
Development of a Multiplex PCR and Magnetic DNA Capture Assay for Detecting Six Species Pathogens of the Genera Anaplasma and Ehrlichia in Canine, Bovine, Caprine and Ovine Blood Samples from Grenada, West Indies

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Optimization of the xMAP Assay
2.2. Analytical Specificity
2.3. The Limit of Detection and Analytical Sensitivity
2.4. Repeatability
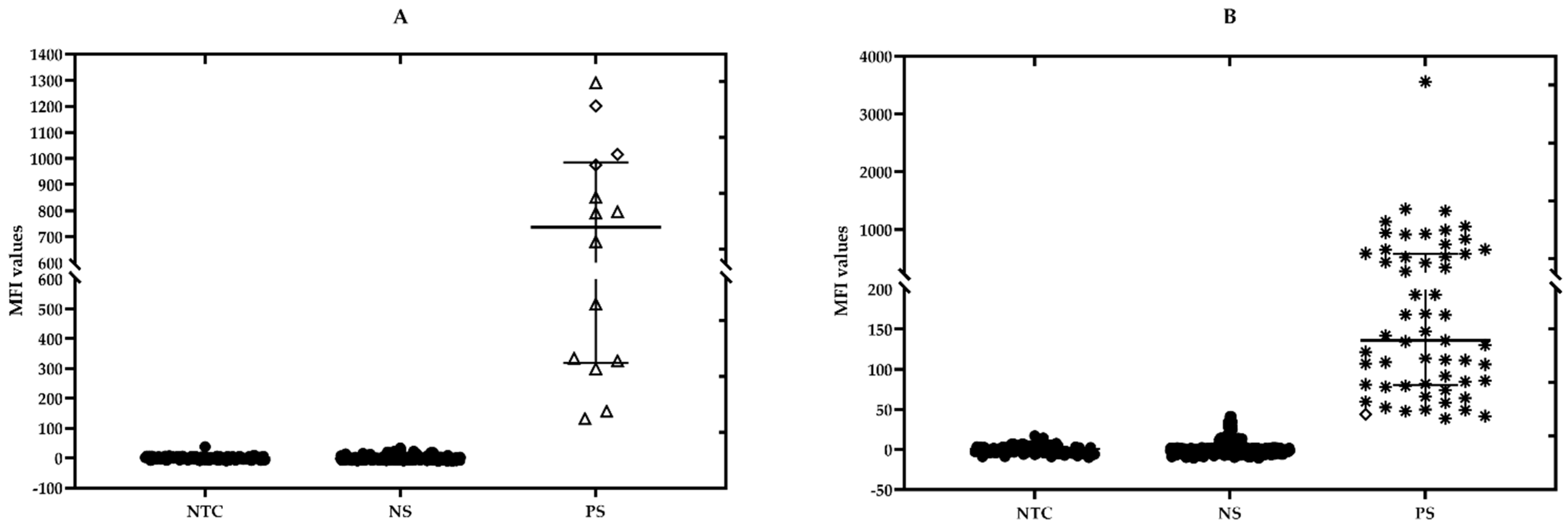
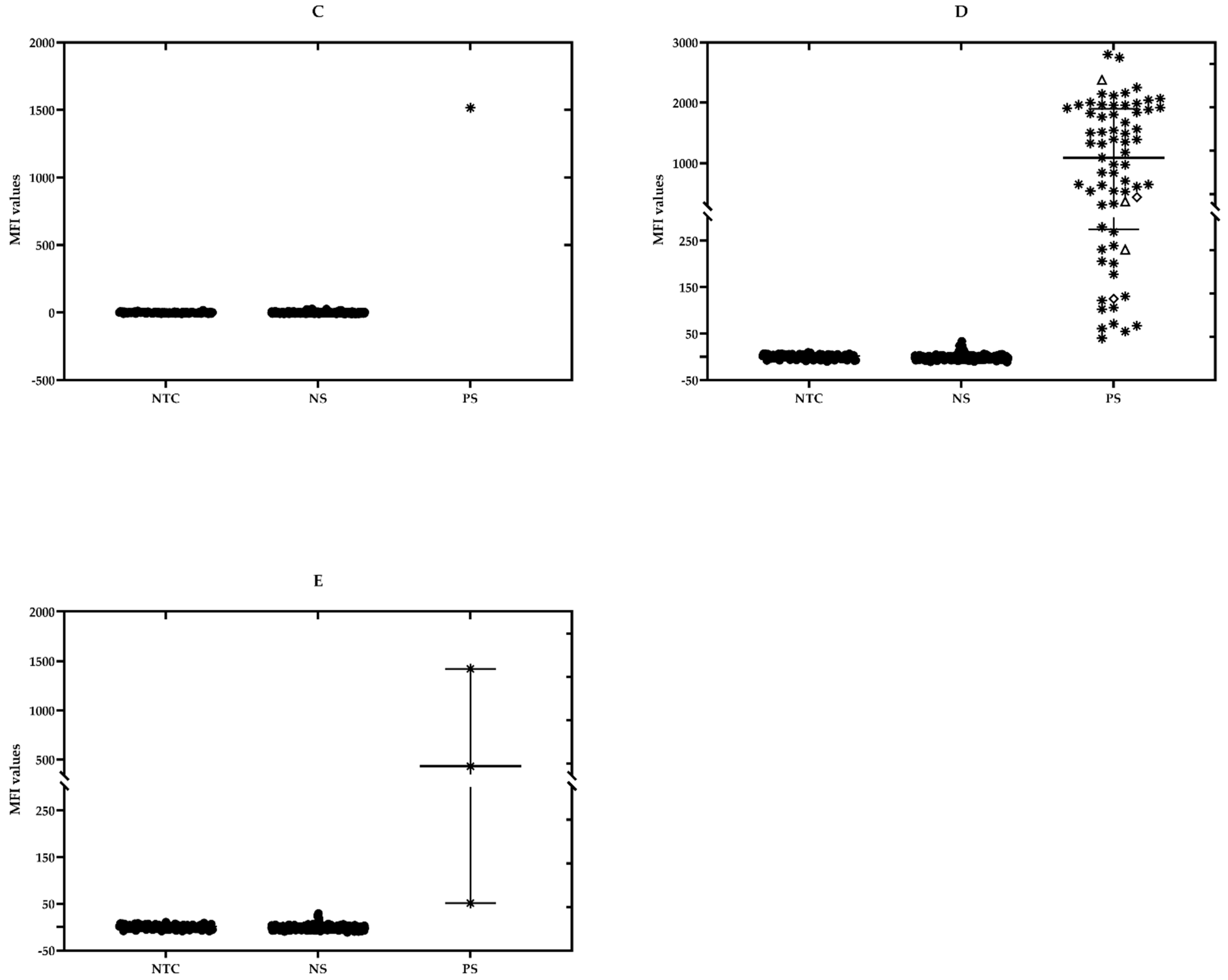
2.5. Testing of the Field Samples
2.6. Confirmatory PCR Assays
3. Discussion
4. Materials and Methods
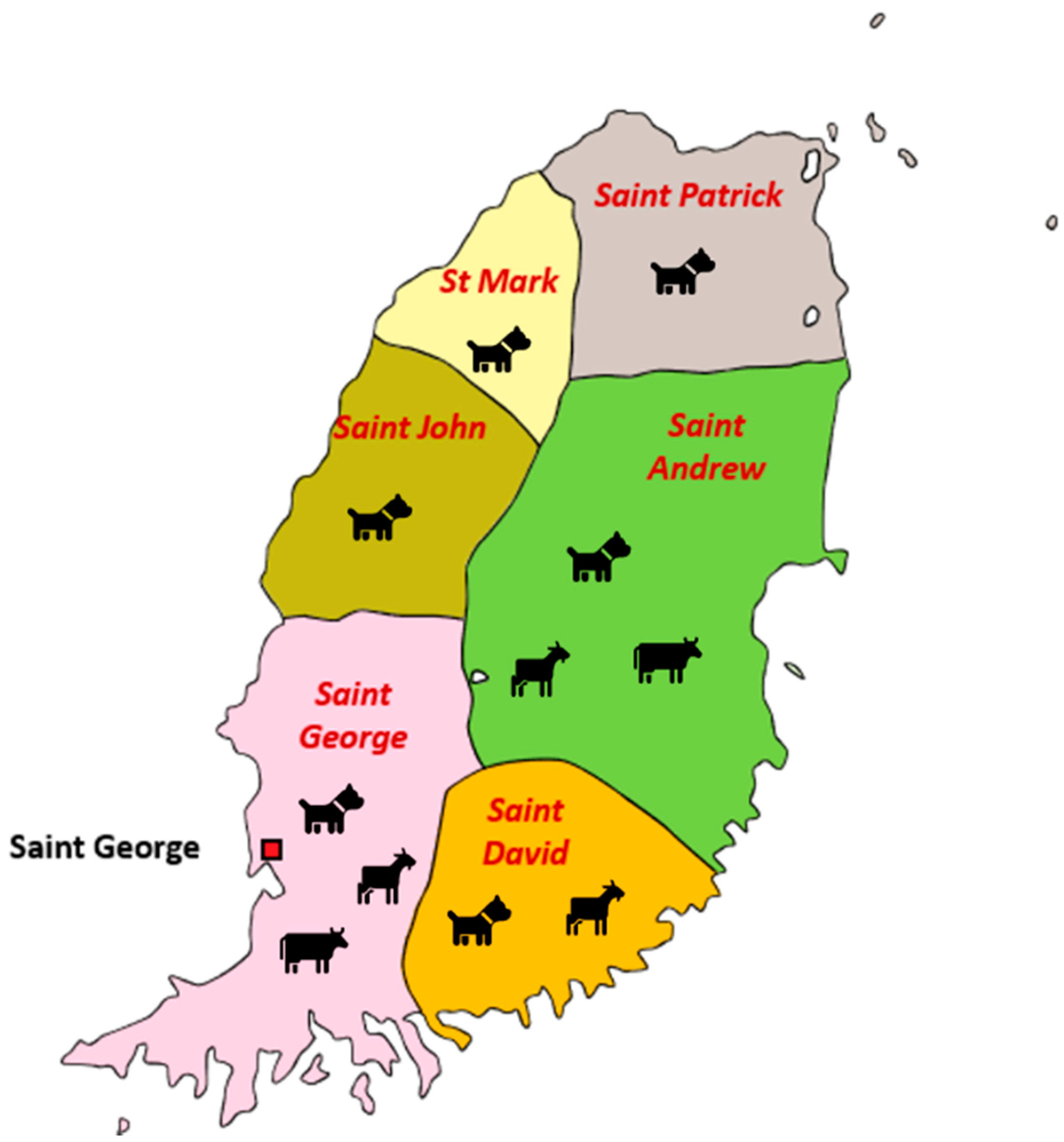
4.1. Collection of the Field Specimens
4.2. DNA Extraction
4.3. Optimization of the PCR and xMAP Hybridization Assay Conditions
4.4. DNA Amplification for xMAP Assay
4.5. Oligonucleotide xMAP Assay
4.5.1. Oligonucleotide Probe Design
4.5.2. Oligonucleotide Probe Coupling to xMAP Beads
4.5.3. Direct Hybridization of Blood-Derived DNA Samples to Six Oligonucleotide Probe-Coupled xMAP Beads
4.5.4. Determination of the Analytical Specificity of the Luminex Assay
4.5.5. Determination of Limit of Detection and Analytical Sensitivity
4.5.6. Intra-Assay and Inter-Assay Variability
4.6. Confirmation of the Results by PCR and Sequencing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rymaszewska, A.; Grenda, S. Bacteria of the genus Anaplasma–characteristics of Anaplasma and their vectors: A review. Vet. Med. 2008, 53, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.; Golovljova, I. Anaplasma, Ehrlichia, and “Candidatus Neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Tickborne Disease Surveillance Data Summary. Available online: https://www.cdc.gov/ticks/data-summary/index.html (accessed on 22 August 2020).
