C. Accessing genetic variation: Genotyping single nucleotide polymorphisms. Nat. Rev. Genet. 2001, 2, 930–942. [Google Scholar] [CrossRef]
- Primmer, C.R.; Borge, T.; Lindell, J.; Saetre, G.-P. Single-nucleotide polymorphism characterization in species with limited available sequence information: High nucleotide diversity revealed in the avian genome. Mol. Ecol. 2002, 11, 603–612. [Google Scholar] [CrossRef]
- Vignal, A.; Milan, D.; Sancristobal, M.; Eggen, A. A review on SNP and other types of molecular markers and their use in animal genetics. Genet. Sel. Evol 2002, 34, 275–305. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.M.; Ma, X.; Boyko, A.R.; Bustamante, C.D.; Oleksiak, M.F. SNP identification, verification, and utility for population genetics in a non-model genus. BMC Genet. 2010, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhas, D.B.B.; Hiasindh Ashmi, A.; Vishnu Bhat, B.; Parija, S.C.; Banupriya, N. Modified low cost SNP genotyping technique using cycle threshold (Ct) & melting temperature (Tm) values in allele specific real-time PCR. Indian J. Med. Res. 2015, 142, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Tian, Y.; Ran, T.; Gao, Z. Genotyping and quantification techniques for single-nucleotide polymorphisms. TrAC—Trends Anal. Chem. 2015, 69, 1–13. [Google Scholar] [CrossRef]
- Kwok, P.Y. Methods for genotyping single nucleotide polymorphisms. Annu. Rev. Genomics Hum. Genet. 2001, 2, 235–258. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Wang, M.; Yan, X.; Abdullahi, A.Y.; Hang, J.; Zhang, P.; Huang, Y.; Liu, Y.; Sun, Y.; Ran, R.; et al. T m-shift detection of dog-derived ancylostoma ceylanicum and a. caninum. Biomed Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Fu, Y.; Abdullahi, A.Y.; Wang, M.; Shi, X.; Yang, F.; Yu, X.; Yan, X.; Zhang, P.; Hang, J.; et al. Development of Tm-shift genotyping method for detection of cat-derived Giardia lamblia. Parasitol. Res. 2017, 116, 1151–1157. [Google Scholar] [CrossRef]
- Wang, J.; Chuang, K.; Ahluwalia, M.; Patel, S.; Umblas, M.; Mirel, D.; Higuchi, R.; Germer, S. High-throughput SNP genotyping by single-tube PCR with Tm-shift primers. Biotechniques 2005, 39, 885–892. [Google Scholar] [CrossRef]
- Shen, G.Q.; Abdullah, K.G.; Wang, Q.K. The TaqMan method for SNP genotyping. Methods Mol. Biol. 2009, 578, 293–306. [Google Scholar] [CrossRef]
- Hui, L.; DelMonte, T.; Ranade, K. Genotyping Using the TaqMan Assay. Curr. Protoc. Hum. Genet. 2008, 56, 2.10.1–2.10.8. [Google Scholar] [CrossRef]
- Broccanello, C.; Chiodi, C.; Funk, A.; McGrath, J.M.; Panella, L.; Stevanato, P. Comparison of three PCR-based assays for SNP genotyping in plants. Plant Methods 2018, 14, 28. [Google Scholar] [CrossRef] [Green Version]
- Laurimäe, T.; Kinkar, L.; Romig, T.; Umhang, G.; Casulli, A.; Omer, R.A.; Sharbatkhori, M.; Mirhendi, H.; Ponce-Gordo, F.; Lazzarini, L.E.; et al. Analysis of nad2 and nad5 enables reliable identification of genotypes G6 and G7 within the species complex Echinococcus granulosus sensu lato. Infect. Genet. Evol. 2019, 74, 103941. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Quality value in TaqMan® Genotyper Software. Available online: https://www.thermofisher.com/content/dam/LifeTech/migration/files/pcr/pdfs.par.74504.file.tmp/co04197-genotyper-quality-values.pdf (accessed on 9 November 2020).


