
Inhibition of Dermatophyte Fungi by Australian Jarrah Honey

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Minimum Inhibitory Concentrations (MICs) and Minimum Antifungal Concentrations (MFCs) for Dermatophytes Treated with Jarrah, Leptospermum and Artificial Honey
2.2. Estimation of H2O2 Production by Jarrah (Barnes 10+) Honey
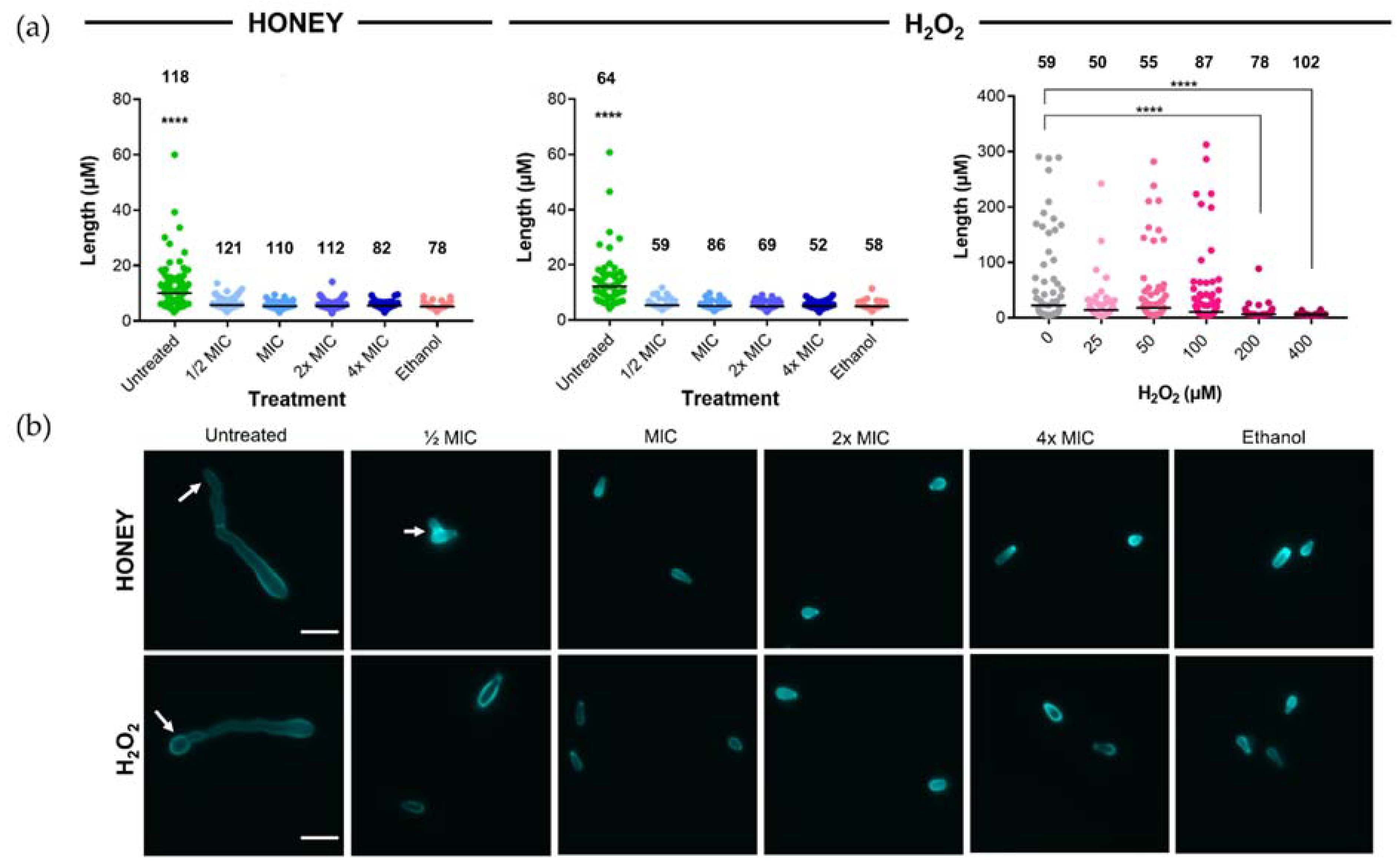
2.3. Microscopic Examination of the Effect of Jarrah Honey on T. rubrum Conidia and Their Germination
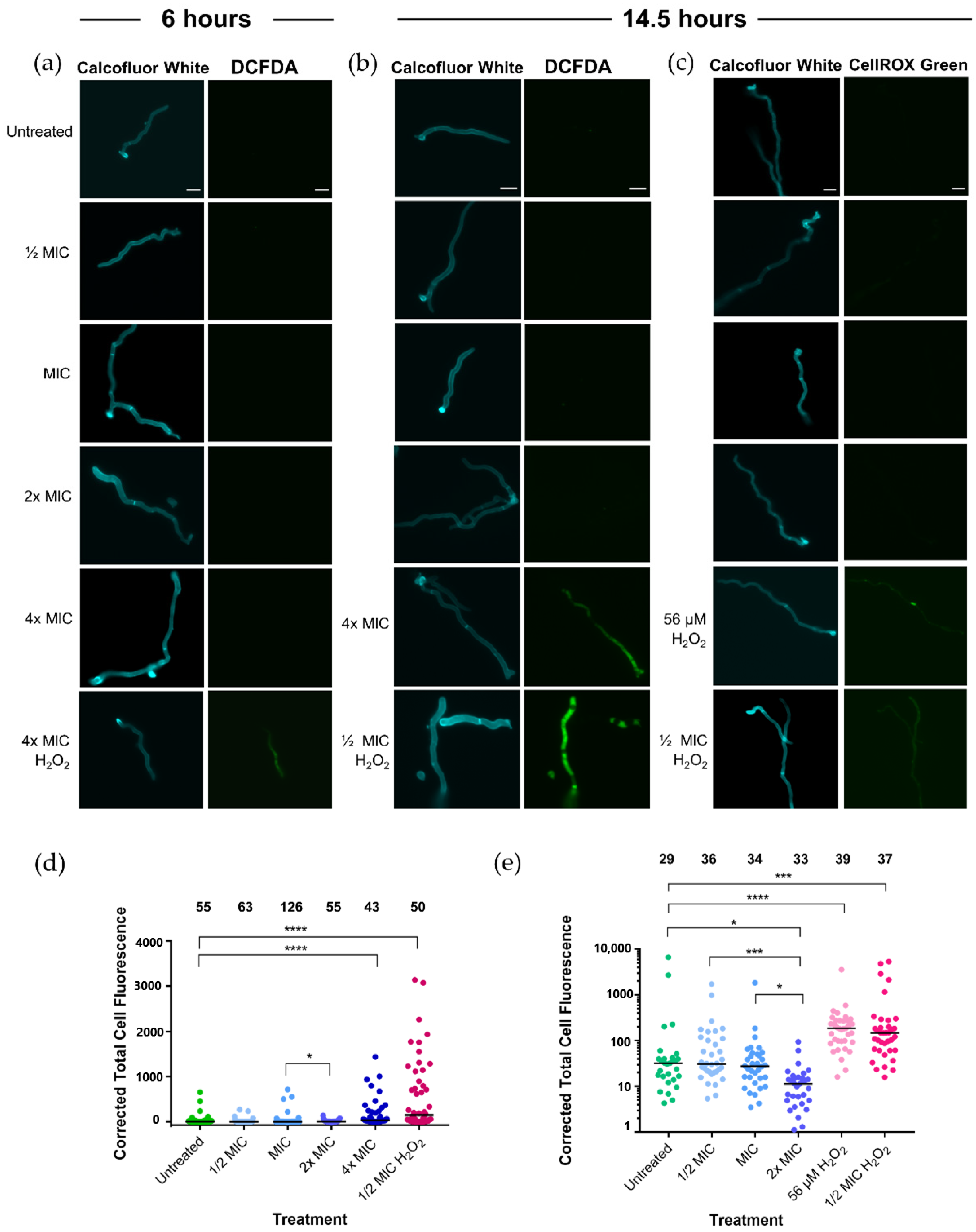
2.4. Analysis of Oxidative Stress in Fungal Hyphae Following Treatment with Jarrah (Barnes 10+) Using DCFDA and CellROX Green
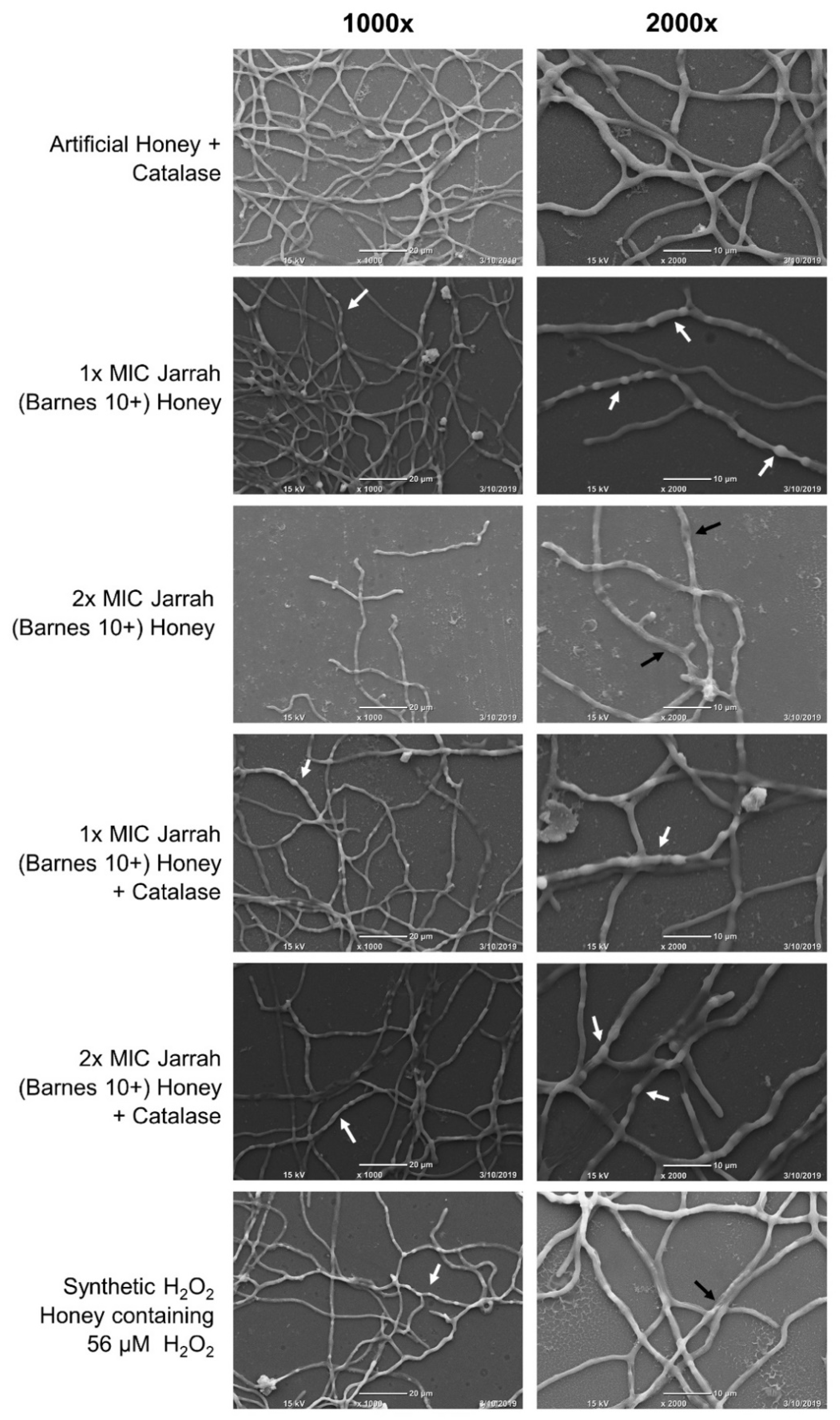
2.5. Analysis of T. rubrum Hyphae Treated with Jarrah Honey by Scanning Electron Microscopy
3. Discussion
3.1. Many Fungal Dermatophyte Species Are Highly Susceptible to Jarrah Honey
3.2. H2O2 Production Is Necessary but Not Sufficient for the Inhibition of Dermatophyte Fungi by Jarrah Honey
3.3. The Antifungal Activity of Jarrah Honey Appears to Be Mediated on the Surface of T. rubrum Hyphae, Causing Deformities and Hyphal Collapse
4. Conclusions
5. Materials and Methods
5.1. Dermatophyte Cultures
5.2. Honey Samples
5.3. Quantification of H2O2 Production
5.4. Honey Component Solutions
5.5. Minimum Inhibitory Concentrations (MICs) and Minimum Fungicidal Concentrations (MFCs) of Honey
5.6. Microscopic Analysis of T. rubrum Conidia Treated with Honey
5.7. Detection of Reactive Oxygen Species in T. rubrum Hyphae Treated with Jarrah Honey
5.8. Scanning Electron Microscopy of T. rubrum Hyphae Treated with Jarrah Honey
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Graser, Y.; Scott, J.; Summerbell, R. The new species concept in dermatophytes—A polyphasic approach. Mycopathologia 2008, 166, 239–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elewski, B.E.; Rich, P.; Tosti, A.; Pariser, D.M.; Scher, R.; Daniel, R.C.; Gupta, A.K. Onchomycosis: An overview. J. Drugs Dermatol. 2013, 12, 96–103. [Google Scholar]