- Alhassan, A.; Pumidonming, W.; Okamura, M.; Hirata, H.; Battsetseg, B.; Fujisaki, K.; Yokoyama, N.; Igarashi, I. Development of a single-round and multiplex PCR method for the simultaneous detection of Babesia caballi and Babesia equi in horse blood. Vet. Parasitol. 2005, 129, 43–49. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Hegarty, B.C.; Hancock, S.I. Sequential evaluation of dogs naturally infected with Ehrlichia canis, Ehrlichia chaffeensis, Ehrlichia equi, Ehrlichia ewingii, or Bartonella vinsonii. J. Clin. Microbiol. 1998, 36, 2645–2651. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.F.; Novosel, V.; Chang, C.F.; Kim, J.B.; Shin, S.J.; Lein, D.H. Detection of human granulocytic ehrlichiosis agent and Borrelia burgdorferi in ticks by polymerase chain reaction. J. Vet. Diagn. Investig. 1998, 10, 56–59. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Zhang, Y.; Jian, F.; Zhang, L.; Wang, R.; Cao, S.; Wang, X.; Yan, Y.; Ning, C. Development of duplex PCR for simultaneous detection of Theileria spp. and Anaplasma spp. in sheep and goats. Exp. Parasitol. 2017, 176, 1–7. [Google Scholar] [CrossRef]
- Hoskins, J.D.; Breitschwerdt, E.B.; Gaunt, S.D.; French, T.W.; Burgdorfer, W. Antibodies to Ehrlichia canis, Ehrlichia platys, and spotted fever group rickettsiae in Louisiana dogs. J. Vet. Intern. Med. 1988, 2, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Hua, P.; Yuhai, M.; Shide, T.; Yang, S.; Bohai, W.; Xiangrui, C. Canine ehrlichiosis caused simultaneously by Ehrlichia canis and Ehrlichia platys. Microbiol. Immunol. 2000, 44, 737–739. [Google Scholar] [CrossRef] [PubMed]
- Kordick, S.K.; Breitschwerdt, E.B.; Hegarty, B.C.; Southwick, K.L.; Colitz, C.M.; Hancock, S.I.; Bradley, J.M.; Rumbough, R.; McPherson, J.T.; MacCormack, J.N. Coinfection with multiple tick-borne pathogens in a Walker Hound kennel in North Carolina. J. Clin. Microbiol. 1999, 37, 2631–2638. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, V.; Wijnveld, M.; Majekodunmi, A.O.; Dongkum, C.; Fajinmi, A.; Dogo, A.G.; Thrusfield, M.; Mugenyi, A.; Vaumourin, E.; Igweh, A.C.; et al. Tick-borne pathogens of zoonotic and veterinary importance in Nigerian cattle. Parasites Vectors 2016, 9, 217. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Mascarelli, P.E.; Havenga, L.N.; Naidoo, V.; Breitschwerdt, E.B. Co-infection with Anaplasma platys, Bartonella henselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasit Vectors 2013, 6, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinkoth, J.H.; Ewing, S.A.; Cowell, R.L.; Dawson, J.E.; Warner, C.K.; Mathew, J.S.; Bowles, M.; Thiessen, A.E.; Panciera, R.J.; Fox, C. Morphologic and molecular evidence of a dual species ehrlichial infection in a dog presenting with inflammatory central nervous system disease. J. Vet. Intern. Med. 1998, 12, 389–393. [Google Scholar] [CrossRef]
- Njiiri, N.E.; Bronsvoort, B.M.; Collins, N.E.; Steyn, H.C.; Troskie, M.; Vorster, I.; Thumbi, S.M.; Sibeko, K.P.; Jennings, A.; van Wyk, I.C.; et al. The epidemiology of tick-borne haemoparasites as determined by the reverse line blot hybridization assay in an intensively studied cohort of calves in western Kenya. Vet. Parasitol. 2015, 210, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Rajput, Z.I.; Hu, S.H.; Arijo, A.G.; Habib, M.; Khalid, M. Comparative study of Anaplasma parasites in tick carrying buffaloes and cattle. J. Zhejiang Univ. Sci. B 2005, 6, 1057–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringo, A.E.; Adjou Moumouni, P.F.; Taioe, M.; Jirapattharasate, C.; Liu, M.; Wang, G.; Gao, Y.; Guo, H.; Lee, S.H.; Zheng, W.; et al. Molecular analysis of tick-borne protozoan and rickettsial pathogens in small ruminants from two South African provinces. Parasitol. Int. 2018, 67, 144–149. [Google Scholar] [CrossRef]
- De Tommasi, A.S.; Otranto, D.; Dantas-Torres, F.; Capelli, G.; Breitschwerdt, E.B.; de Caprariis, D. Are vector-borne pathogen co-infections complicating the clinical presentation in dogs? Parasites Vectors 2013, 6, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonakis, M.E.; Koutinas, A.F.; Baneth, G.; Polizopoulou, Z.; Fytianou, A. Mixed Ehrlichia canis, Hepatozoon canis, and presumptive Anaplasma phagocytophilum infection in a dog. Vet. Clin. Pathol. 2004, 33, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, A.D.; Birkenheuer, A.J.; Juopperi, T.; Levy, M.G.; Breitschwerdt, E.B. Concurrent bartonellosis and babesiosis in a dog with persistent thrombocytopenia. J. Am. Vet. Med. Assoc. 2003, 223, 1306–1310. [Google Scholar] [CrossRef]
- Gaunt, S.; Beall, M.; Stillman, B.; Lorentzen, L.; Diniz, P.; Chandrashekar, R.; Breitschwerdt, E. Experimental infection and co-infection of dogs with Anaplasma platys and Ehrlichia canis: Hematologic, serologic and molecular findings. Parasit Vectors 2010, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Gal, A.; Harrus, S.; Arcoh, I.; Lavy, E.; Aizenberg, I.; Mekuzas-Yisaschar, Y.; Baneth, G. Coinfection with multiple tick-borne and intestinal parasites in a 6-week-old dog. Can. Vet. J. 2007, 48, 619–622. [Google Scholar] [PubMed]
