
Zur: Zinc-Sensing Transcriptional Regulator in a Diverse Set of Bacterial Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Zinc: An Indispensable Transition Metal
1.1. Zn and Bacterial Pathogenesis
1.2. Maintenance of Zn Homeostasis Is Crucial
2. Role of Zn in the Host-Pathogen Interaction
2.1. Role of Zn in the Host
2.2. A Host–Pathogen Tug of War for Zn
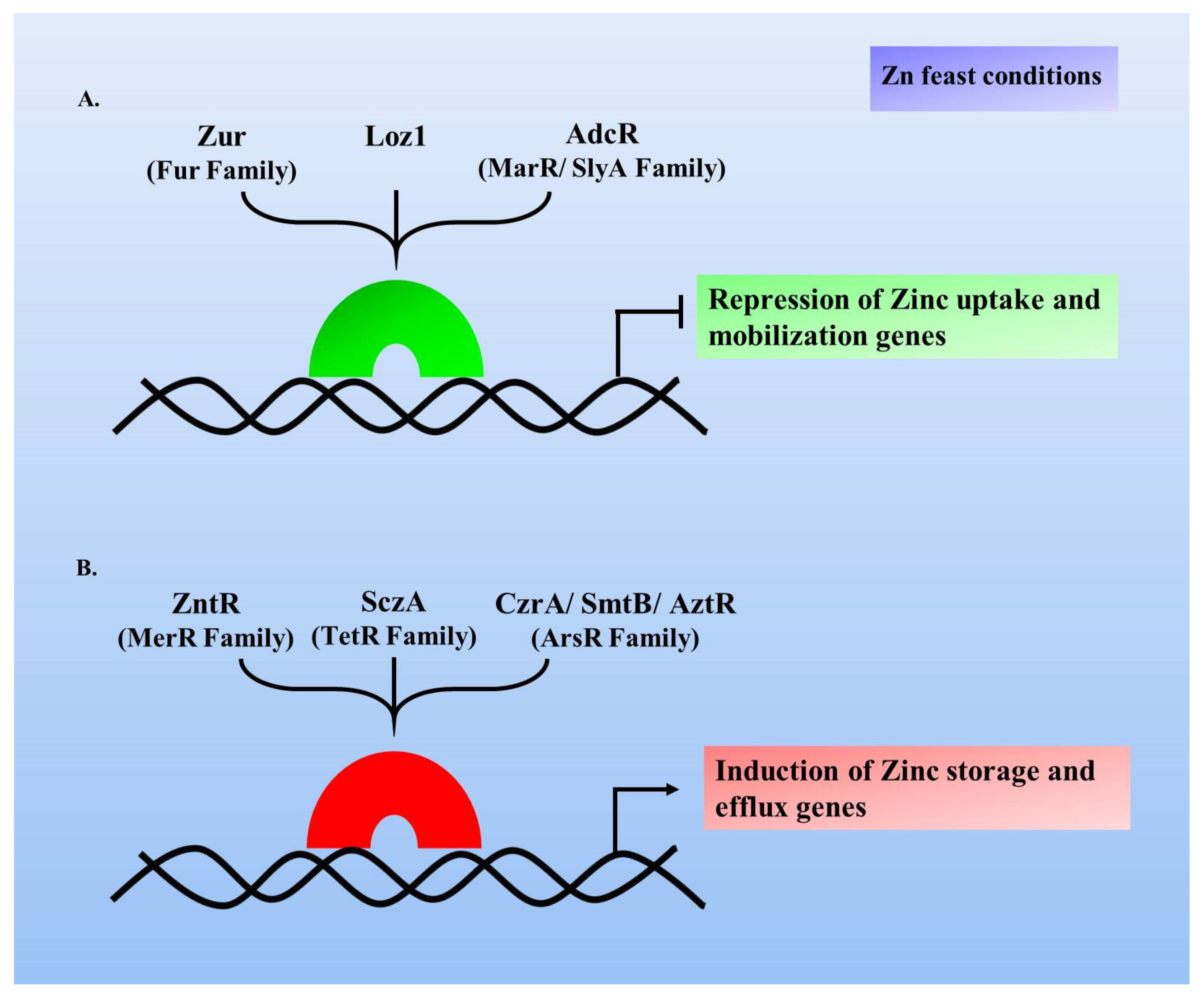
3. Zn Homeostasis Commences at the Transcriptional Level
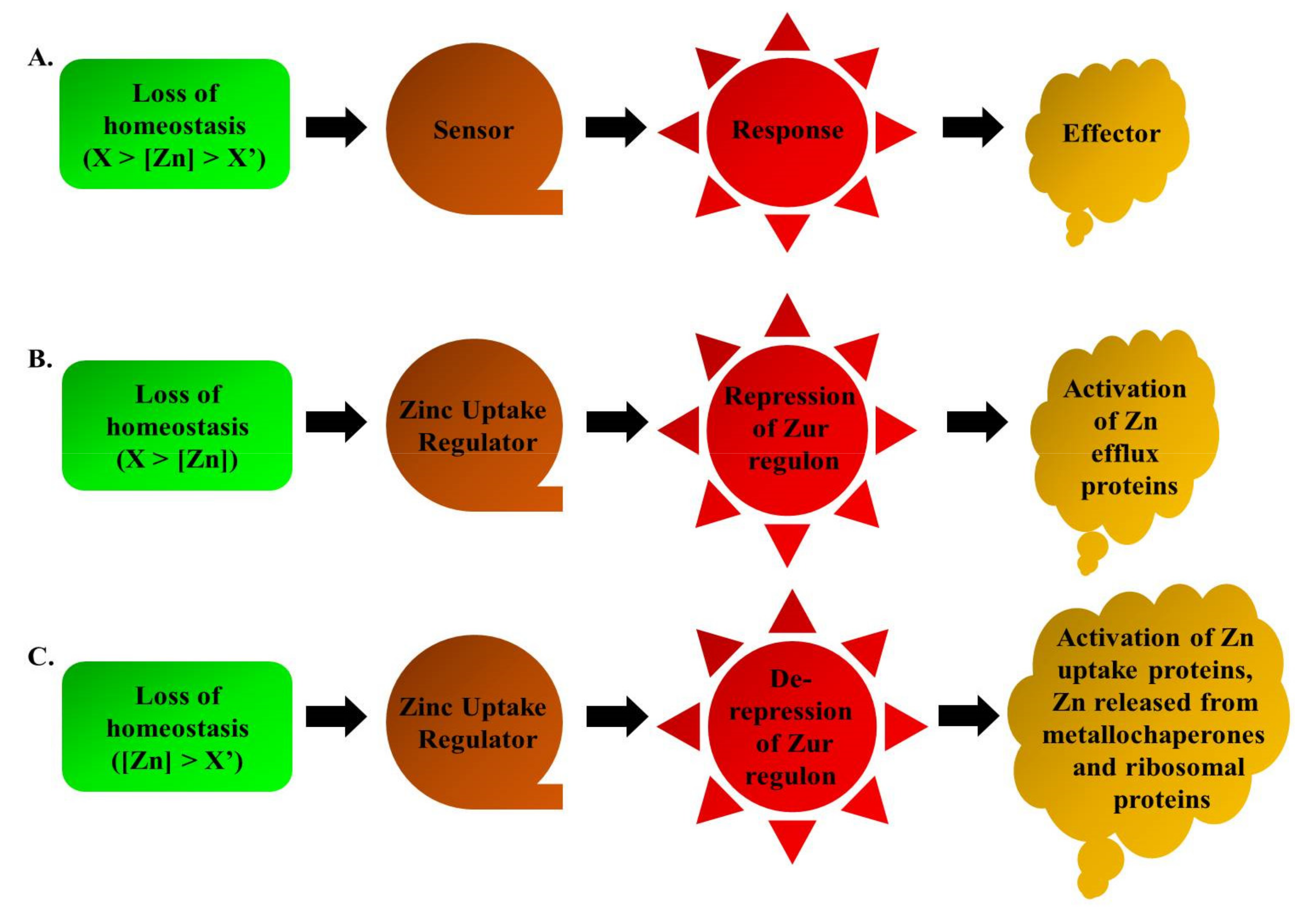
4. Zinc Uptake Regulator
4.1. Background of Zur
4.2. Zur and the Intracellular Zn
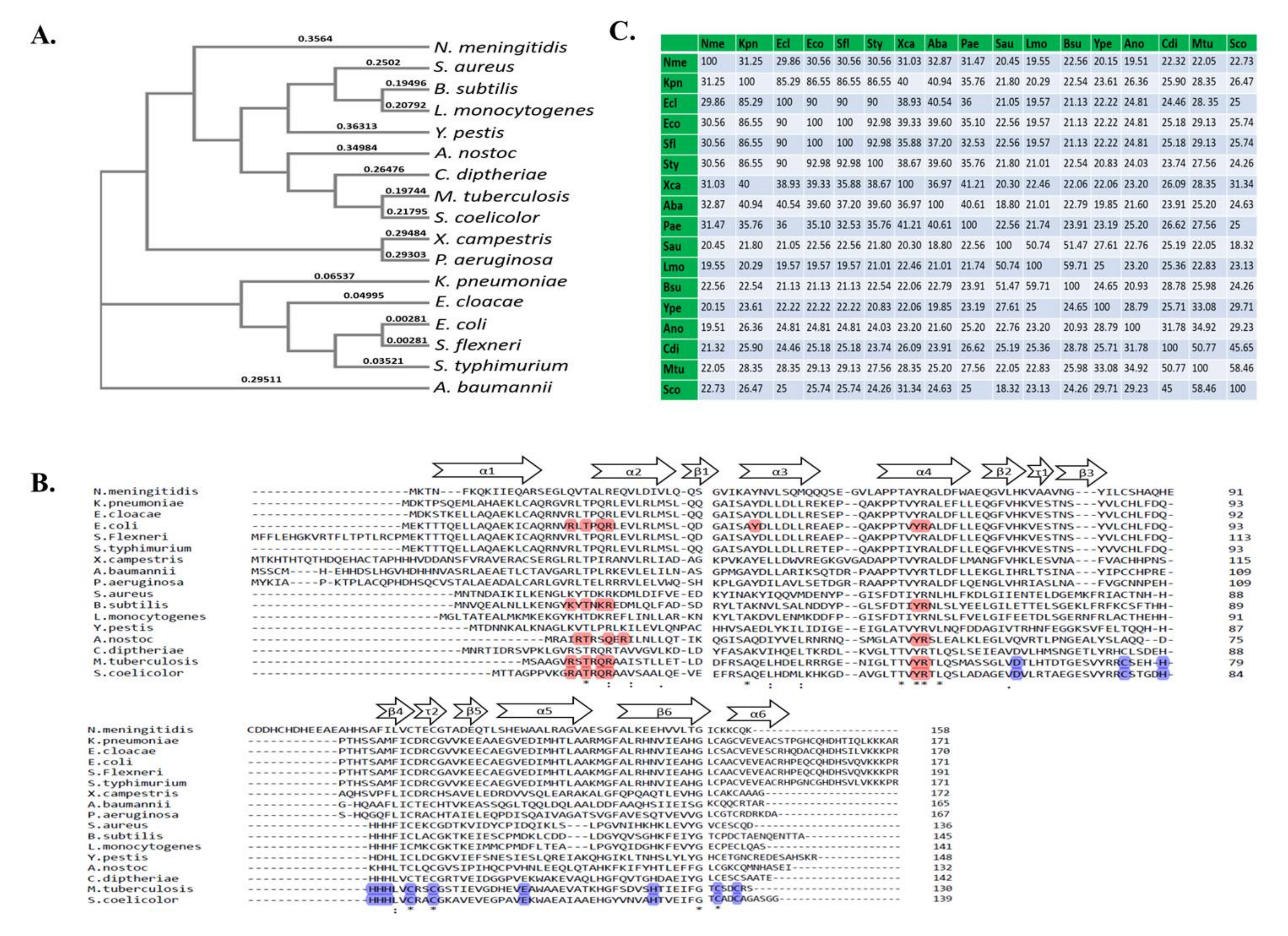
4.3. Zur in Bacterial Species
4.4. Zur-Mediated Zn Homeostasis
5. Structure and the Molecular Mechanisms of Zur
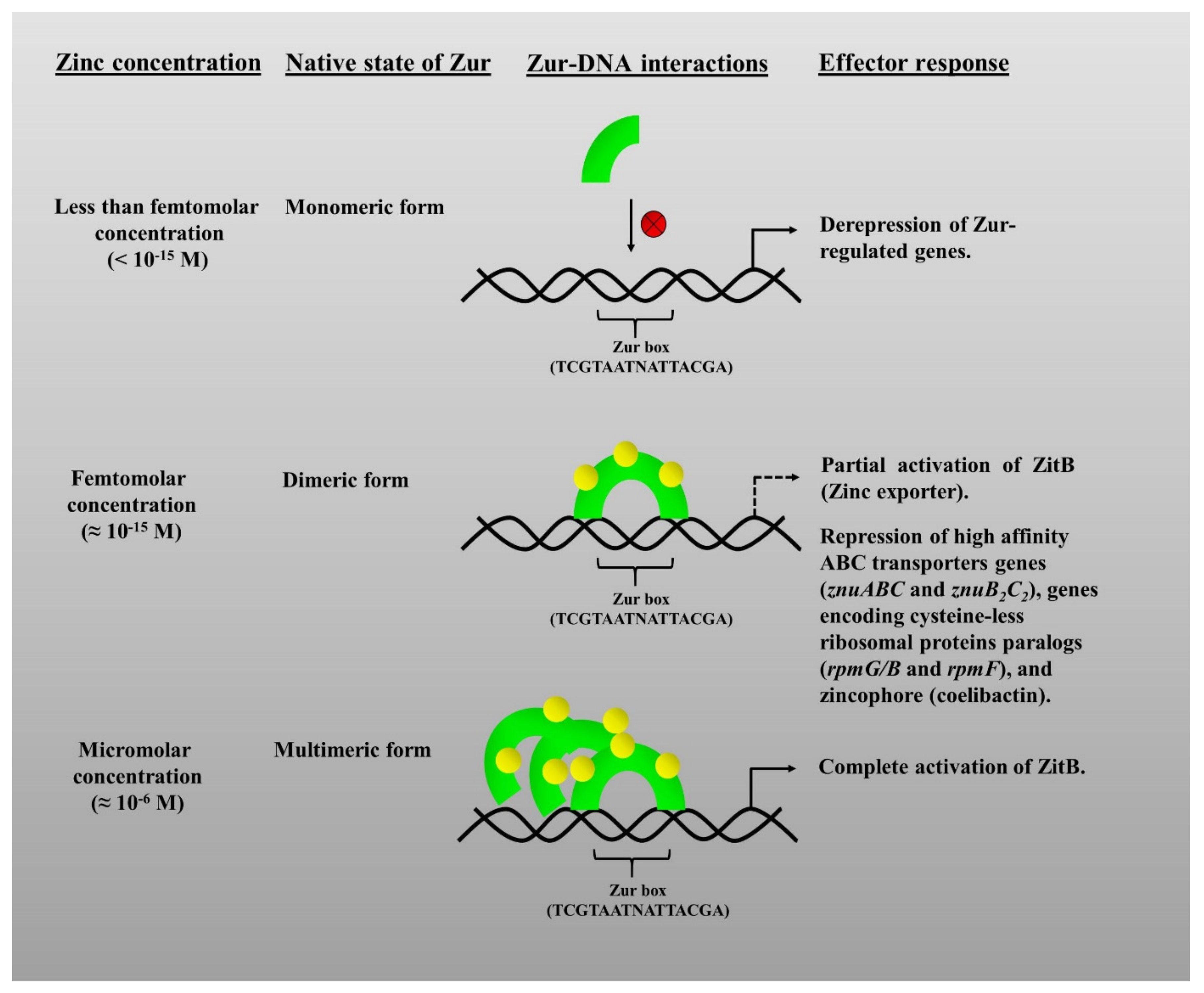
5.1. Structural Aspects of Zur
5.2. The Graded Response of Zur
5.3. Molecular Mechanisms of Zur Proteins
6. Regulon of Zur
6.1. The Non-Zn-Binding Paralogs of Ribosomal Proteins
6.2. High-Affinity Zn Uptake Systems
6.3. Non-Zn-Binding Paralogs of Enzymes
6.4. Metallochaperons
7. Zur and Zur-Regulated Genes, Explored for Their Role in Pathogenicity
8. Scope of Zur Protein as a Putative Antimicrobial Target