- Havlickova, B.; Czaika, V.A.; Friedrich, M. Epidemiological trends in skin mycoses worldwide. Mycoses 2008, 51, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Achterman, R.R.; White, T.C. A foot in the door for dermatophyte research. PLoS Path. 2012, 8, e1002564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, I.P.; Kakkar, S. Topical delivery of antifungal agents. Expert Opin. Drug Deliv. 2010, 7, 1303–1327. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Chow, M.; Daniel, C.R.; Aly, R. Treatments of tinea pedis. Dermatol. Clin. 2003, 21, 431–462. [Google Scholar] [CrossRef]
- Burlando, B.; Cornara, L. Honey in dermatology and skin care: A review. J. Cosmet. Dermatol. 2013, 12, 306–313. [Google Scholar] [CrossRef]
- Irish, J.; Carter, D.A.; Shokohi, T.; Blair, S.E. Honey has an antifungal effect against Candida species. Med. Mycol. 2006, 44, 289–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Vandenbroucke-Grauls, C.M.J.E.; Zaat, S.A.J. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molan, P.C. The antibacterial activity of honey: 2. Variation in the potency of the antibacterial activity. N. Z. Bee World 1992, 73, 59–76. [Google Scholar] [CrossRef]
- Carter, D.A.; Blair, S.E.; Cokcetin, N.N.; Bouzo, D.; Brooks, P.; Schothauer, R.; Harry, E.J. Therapeutic manuka honey: No longer so alternative. Front. Microbiol. 2016, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwakman, P.H.S.; Zaat, S.A.J. Antibacterial components of honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef]
- Brudzynski, K.; Miotto, D. Honey melanoidins: Analysis of the compositions of the high molecular weight melanoidins exhibiting radical-scavenging activity. Food Chem. 2011, 127, 1023–1030. [Google Scholar] [CrossRef]
- Weston, R.J. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chem. 2000, 71, 235–239. [Google Scholar] [CrossRef]