- Otranto, D.; Dantas-Torres, F.; Breitschwerdt, E.B. Managing canine vector-borne diseases of zoonotic concern: Part two. Trends Parasitol. 2009, 25, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.S.; Bakken, J.S.; Folk, S.M.; Paddock, C.D.; Bloch, K.C.; Krusell, A.; Sexton, D.J.; Buckingham, S.C.; Marshall, G.S.; Storch, G.A.; et al. Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever, ehrlichioses, and anaplasmosis--United States: A practical guide for physicians and other health-care and public health professionals. MMWR Recomm. Rep. 2006, 55, 1–27. [Google Scholar] [PubMed]
- Reller, M.E.; Dumler, J.S. Ehrlichia, Anaplasma, and related intracellular bacteria. Man. Clin. Microbiol. 2015, 1135–1149. [Google Scholar] [CrossRef]
- Walker, D.H.; Bouyer, D.H. Rickettsia and orientia. In Manual of Clinical Microbiology, 11th ed.; Jorgensen, J.H., Carroll, K.C., Funke, G., Pfaller, M.A., Landry, M.L., Richter, S.S., Warnock, D.W., Carroll, K.C., Funke, G., Bernard, K.A., et al., Eds.; American Society of Microbiology Press: Washington, DC, USA, 2015; pp. 1122–1134. [Google Scholar]
- Chandrashekar, R.; Mainville, C.A.; Beall, M.J.; O’Connor, T.; Eberts, M.D.; Alleman, A.R.; Gaunt, S.D.; Breitschwerdt, E.B. Performance of a commercially available in-clinic ELISA for the detection of antibodies against Anaplasma phagocytophilum, Ehrlichia canis, and Borrelia burgdorferi and Dirofilaria immitis antigen in dogs. Am. J. Vet. Res. 2010, 71, 1443–1450. [Google Scholar] [CrossRef]
- Malheiros, J.; Costa, M.M.; do Amaral, R.B.; de Sousa, K.C.M.; André, M.R.; Machado, R.Z.; Vieira, M.I.B. Identification of vector-borne pathogens in dogs and cats from Southern Brazil. Ticks Tick Borne Dis. 2016, 7, 893–900. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, T.P.; Hanscom, J.L.; Hegarty, B.C.; Groat, R.G.; Breitschwerdt, E.B. Comparison of an indirect immunofluorescence assay, western blot analysis, and a commercially available ELISA for detection of Ehrlichia canis antibodies in canine sera. Am. J. Vet. Res. 2006, 67, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Biggs, H.M.; Behravesh, C.B.; Bradley, K.K.; Dahlgren, F.S.; Drexler, N.A.; Dumler, J.S.; Folk, S.M.; Kato, C.Y.; Lash, R.R.; Levin, M.L.; et al. Diagnosis and Management of Tickborne Rickettsial Diseases: Rocky Mountain Spotted Fever and Other Spotted Fever Group Rickettsioses, Ehrlichioses, and Anaplasmosis—United States. MMWR Recomm. Rep. 2016, 65, 1–44. [Google Scholar] [CrossRef]
- World Organization for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, Chapter 3.4.1. Bovine Anaplasmosis. Available online: https://www.oie.int/en/standard-setting/terrestrial-manual/access-online/ (accessed on 9 February 2021).
- Chen, S.M.; Dumler, J.S.; Bakken, J.S.; Walker, D.H. Identification of a granulocytotropic Ehrlichia species as the etiologic agent of human disease. J. Clin. Microbiol. 1994, 32, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.E.; Biggie, K.L.; Warner, C.K.; Cookson, K.; Jenkins, S.; Levine, J.F.; Olson, J.G. Polymerase chain reaction evidence of Ehrlichia chaffeensis, an etiologic agent of human ehrlichiosis, in dogs from southeast Virginia. Am. J. Vet. Res. 1996, 57, 1175–1179. [Google Scholar]
- Eddlestone, S.M.; Gaunt, S.D.; Neer, T.M.; Boudreaux, C.M.; Gill, A.; Haschke, E.; Corstvet, R.E. PCR detection of Anaplasma platys in blood and tissue of dogs during acute phase of experimental infection. Exp. Parasitol. 2007, 115, 205–210. [Google Scholar] [CrossRef]
- Hulínská, D.; Langrová, K.; Pejcoch, M.; Pavlásek, I. Detection of Anaplasma phagocytophilum in animals by real-time polymerase chain reaction. Apmis 2004, 112, 239–247. [Google Scholar] [CrossRef]
- Sirigireddy, K.R.; Ganta, R.R. Multiplex detection of Ehrlichia and Anaplasma species pathogens in peripheral blood by real-time reverse transcriptase-polymerase chain reaction. J. Mol. Diagn. 2005, 7, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Doyle, C.K.; Labruna, M.B.; Breitschwerdt, E.B.; Tang, Y.W.; Corstvet, R.E.; Hegarty, B.C.; Bloch, K.C.; Li, P.; Walker, D.H.; McBride, J.W. Detection of medically important Ehrlichia by quantitative multicolor TaqMan real-time polymerase chain reaction of the dsb gene. J. Mol. Diagn. 2005, 7, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Benevenute, J.L.; Dumler, J.S.; Ogrzewalska, M.; Roque, A.L.R.; Mello, V.V.C.; de Sousa, K.C.M.; Gonçalves, L.R.; D’Andrea, P.S.; de Sampaio Lemos, E.R.; Machado, R.Z.; et al. Assessment of a quantitative 5’ nuclease real-time polymerase chain reaction using groEL gene for Ehrlichia and Anaplasma species in rodents in Brazil. Ticks Tick Borne Dis. 2017, 8, 646–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lew, A.E.; Gale, K.R.; Minchin, C.M.; Shkap, V.; de Waal, D.T. Phylogenetic analysis of the erythrocytic Anaplasma species based on 16S rDNA and GroEL (HSP60) sequences of A. marginale, A. centrale, and A. ovis and the specific detection of A. centrale vaccine strain. Vet. Microbiol. 2003, 92, 145–160. [Google Scholar] [CrossRef]
- Lew, A.E.; Bock, R.E.; Minchin, C.M.; Masaka, S. A msp1alpha polymerase chain reaction assay for specific detection and differentiation of Anaplasma marginale isolates. Vet. Microbiol. 2002, 86, 325–335. [Google Scholar] [CrossRef]