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Auld, D.S. Zinc coordination sphere in biochemical zinc sites. BioMetals 2001, 14, 271–313. [Google Scholar] [CrossRef]
- Cerasi, M.; Ammendola, S.; Battistoni, A. Competition for zinc binding in the host-pathogen interaction. Front. Cell. Infect. Microbiol. 2013, 3, 108. [Google Scholar] [CrossRef]
- Outten, C.E.; O’Halloran, T.V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 2001, 292, 2488–2492. [Google Scholar] [CrossRef] [Green Version]
- Klug, A. The discovery of zinc fingers and their applications in gene regulation and genome manipulation. Annu. Rev. Biochem. 2010, 79, 213–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the Three Domains of Life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef]
- Rahman, M.T.; Karim, M.M. Metallothionein: A Potential Link in the Regulation of Zinc in Nutritional Immunity. Biol. Trace Elem. Res. 2018, 182, 1–13. [Google Scholar] [CrossRef]
- Lonergan, Z.R.; Skaar, E.P. Nutrient Zinc at the Host-Pathogen Interface. Trends Biochem. Sci. 2019, 44, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, F.E.; Kazmierczak, K.M.; Lisher, J.P.; Winkler, M.E.; Giedroc, D.P. Interplay between manganese and zinc homeostasis in the human pathogen Streptococcus pneumoniae. Metallomics 2011, 3, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Beard, S.J.; Hughes, M.N.; Poole, R.K. Inhibition of the cytochromebd-terminated NADH oxidase system inEscherichia coliK-12 by divalent metal cations. FEMS Microbiol. Lett. 1995, 131, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Anraku, Y. Succinate- and NADH oxidase systems of Escherichia coli membrane vesicles. Mechanism of selective inhibition of the systems by zinc ions. J. Biochem. 1974, 76, 967–976. [Google Scholar] [PubMed]
- Singh, A.P.; Bragg, P.D. Inhibition of energization of Salmonella typhimurium membrane by zinc ions. FEBS Lett. 1974, 40, 200–202. [Google Scholar] [CrossRef] [Green Version]
- Finney, L.A.; O’Halloran, T.V. Transition metal speciation in the cell: Insights from the chemistry of metal ion receptors. Science 2003, 300, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobine, P.A.; Cruz, L.F.; Navarrete, F.; Duncan, D.; Tygart, M.; De La Fuente, L. Xylella fastidiosa differentially accumulates mineral elements in biofilm and planktonic cells. PLoS ONE 2013, 8, e54936. [Google Scholar] [CrossRef]