- White, J.W.; Subers, M.H.; Schepartz, A.I. The identification of inhibine, the antibacterial factor in honey, as hydrogen peroxide and its origin in a honey glucose-oxidase system. BBA 1963, 73, 57–70. [Google Scholar] [CrossRef]
- Adams, C.J.; Manley-Harris, M.; Molan, P.C. The origin of methylglyoxal in New Zealand manuka (Leptospermum scoparium) honey. Carbohydr. Res. 2009, 344, 1050–1053. [Google Scholar] [CrossRef] [PubMed]
- Cokcetin, N.; Pappalardo, M.; Campbell, L.T.; Brooks, P.; Carter, D.A.; Blair, S.E.; Harry, E.J. The antibacterial activity of Australian Leptospermum honey correlates with methylglyoxal levels. PLoS ONE 2016, 11, e0167780. [Google Scholar] [CrossRef]
- Allen, K.L.; Molan, P.C.; Reid, G.M. A survey of the antibacterial activity of some New Zealand honeys. J. Pharm. Pharmacol. 1991, 43, 817–822. [Google Scholar] [CrossRef]
- Brady, N.F.; Molan, P.C.; Harfoot, C.G. The sensitivity of dermatophytes to the antimicrobial activity of manuka honey and other honey. Pharm. Pharmacol. Commun. 1996, 2, 471–473. [Google Scholar] [CrossRef]
- Katiraee, F.; Mahmodi, R.; Mardani, K.; Babaei, E. Antifungal activity of Iranian honeybees against Candida, Aspergillus species and Trichophyton rubrum. J. Food Process. Preserv. 2014, 38, 2078–2082. [Google Scholar] [CrossRef]
- Irish, J.; Blair, S.; Carter, D.A. The antibacterial activity of honey derived from Australian flora. PLoS ONE 2011, 6, e18229. [Google Scholar] [CrossRef]
- Guttentag, A.; Krishnakumar, K.; Cokcetin, N.; Harry, E.; Carter, D.A. Factors affecting the production and measurement of hydrogen peroxide in honey samples. Access Microbiol. 2021. [Google Scholar] [CrossRef]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef]
- Yagi, H.; Tan, J.; Tuan, R.S. Polyphenols suppress hydrogen peroxide-induced oxidative stress in human bone-marrow derived mesenchymal stem cells. J. Cell. Biochem. 2013, 114, 1163–1173. [Google Scholar] [CrossRef]