- Vidotto, M.C.; Kano, S.F.; Gregori, F.; Headley, S.A.; Vidotto, O. Phylogenetic analysis of Anaplasma marginale strains from Paraná State, Brazil, using the msp1alpha and msp4 genes. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 404–411. [Google Scholar] [CrossRef]
- Gusa, A.A.; Buller, R.S.; Storch, G.A.; Huycke, M.M.; Machado, L.J.; Slater, L.N.; Stockham, S.L.; Massung, R.F. Identification of a p28 gene in Ehrlichia ewingii: Evaluation of gene for use as a target for a species-specific PCR diagnostic assay. J. Clin. Microbiol. 2001, 39, 3871–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Xiong, Q.; Kikuchi, T.; Rikihisa, Y. Identification of 19 polymorphic major outer membrane protein genes and their immunogenic peptides in Ehrlichia ewingii for use in a serodiagnostic assay. Clin. Vaccine Immunol. 2008, 15, 402–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inokuma, H.; Brouqui, P.; Drancourt, M.; Raoult, D. Citrate synthase gene sequence: A new tool for phylogenetic analysis and identification of Ehrlichia. J. Clin. Microbiol. 2001, 39, 3031–3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Gibson, K.; Fitzpatrick, D.; Stone, D.; Noel, T.; MacPherson, C. Vector-borne diseases in the Caribbean: History and current status. Cab. Rev. 2016, 11, 1–28. [Google Scholar] [CrossRef]
- Lanza-Perea, M.; Zieger, U.; Qurollo, B.A.; Hegarty, B.C.; Pultorak, E.L.; Kumthekar, S.; Bruhl-Day, R.; Breitschwerdt, E.B. Intraoperative bleeding in dogs from Grenada seroreactive to Anaplasma platys and Ehrlichia canis. J. Vet. Intern. Med. 2014, 28, 1702–1707. [Google Scholar] [CrossRef]
- Wilkerson, M.J.; Black, K.E.; Lanza-Perea, M.; Sharma, B.; Gibson, K.; Stone, D.M.; George, A.; Nair, A.D.; Ganta, R.R. Initial development and preliminary evaluation of a multiplex bead assay to detect antibodies to Ehrlichia canis, Anaplasma platys, and Ehrlichia chaffeensis outer membrane peptides in naturally infected dogs from Grenada, West Indies. J. Vet. Diagn. Investig. 2017, 29, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Yabsley, M.J.; McKibben, J.; Macpherson, C.N.; Cattan, P.F.; Cherry, N.A.; Hegarty, B.C.; Breitschwerdt, E.B.; O’Connor, T.; Chandrashekar, R.; Paterson, T.; et al. Prevalence of Ehrlichia canis, Anaplasma platys, Babesia canis vogeli, Hepatozoon canis, Bartonella vinsonii berkhoffii, and Rickettsia spp. in dogs from Grenada. Vet. Parasitol. 2008, 151, 279–285. [Google Scholar] [CrossRef]
- Camus, E.; Barré, N. Vector situation of tick-borne diseases in the Caribbean islands. Vet. Parasitol. 1995, 57, 167–176. [Google Scholar] [CrossRef]
- Camus, E.; Maran, M.; Montenegro-James, S.; Accipe, A. Sero-Epidemiological Survey on Bovine Tick-Borne Diseases in the Lesser Antilles. In Proceedings of the Final Research Co-Ordination Meetings of FAO/IAEA/SIDA Co-Ordinated Research Projects, Guadeloupe, Lesser Antilles, France, 13–17 June 1994; International Atomic Energy Agency: Vienna, Austria, 1998; pp. 241–245. [Google Scholar]
- Camus, E.; Montenegro-James, S. Bovine anaplasmosis and babesiosis in the Lesser Antilles: Risk assessment of an unstable epidemiologic situation. Vet. Res. 1994, 25, 313–317. [Google Scholar]
- Zhang, J.; Kelly, P.; Guo, W.; Xu, C.; Wei, L.; Jongejan, F.; Loftis, A.; Wang, C. Development of a generic Ehrlichia FRET-qPCR and investigation of ehrlichioses in domestic ruminants on five Caribbean islands. Parasit. Vectors 2015, 8, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gondard, M.; Cabezas-Cruz, A.; Charles, R.A.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Ticks and Tick-Borne Pathogens of the Caribbean: Current Understanding and Future Directions for More Comprehensive Surveillance. Front. Cell. Infect. Microbiol. 2017, 7, 490. [Google Scholar] [CrossRef] [PubMed]
- Christopher-Hennings, J.; Araujo, K.P.; Souza, C.J.; Fang, Y.; Lawson, S.; Nelson, E.A.; Clement, T.; Dunn, M.; Lunney, J.K. Opportunities for bead-based multiplex assays in veterinary diagnostic laboratories. J. Vet. Diagn. Investig. 2013, 25, 671–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livengood, J.; Hutchinson, M.L.; Thirumalapura, N.; Tewari, D. Detection of Babesia, Borrelia, Anaplasma, and Rickettsia spp. in Adult Black-Legged Ticks (Ixodes scapularis) from Pennsylvania, United States, with a Luminex Multiplex Bead Assay. Vector Borne Zoonotic Dis. 2020, 20, 406–411. [Google Scholar] [CrossRef]
- Reslova, N.; Huvarova, V.; Hrdy, J.; Kasny, M.; Kralik, P. A novel perspective on MOL-PCR optimization and MAGPIX analysis of in-house multiplex foodborne pathogens detection assay. Sci. Rep. 2019, 9, 2719. [Google Scholar] [CrossRef]
- Reslova, N.; Michna, V.; Kasny, M.; Mikel, P.; Kralik, P. xMAP Technology: Applications in Detection of Pathogens. Front. Microbiol. 2017, 8, 55. [Google Scholar] [CrossRef]
- Angeloni, S.D.S.; Dunbar, S.; Stone, V.; Swift, S. xMAP® Cookbook. A Collection of Methods and Protocols for Developing Multiplex Assays with xMAP Technology. Available online: https://cdn2.hubspot.net/hubfs/128032/Cookbook/BR76862.xMAPCookbook.Ed4.WR.pdf (accessed on 9 February 2021).