| SNP | Primer | Sequence | Product Length (bp) | Product GC Content (%) |
|---|---|---|---|---|
| 1123 | SNP_1123_G1 Fw SNP_1123_G3 Fw SNP_1123 Rv | 5’-GCGGGCAGGGCGGCCGTTATGACTATTTGTTACATTTgG-3’ 5’-GCGGGCCGTTATGACTATTTGTTACATTTgA-3’ 5’-ACAAAGCCACAATCTTCTTC-3’ | 141 133 | 41.10 37.68 |
| 1380 | SNP_1380_G1 Fw SNP_1380_G3 Fw SNP_1380 Rv | 5’- GCGGGCAGGGCGGCGGGTGGTTCACAGGCTAaG -3’ 5’- GCGGGCGGGTGGTTCACAGGCTAaA -3’ 5’- AACAACCCAAACAAATTCC -3’ | 86 78 | 53.85 49.40 |
| Genotype G1 | Genotype G3 | |||
|---|---|---|---|---|
| Ct (Mean ± SD) | Tm °C (Mean ± SD) | Ct (Mean ± SD) | Tm °C (Mean ± SD) | |
| SNP1123 | 21.52 ± 3.72 | 76.51 ± 0.22 | 26.42 ± 2.87 | 74.69 ± 0.30 |
| SNP1380 | 21.63 ± 3.12 | 79.09 ± 0.29 | 24.10 ± 2.95 | 76.87 ± 0.23 |
| SNP | Primer | Sequence |
|---|---|---|
| 758 | SNP_758_F SNP_758_R Probe_G1 Probe_G3 | 5’-GGTTTATGTTGTTGAAGTTGATTGTTTTGT-3’ 5’-AAAACCTAACAACACCTAAATACTCTCAAAGAA-3’ VIC-5’- TGTTGGTATGTAGTGGTGAT-3’ FAM-5’- TGTTGGTATGTACTGGTGAT-3’ |
| 1123 | SNP_1123_F SNP_1123_R Probe_G1 Probe_G3 | 5’-CTGGTGTTTGGTTTGTTATGCGTTA-3’ 5’-CCAGTAATAAAAACCGTCAACAAAAGCA-3’ VIC-5’-CGACCTACCAAAATG-3’ FAM-5’- CCGACCTACTAAAATG-3’ |
| 1380 | SNP_1380_F SNP_1380_R Probe_G1 Probe_G3 | 5’-GTGATGTGATGAGCGGTAGGG-3’ 5’-CACGACCCATACAAAACAGACCTAT-3’ VIC-5’- CAGGCTAGGAATTGT-3’ FAM-5’-CAGGCTAGAAATTGT-3’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonelli, P.; Dei Giudici, S.; Peruzzu, A.; Mura, L.; Santucciu, C.; Maestrale, C.; Masala, G. Identification of Echinococcus granulosus Genotypes G1 and G3 by SNPs Genotyping Assays. Pathogens 2021, 10, 125. https://doi.org/10.3390/pathogens10020125
Bonelli P, Dei Giudici S, Peruzzu A, Mura L, Santucciu C, Maestrale C, Masala G. Identification of Echinococcus granulosus Genotypes G1 and G3 by SNPs Genotyping Assays. Pathogens. 2021; 10(2):125. https://doi.org/10.3390/pathogens10020125
Chicago/Turabian StyleBonelli, Piero, Silvia Dei Giudici, Angela Peruzzu, Lorena Mura, Cinzia Santucciu, Caterina Maestrale, and Giovanna Masala. 2021. "Identification of Echinococcus granulosus Genotypes G1 and G3 by SNPs Genotyping Assays" Pathogens 10, no. 2: 125. https://doi.org/10.3390/pathogens10020125
APA StyleBonelli, P., Dei Giudici, S., Peruzzu, A., Mura, L., Santucciu, C., Maestrale, C., & Masala, G. (2021). Identification of Echinococcus granulosus Genotypes G1 and G3 by SNPs Genotyping Assays. Pathogens, 10(2), 125. https://doi.org/10.3390/pathogens10020125