- Lehmann, D.M.; Krishnakumar, K.; Batres, M.A.; Hakola-Parry, A.; Cokcetin, N.; Harry, E.; Carter, D.A. A cost-effective colourimetric assay for quantifying hydrogen peroxide in honey. Access Microbiol. 2019, 1. [Google Scholar] [CrossRef]
- Anyanwu, C. Investigation of in vitro antifungal activity of honey. JMPR 2012, 6, 3512–3516. [Google Scholar] [CrossRef]
- Brady, N.; Molan, P.; Bang, L. A survey of non-manuka New Zealand honeys for antibacterial and antifungal activities. J. Apic. Res. 2004, 43, 47–52. [Google Scholar] [CrossRef]
- Anand, S.; Deighton, M.; Livanos, G.; Pang, E.C.K.; Mantri, N. Agastache honey has superior antifungal activity in comparison with important commercial honeys. Sci. Rep. 2019, 9, 18197. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Campbell, L.T.; Blair, S.E.; Carter, D.A. The effect of standard heat and filtration processing procedures on antimicrobial activity and hydrogen peroxide levels in honey. Front. Microbiol. 2012, 3, 265. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Abubaker, K.; Wang, T. Powerful bacterial killing by buckwheat honeys is concentration-dependent, involves complete DNA degradation and requires hydrogen peroxide. Front. Microbiol. 2012, 3, 242. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Buriova, M.; Pekarik, L.; Majtan, V.; Majtan, J. Phytochemicals-mediated production of hydrogen peroxide is crucial for high antibacterial activity of honeydew honey. Sci. Rep. 2018, 8, 9061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, S.; Pang, E.; Livanos, G.; Mantri, N. Characterization of physico-chemical properties and antioxidant capacities of bioactive honey produced from Australian grown Agastache rugosa and its correlation with colour and poly-phenol content. Molecules 2018, 23, 108. [Google Scholar] [CrossRef] [Green Version]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Speijer, D.; Vandenbroucke-Grauls, C.; Zaat, S.A.J. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, K.; Miotto, D. The relationship between the content of Maillard reaction-like products and bioactivity of Canadian honeys. Food Chem. 2011, 124, 869–874. [Google Scholar] [CrossRef]