- Ros-García, A.; Juste, R.A.; Hurtado, A. A highly sensitive DNA bead-based suspension array for the detection and species identification of bovine piroplasms. Int. J. Parasitol. 2012, 42, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Abanda, B.; Paguem, A.; Abdoulmoumini, M.; Kingsley, M.T.; Renz, A.; Eisenbarth, A. Molecular identification and prevalence of tick-borne pathogens in zebu and taurine cattle in North Cameroon. Parasit. Vectors 2019, 12, 448. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.J.; Xu, C.; Lucas, H.; Loftis, A.; Abete, J.; Zeoli, F.; Stevens, A.; Jaegersen, K.; Ackerson, K.; Gessner, A.; et al. Ehrlichiosis, babesiosis, anaplasmosis and hepatozoonosis in dogs from St. Kitts, West Indies. PLoS ONE 2013, 8, e53450. [Google Scholar] [CrossRef] [Green Version]
- Lara, B.; Conan, A.; Thrall, M.A.; Ketzis, J.K.; Branford, G.C.; Rajeev, S. Serologic and Molecular Diagnosis of Anaplasma platys and Ehrlichia canis Infection in Dogs in an Endemic Region. Pathogens 2020, 9, 488. [Google Scholar] [CrossRef]
- Peter, S.G.; Aboge, G.O.; Kariuki, H.W.; Kanduma, E.G.; Gakuya, D.W.; Maingi, N.; Mulei, C.M.; Mainga, A.O. Molecular prevalence of emerging Anaplasma and Ehrlichia pathogens in apparently healthy dairy cattle in peri-urban Nairobi, Kenya. BMC Vet. Res. 2020, 16, 364. [Google Scholar] [CrossRef]
- Tana-Hernández, L.; Navarrete-Arroyo, K.; Ron-Román, J.; Reyna-Bello, A.; Chávez-Larrea, M.A. PCR-diagnosis of Anaplasma marginale in cattle populations of Ecuador and its molecular identification through sequencing of ribosomal 16S fragments. BMC Vet. Res. 2017, 13, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftis, A.D.; Kelly, P.J.; Freeman, M.D.; Fitzharris, S.; Beeler-Marfisi, J.; Wang, C. Tick-borne pathogens and disease in dogs on St. Kitts, West Indies. Vet. Parasitol. 2013, 196, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkey, L.A.; Newton, K.; Brunker, J.; Crowdis, K.; Edourad, E.J.P.; Meneus, P.; Little, S.E. Prevalence of vector-borne pathogens in dogs from Haiti. Vet. Parasitol. 2016, 224, 7–12. [Google Scholar] [CrossRef]
- Carelli, G.; Decaro, N.; Lorusso, A.; Elia, G.; Lorusso, E.; Mari, V.; Ceci, L.; Buonavoglia, C. Detection and quantification of Anaplasma marginale DNA in blood samples of cattle by real-time PCR. Vet. Microbiol. 2007, 124, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Cossío-Bayúgar, R.; Rodríguez, S.D.; García-Ortiz, M.A.; García-Tapia, D.; Aboytes-Torres, R. Bovine anaplasmosis prevalence in northern Veracruz state, Mexico. Prev. Vet. Med. 1997, 32, 165–170. [Google Scholar] [CrossRef]
- da Silva, N.B.; Taus, N.S.; Johnson, W.C.; Mira, A.; Schnittger, L.; Valente, J.D.M.; Vidotto, O.; Masterson, H.E.; Vieira, T.; Ueti, M.W.; et al. First report of Anaplasma marginale infection in goats, Brazil. PLoS ONE 2018, 13, e0202140. [Google Scholar] [CrossRef] [Green Version]
- Torioni de Echaide, S.; Knowles, D.P.; McGuire, T.C.; Palmer, G.H.; Suarez, C.E.; McElwain, T.F. Detection of cattle naturally infected with Anaplasma marginale in a region of endemicity by nested PCR and a competitive enzyme-linked immunosorbent assay using recombinant major surface protein 5. J. Clin. Microbiol. 1998, 36, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, A.; Rahbari, S.; Shayan, P.; Sadeghi-dehkordi, Z.; Bahonar, A. Molecular detection of Anaplasma marginale and Anaplasma ovis in sheep and goat in west highland pasture of Iran. Asian Pac. J. Trop. Biomed. 2017, 7, 455–459. [Google Scholar] [CrossRef]
- Fosgate, G.T.; Urdaz-Rodríguez, J.H.; Dunbar, M.D.; Rae, D.O.; Donovan, G.A.; Melendez, P.; Dobek, G.L.; Alleman, A.R. Diagnostic accuracy of methods for detecting Anaplasma marginale infection in lactating dairy cattle of Puerto Rico. J. Vet. Diagn. Investig. 2010, 22, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Sánchez, A.A.; Meli, M.L.; Obregón Álvarez, D.; Fonseca-Rodríguez, O.; Cabezas-Cruz, A.; Hofmann-Lehmann, R.; Corona-González, B. Development and application of a multiplex TaqMan® real-time qPCR assay for the simultaneous detection of Anaplasma marginale and Theileria annulata and molecular characterization of Anaplasma marginale from cattle in Western Cuba. Ticks Tick Borne Dis. 2020, 11, 101356. [Google Scholar] [CrossRef] [PubMed]