- Kallifidas, D.; Pascoe, B.; Owen, G.A.; Strain-Damerell, C.M.; Hong, H.-J.; Paget, M.S.B. The zinc-responsive regulator Zur controls expression of the coelibactin gene cluster in Streptomyces coelicolor. J. Bacteriol. 2009, 192, 608–611. [Google Scholar] [CrossRef] [Green Version]
- Marguerettaz, M.; Dieppois, G.; Que, Y.A.; Ducret, V.; Zuchuat, S.; Perron, K. Sputum containing zinc enhances carbapenem resistance, biofilm formation and virulence of Pseudomonas aeruginosa. Microb. Pathog. 2014, 77, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-H.; Oh, S.-Y.; Kim, S.-J.; Roe, J.-H. The zinc-responsive regulator Zur controls a zinc uptake system and some ribosomal proteins in Streptomyces coelicolor A3(2). J. Bacteriol. 2007, 189, 4070–4077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisner, H.; Porzecanski, B. Inactivation of Penicillin by Zinc Salts. Science 1946, 103, 629–630. [Google Scholar] [CrossRef] [PubMed]
- Novák-Pékli, M.; Mesbah, M.E.-H.; Pethö, G. Equilibrium studies on tetracycline-metal ion systems. J. Pharm. Biomed. Anal. 1996, 14, 1025–1029. [Google Scholar] [CrossRef]