- Masoura, M.; Passaretti, P.; Overton, T.W.; Lund, P.A.; Gkatzionis, K. Use of a model to understand the synergies underlying the antibacterial mechanism of H2O2-producing honeys. Sci. Rep. 2020, 10, 17692. [Google Scholar] [CrossRef] [PubMed]
- Jansen, T.; Borgers, M.; van de Ven, M.A.; Xhonneux, B.; Wouters, L.; van Gerven, F.; van Cutsem, J. The effects of saperconazole on the morphology of Candida albicans, Pityrosporum ovale and Trichophyton rubrum in vitro. J. Med. Vet. Mycol. 1991, 29, 293–303. [Google Scholar] [CrossRef]
- Smijs, T.G.M.; Mulder, A.A.; Pavel, S.; Onderwater, J.J.M.; Koerten, H.K.; Bouwstra, J.A. Morphological changes of the dermatophyte Trichophyton rubrum after photodynamic treatment: A scanning electron microscopy study. Med. Mycol. 2008, 46, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.; Isham, N.; Henry, W.; Kroon, H.A.; Yurdakul, S. Evaluation of the morphological effects of TDT 067 (terbinafine in transfersome) and conventional terbinafine on dermatophyte hyphae in vitro and in vivo. Antimicrob. Agents Chemother. 2012, 56, 2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriques, A.F.; Jenkins, R.E.; Burton, N.F.; Cooper, R.A. The effect of manuka honey on the structure of Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Nishio, E.K.; Ribeiro, J.M.; Oliveira, A.G.; Andrade, C.G.T.J.; Proni, E.A.; Kobayashi, R.K.T.; Nakazato, G. Antibacterial synergic effect of honey from two stingless bees: Scaptotrigona bipunctata Lepeletier, 1836, and S. postica Latreille, 1807. Sci. Rep. 2016, 6, 21641. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, J.; Gao, H.; Wang, Z.; Dong, N.; Ma, Q.; Shan, A. Antimicrobial properties and membrane-active mechanism of a potential α-helical antimicrobial derived from cathelicidin PMAP-36. PLoS ONE 2014, 9, e86364. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, K.E.; Weeks, K.; Carter, D.A. Lactoferrin is broadly active against yeasts and highly synergistic with amphotericin B. Antimicrob. Agents Chemother. 2020, 64, e02284-19. [Google Scholar] [CrossRef] [Green Version]