- Obregón, D.; Cabezas-Cruz, A.; Armas, Y.; Silva, J.B.; Fonseca, A.H.; André, M.R.; Alfonso, P.; Oliveira, M.C.S.; Machado, R.Z.; Corona-González, B. High co-infection rates of Babesia bovis, Babesia bigemina, and Anaplasma marginale in water buffalo in Western Cuba. Parasitol. Res. 2019, 118, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Bergval, I.; Sengstake, S.; Brankova, N.; Levterova, V.; Abadía, E.; Tadumaze, N.; Bablishvili, N.; Akhalaia, M.; Tuin, K.; Schuitema, A.; et al. Combined species identification, genotyping, and drug resistance detection of Mycobacterium tuberculosis cultures by MLPA on a bead-based array. PLoS ONE 2012, 7, e43240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuyts, V.; Roosens, N.H.; Bertrand, S.; Marchal, K.; De Keersmaecker, S.C. Guidelines for optimisation of a multiplex oligonucleotide ligation-PCR for characterisation of microbial pathogens in a microsphere suspension array. BioMed Res. Int. 2015, 2015, 790170. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.; Gans, J.; Graves, S.W.; Green, L.; Taylor, L.; Kim, H.B.; Kunde, Y.A.; Leonard, P.M.; Li, P.E.; Mark, J.; et al. A rapid multiplex assay for nucleic acid-based diagnostics. J. Microbiol. Methods 2010, 80, 155–163. [Google Scholar] [CrossRef]
- Thierry, S.; Hamidjaja, R.A.; Girault, G.; Löfström, C.; Ruuls, R.; Sylviane, D. A multiplex bead-based suspension array assay for interrogation of phylogenetically informative single nucleotide polymorphisms for Bacillus anthracis. J. Microbiol. Methods 2013, 95, 357–365. [Google Scholar] [CrossRef]
- Armbruster, D.A.; Pry, T. Limit of blank, limit of detection and limit of quantitation. Clin. Biochem. Rev. 2008, 29 (Suppl. 1), S49–S52. [Google Scholar]
- Sharma, B. Development of a PCR Based Direct DNA Hybridization Oligonucleotide Microbead Assay for Detection of Ehrlichia and Anaplasma Species in Animals and Ticks from Grenada, West Indies. Ph.D. Thesis, St. George’s University, West Indies, Grenada, 2020. [Google Scholar]
- Chang, W.L.; Pan, M.J. Specific amplification of Ehrlichia platys DNA from blood specimens by two-step PCR. J. Clin. Microbiol. 1996, 34, 3142–3146. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.E.; Sumner, J.W.; Dawson, J.E.; Tzianabos, T.; Greene, C.R.; Olson, J.G.; Fishbein, D.B.; Olsen-Rasmussen, M.; Holloway, B.P.; George, E.H.; et al. Detection of the etiologic agent of human ehrlichiosis by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.E.; Greene, C.E.; Jones, D.C.; Dawson, J.E. Ehrlichia ewingii sp. nov., the etiologic agent of canine granulocytic ehrlichiosis. Int. J. Syst. Bacteriol. 1992, 42, 299–302. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| DNA Targets | MFI for Hybridization of Species-Specific Oligonucleotides to DNA Targets | |||||
|---|---|---|---|---|---|---|
| A. marginale | E. canis | E. ewingii | A. platys | E. chaffeensis | E. ruminantium | |
| NTCs c | 23.5 ± 11.1 a (>34.6) | 24 ± 14.1 a (>38.5) | 23.1 ± 11.4 a (>35) | 24.6 ± 10.8 a (>35.8) | 23.1 ± 10.2 a (>33.5) | 25.5 ± 10.2 a (>36) |
| A. marginale # | 670.5b ± 118.6 | 21.6 ± 3.7 | 24 ± 2.6 | 22 ± 2.6 | 21.6 ± 4.6 | 24 ± 3.4 |
| E. canis # | 21.6 ± 1.1 | 2088.6b ± 92.5 | 21.6 ± 2 | 22 ± 2 | 22 ± 1 | 24 ± 1.7 |
| E. ewingii # | 18.3 ± 1.5 | 20.3 ± 0.5 | 1605.3b ± 147 | 20.6 ± 2.5 | 19.3 ± 1.1 | 21 ± 2 |
| A. platys # | 21 ± 2.6 | 22.8 ± 3.1 | 21.6 ± 1.1 | 1870.2b ± 270 | 21.3 b ± 3.2 | 24.6 ± 3 |
| E. chaffeensis # | 23.5 ± 2.3 | 25.3 ± 2 | 24 ± 1.7 | 23.6 ± 1.1 | 2715.8b ± 321.4 | 27 ± 1.7 |
| E. ruminantium # | 20.3 ± 1.5 | 22.3 ± 2.3 | 21.6 ± 1.5 | 21 ± 1.7 | 19.3 ± 2 | 828b ± 83.9 |
| DNA Targets | MFI for Hybridization of Species-Specific Oligonucleotides to DNA Targets | |||||
|---|---|---|---|---|---|---|
| A. marginale | E. canis | E. ewingii | A. platys | E. chaffeensis | E. ruminantium | |
| NTC c | 19.2 ± 4.5 a (>23.7) | 20.3 ± 1.5 a (>21.8) | 20.3 ± 1.5 a (>21.8) | 21.1 ± 4.2 a (>25.3) | 19.5 ± 1.5 a (>21) | 22 ± 2.4 a (>24.4) |
| Neg D d | 17.8 ± 0.5 | 19 ± 0 | 18 ± 0.8 | 19.2 ± 0.5 | 18 ± 0.8 | 20.5 ± 1 |
| Neg C e | 19.5 ± 0.5 | 19.8 ± 0.9 | 19.2 ± 0.9 | 20 ± 0.8 | 19.2 ± 0.5 | 21.5 ± 2 |
| EC-AP f | 20 ± 0 | 1344.2b ± 16.2 | 20.5 ± 0.5 | 331.1b ± 7.3 | 20.8 ± 1.7 | 23.6 ± 1.3 |
| AP-EC g | 18.9 ± 0.2 | 36b ± 2.1 | 19.8 ± 1.2 | 1242.6b ± 39.3 | 20.2 ± 2.3 | 21.6 ± 1.7 |
| EC-ECH h | 21.2 ± 0.9 | 1500.5b ± 19.2 | 22.8b ± 1.8 | 22 ± 0.8 | 55b ± 0 | 24.5b ± 1.2 |
| ECH-EC i | 18.8 ± 1.7 | 28.2b ± 2.6 | 18.5 ± 2 | 18.9 ± 1.6 | 2231.6b ± 80.9 | 20.4 ± 1.7 |
| AM-ECH j | 1077.6b ± 26.8 | 20 ± 0.8 | 25.2b ± 0.9 | 19.8 ± 0.5 | 107.5b ± 2.6 | 21.8 ± 0.5 |
| ECH-AM k | 25.8b ± 1.7 | 26.2b ± 1.2 | 23.5b ± 0.5 | 24.9 ± 1 | 3997b ± 120.8 | 27b ± 2 |
| Species/# Sampled | # of Samples | Parish (# Sampled) | Year of Collection | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SG | SA | SM | SP | SD | SJ | Uk | |||
| Canine/353 | 358 * | 185 | 48 | 12 | 4 | 50 | 1 | 58 | 2014–2018 |
| Caprine and Ovine/65 | 65 | 18 | 25 | - | - | 22 | - | - | 2017–2018 |
| Bovine/32 | 32 | 25 | 7 | - | - | - | - | - | 2017 |
| Total (%) | 455 | 228 (50.1) | 80 (17.6) | 12 (2.6) | 4 (0.8) | 72 (15.8) | 1 (0.2) | 58 (12.7) | |
| Bacterial sp. | Canine | Bovine | Caprine and Ovine | Total | ||
|---|---|---|---|---|---|---|
| Animal sp. | ||||||
| Single speciesinfections | AM | - | 11/32 (34.3) | 3/65 (4.6) | 14 | |
| EC | 50/358 (13.9) | - | 1/65 (1.5) | 51 | ||
| AP | 59/358 (16.4) | 3/32 (9.3) | 2/65 (3) | 64 | ||
| ECH | 1/358 (0.2) | - | - | 1 | ||
| Total | 110/358 (30.7) | 14/32 (43.7) | 6/65 (9.2) | 130 | ||
| Co-infections | EC-ECH | 2/358 (0.5) | - | - | 2 | |
| EC-AP | 4/358 (1.1) | - | - | 4 | ||
| EE-AP | 1/358 (0.2) | - | - | 1 | ||
| Total | 7/358 (1.9) | 0/36 (0) | 0/65 (0) | 7 | ||
| Grand Total | 117/358 (32.6) | 14/32 (43.7) | 6/65 (9.2) | 137 | ||
| Species | Target Gene | # of Samples Tested | # of Samples Sequenced | Deposited Sequence GenBank #s | Length (bp) | Percentage of Identity (%) | Reference Sequence |
|---|---|---|---|---|---|---|---|
| A. marginale | msp1a | 8 | 2 | MW486117 MW486118 | 568 326 | 94.00 94.00 | NC_012026 NC_012026 |
| E. canis | 16S rRNA | 6 | 2 | MW474807 MW474808 | 335 335 | 99.40 99.40 | NR_118741 NR_118741 |
| E. chaffeensis | 16S rRNA | 10 | 6 | MW474809 MW474810 MW474811 MW474812 MW474813 MW474814 | 300 334 318 334 333 334 | 100.00 100.00 100.00 100.00 100.00 100.00 | NR_074500 NR_074500 NR_074500 NR_074500 NR_074500 NR_074500 |
| E. ewingii | 16S rRNA | 6 | 1 | MW474815 | 308 | 100.00 | NR_074500 |
| Optimization Conditions | Test Conditions | |
|---|---|---|
| PCR optimization | Primer Annealing temperature (°C) | 50, 52 |
| MgCl2 concentration (mM) | 1.5, 2.5 | |
| PCR cycle numbers | 30, 35, 40 | |
| xMAP optimization | Hybridization Temperature (°C) | 50, 52, 55, 60 |
| Hybridization time (min.) | 10, 15, 20 | |
| PCR product volume (µL) | 5, 10 | |
| xMAP protocol | Washed, no-wash | |
| Concentration of the probes (nmol/µL) | 0.1, 0.2 |
| Probes | Bacterial Species | Sequences (5′-3′) | xMAP COOH-Microsphere Regions for Probe Binding | Reference |
|---|---|---|---|---|
| RG270Ecan | E. canis | TATAGCCTCTGGCTATAGGAAATTGTTAG | R25 | [37] |
| RG266Echaf | E. chaffeensis | CTTATAACCTTTTGGTTATAAATAATTGTTAG | R43 | [37] |
| RG268Eewin | E. ewingii | CTAAATAGTCTCTGATTTAGATAGTTGTTAG | R34 | [37] |
| RG260Erum | E. ruminantium | GTTATTTATAGCTTCGGCTAT | R48 | [37] |
| RG272Aplat | A. platys | CGGATTTTTGTCGTAGCTTGCTATGAT | R38 | [37] |
| RG262Amarg | A. marginale | CGTATACGCAGCTTGCTGCGT | R20 | This study |
| Species | Mix | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 ✴ | 2 ✴ | 3 ✴ | 4 ✴ | 5 ✴ | 6 ✴ | 7 and 8 ★ | 9 and 10 ★ | 11 and 12 ★ | |
| E. canis | ✓ | ✗ | ✗ | ✗ | ✗ | ✗ | ✓ | ✓ | ✗ |
| E. chaffeensis | ✗ | ✓ | ✗ | ✗ | ✗ | ✗ | ✗ | ✓ | ✓ |
| E. ewingii | ✗ | ✗ | ✓ | ✗ | ✗ | ✗ | ✗ | ✗ | ✗ |
| E. ruminantium | ✗ | ✗ | ✗ | ✓ | ✗ | ✗ | ✗ | ✗ | ✗ |
| A. platys | ✗ | ✗ | ✗ | ✗ | ✓ | ✗ | ✓ | ✗ | ✗ |
| A. marginale | ✗ | ✗ | ✗ | ✗ | ✗ | ✓ | ✗ | ✗ | ✓ |
| Species | Target Gene | Primer Name | Sequence (5′→3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| A. marginale | msp1a | MSP1aF1 MSP1aRN | GCATTACAACGCAACGCTTGAG CAGGAGCACCACCAAACATCATCACA | 1638 | This study |
| A. platys | 16S rRNA | EP2 EP3 | GAAGATAATGACGGTACCC CGTTTTGTCTCTGTGTTG | 385 | [83] |
| E. canis | 16S rRNA | ECA HE3 | CAATTATTTATAGCCTCTGGCTATAGG TATAGGTACCGTCATTATCTTCCCTAT | 385 | [34] |
| E. chaffeensis | 16S rRNA | HE1 HE3 | CAATTGCTTATAACCTTTTGGTTATAAAT TATAGGTACCGTCATTATCTTCCCTAT | 385 | [84] |
| E. ewingii | 16S rRNA | EE72 HE3 | CAATTCCTAAATAGTCTCTGACTATT TATAGGTACCGTCATTATCTTCCCTAT | 385 | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, B.; Ganta, R.R.; Stone, D.; Alhassan, A.; Lanza-Perea, M.; Matthew Belmar, V.; Karasek, I.; Cooksey, E.; Butler, C.M.; Gibson, K.; et al. Development of a Multiplex PCR and Magnetic DNA Capture Assay for Detecting Six Species Pathogens of the Genera Anaplasma and Ehrlichia in Canine, Bovine, Caprine and Ovine Blood Samples from Grenada, West Indies. Pathogens 2021, 10, 192. https://doi.org/10.3390/pathogens10020192
Sharma B, Ganta RR, Stone D, Alhassan A, Lanza-Perea M, Matthew Belmar V, Karasek I, Cooksey E, Butler CM, Gibson K, et al. Development of a Multiplex PCR and Magnetic DNA Capture Assay for Detecting Six Species Pathogens of the Genera Anaplasma and Ehrlichia in Canine, Bovine, Caprine and Ovine Blood Samples from Grenada, West Indies. Pathogens. 2021; 10(2):192. https://doi.org/10.3390/pathogens10020192
Chicago/Turabian StyleSharma, Bhumika, Roman R. Ganta, Diana Stone, Andy Alhassan, Marta Lanza-Perea, Vanessa Matthew Belmar, Inga Karasek, Elizabeth Cooksey, Catherine M. Butler, Kathryn Gibson, and et al. 2021. "Development of a Multiplex PCR and Magnetic DNA Capture Assay for Detecting Six Species Pathogens of the Genera Anaplasma and Ehrlichia in Canine, Bovine, Caprine and Ovine Blood Samples from Grenada, West Indies" Pathogens 10, no. 2: 192. https://doi.org/10.3390/pathogens10020192
APA StyleSharma, B., Ganta, R. R., Stone, D., Alhassan, A., Lanza-Perea, M., Matthew Belmar, V., Karasek, I., Cooksey, E., Butler, C. M., Gibson, K., & Wilkerson, M. J. (2021). Development of a Multiplex PCR and Magnetic DNA Capture Assay for Detecting Six Species Pathogens of the Genera Anaplasma and Ehrlichia in Canine, Bovine, Caprine and Ovine Blood Samples from Grenada, West Indies. Pathogens, 10(2), 192. https://doi.org/10.3390/pathogens10020192