- Sokol-Anderson, M.L.; Brajtburg, J.; Medoff, G. Amphotericin B-induced oxidative damage and killing of Candida albicans. J. Infect. Dis. 1986, 154, 76–83. [Google Scholar] [CrossRef]
- Sangalli-Leite, F.; Scorzoni, L.; Mesa-Arango, A.C.; Casas, C.; Herrero, E.; Soares Mendes Gianinni, M.J.; Rodríguez-Tudela, J.L.; Cuenca-Estrella, M.; Zaragoza, O. Amphotericin B mediates killing in Cryptococcus neoformans through the induction of a strong oxidative burst. Microb. Infect. 2011, 13, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.-W.; Pang, C.N.I.; Campbell, L.T.; Chen, S.C.A.; Wilkins, M.R.; Carter, D.A. Different pathways mediate amphotericin-lactoferrin drug synergy in Cryptococcus and Saccharomyces. Front. Microbiol. 2019, 10, 2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, C.N.; Lai, Y.W.; Campbell, L.T.; Chen, S.C.; Carter, D.A.; Wilkins, M.R. Transcriptome and network analyses in Saccharomyces cerevisiae reveal that amphotericin B and lactoferrin synergy disrupt metal homeostasis and stress response. Sci. Rep. 2017, 7, 40232. [Google Scholar] [CrossRef] [Green Version]
- de Castro, P.A.; Savoldi, M.; Bonatto, D.; Barros, M.H.; Goldman, M.H.; Berretta, A.A.; Goldman, G.H. Molecular characterization of propolis-induced cell death in Saccharomyces cerevisiae. Eukaryot. Cell 2011, 10, 398–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, P.; Haza, A.I. Antiproliferative and apoptotic effects of Spanish honeys. Pharmacogn. Mag. 2013, 9, 231–237. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous fungi: Approved Standard, 2nd ed.; CLSI document M28-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Nenoff, P.; Krüger, C.; Ginter-Hanselmayer, G.; Tietz, H.J. Mycology—An update. Part 1: Dermatomycoses: Causative agents, epidemiology and pathogenesis. J. Dtsch. Dermatol. Ges. 2014, 12. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Honey (% (w/v)) | VOR 3 MIC80 (µg/mL) | |||||
|---|---|---|---|---|---|---|---|
| Jarrah 2017 | Leptospermum | Artificial | |||||
| MIC80 1 | MFC 2 | MIC80 | MFC | MIC80 | MFC | ||
| Microsporum canis | 1.5 | 2 | 7.5 | 10 | 20 | >50 | 0.023 |
| Microsporum nanum | 1.5 | 2 | 10 | 10 | 30 | >50 | 0.031 |
| Nannizzia gypsea | 3.5 | 5 | 15 | 25 | 40 | >50 | 0.094 |
| Trichophyton interdigitale | 3.5 | 5 | 15 | 17.5 | 40 | >50 | 0.125 |
| Trichophyton rubrum | 2.5 | 4 | 10 | 12.5 | 30 | >50 | 0.031 |
| Trichophyton tonsurans | 2.5 | 3.5 | 10 | 10 | 30 | >50 | 0.125 |
| MIC100 [% (w/v)] | ||||||
|---|---|---|---|---|---|---|
| Jarrah (Barnes 10+) | Jarrah 10/13 | Stringybark 19 | ||||
| Catalase Treatment | − | + | − | + | − | + |
| Nannizzia gypsea | 3.1 | 25 | 1.56 | 25 | 3.1 | 25 |
| Trichophyton interdigitale | 3.1 | ≥25 | 1.56 | ≥25 | 4.2 ± 1.0 | >25 |
| Trichophyton rubrum | 1.56 | 25 | 1.56 | 25 | 1.56 | 25 |
| Honey Sample | Floral Source | Active Component | Maximum Hydrogen Peroxide (H2O2) Production (mM) | Origin |
|---|---|---|---|---|
| Jarrah 10 | Eucalyptus marginata | H2O2 | 2.86 ± 0.31 | [21] |
| Jarrah 13 | Eucalyptus marginata | H2O2 | 3.84 ± 0.24 | [21] |
| Stringybark 19 | Eucalyptus species | H2O2 | 0.93 ± 0.12 | [21] |
| Leptospermum 2 | Leptospermum speciosum | MGO | – | UTS honey collection |
| Jarrah (Barnes 10+) | Eucalyptus marginata | H2O2 | 0.136 ± 0.01 | Commercial |
| Jarrah 2017 | Eucalyptus marginata | H2O2 | 1.59 ± 0.18 | Capilano |
| Honey Solution | Final Concentration of Components in RPMI-1640 | Figures | |||
|---|---|---|---|---|---|
| Artificial Honey | Jarrah (Barnes 10+) Honey | H2O2 | Catalase | ||
| [% (w/v)] | [% (w/v)] | [µM] | +/– | ||
| Control | |||||
| Untreated | 10 | 0 | 0 | – | Figure 1, Figure 2 |
| Jarrah (Barnes 10+) + Artificial Honey | |||||
| ½× MIC | 9.22 | 0.78 | 0 | – | Figure 1, Figure 2 |
| 1× MIC | 8.44 | 1.56 | 0 | – | Figure 1, Figure 2, Figure 3 |
| 2× MIC | 6.88 | 3.12 | 0 | – | Figure 1, Figure 2, Figure 3 |
| 4× MIC | 3.76 | 6.24 | 0 | – | Figure 1, Figure 2a,b,d |
| Synthetic H2O2 Honey | |||||
| ½× MIC | 10 | 0 | 215 | – | Figure 1, Figure 2b–e |
| 1× MIC | 10 | 0 | 430 | – | Figure 1 |
| 2× MIC | 10 | 0 | 860 | – | Figure 1 |
| 4× MIC | 10 | 0 | 1720 | – | Figure 1, Figure 2a |
| 0 µM | 10 | 0 | 0 | – | Figure 1a |
| 25 µM | 10 | 0 | 25 | – | Figure 1a |
| 50 µM | 10 | 0 | 50 | – | Figure 1a |
| 100 µM | 10 | 0 | 100 | – | Figure 1a |
| 200 µM | 10 | 0 | 200 | – | Figure 1a |
| 400 µM | 10 | 0 | 400 | – | Figure 1a |
| 56 µM H2O2 | 10 | 0 | 56 | – | Figure 2c,e, Figure 3 |
| Jarrah (Barnes 10+) + Catalase + Artificial Honey | |||||
| Control (Artificial honey + Catalase) | 10 | 0 | 0 | + | Figure 3 |
| 1× MIC Jarrah (Barnes 10+) Honey + Catalase | 8.44 | 1.56 | 0 | + | Figure 3 |
| 2× MIC Jarrah (Barnes 10+) Honey + Catalase | 6.88 | 3.12 | 0 | + | Figure 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guttentag, A.; Krishnakumar, K.; Cokcetin, N.; Hainsworth, S.; Harry, E.; Carter, D. Inhibition of Dermatophyte Fungi by Australian Jarrah Honey. Pathogens 2021, 10, 194. https://doi.org/10.3390/pathogens10020194
Guttentag A, Krishnakumar K, Cokcetin N, Hainsworth S, Harry E, Carter D. Inhibition of Dermatophyte Fungi by Australian Jarrah Honey. Pathogens. 2021; 10(2):194. https://doi.org/10.3390/pathogens10020194
Chicago/Turabian StyleGuttentag, Annabel, Krishothman Krishnakumar, Nural Cokcetin, Steven Hainsworth, Elizabeth Harry, and Dee Carter. 2021. "Inhibition of Dermatophyte Fungi by Australian Jarrah Honey" Pathogens 10, no. 2: 194. https://doi.org/10.3390/pathogens10020194
APA StyleGuttentag, A., Krishnakumar, K., Cokcetin, N., Hainsworth, S., Harry, E., & Carter, D. (2021). Inhibition of Dermatophyte Fungi by Australian Jarrah Honey. Pathogens, 10(2), 194. https://doi.org/10.3390/pathogens10020194