- Mikhaylina, A.; Ksibe, A.Z.; Scanlan, D.J.; Blindauer, C.A. Bacterial zinc uptake regulator proteins and their regulons. Biochem. Soc. Trans. 2018, 46, 983–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lansdown, A.B.G.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Ågren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef]
- Wang, X.X.; Zhang, M.J.; Li, X.B. Advances in the research of zinc deficiency and zinc supplementation treatment in patients with severe burns. Zhonghua Shao Shang Za Zhi 2018, 34, 57–59. [Google Scholar] [CrossRef]
- De Moura, M.S.B.; Soares, N.R.M.; Barros, S.; Érika, D.L.; De Pinho, F.A.; Silva, T.M.C.; Bráz, D.C.; Vieira, E.C.; Lima, M.M.; Parente, J.M.L.; et al. Zinc gluconate supplementation impacts the clinical improvement in patients with ulcerative colitis. BioMetals 2020, 33, 15–27. [Google Scholar] [CrossRef]
- Patel, G.K.; Harding, K.G. Wound problems due to zinc deficiency. Int. Wound J. 2004, 1, 150–151. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Genet. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Kehl-Fie, T.E.; Skaar, E.P. Nutritional immunity beyond iron: A role for manganese and zinc. Curr. Opin. Chem. Biol. 2010, 14, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, A.; Heesemann, L.; Wagener, J.; Marcos, V.; Hartl, D.; Loeffler, J.; Heesemann, J.; Ebel, F. NETs formed by human neutrophils inhibit growth of the pathogenic mold Aspergillus fumigatus. Microbes Infect. 2010, 12, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Moayeri, M.; Leppla, S.H.; Vrentas, C.; Pomerantsev, A.P.; Liu, S. Anthrax Pathogenesis. Annu. Rev. Microbiol. 2015, 69, 185–208. [Google Scholar] [CrossRef]
- Moroz, O.V.; Antson, A.; Grist, S.J.; Maitland, N.; Dodson, G.G.; Wilson, K.S.; Lukanidin, E.; Bronstein, I.B. Structure of the human S100A12-copper complex: Implications for host-parasite defence. Acta Crystallogr. Sect. D Biol. Crystallogr. 2003, 59 Pt 5, 859–867. [Google Scholar] [CrossRef]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef]
- Gläser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schröder, J.-M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef]
- Moroz, O.V.; Burkitt, W.; Wittkowski, H.; He, W.; Ianoul, A.; Novitskaya, V.; Xie, J.; Polyakova, O.; Lednev, I.K.; Shekhtman, A.; et al. Both Ca2+ and Zn2+ are essential for S100A12 protein oligomerization and function. BMC Biochem. 2009, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaia, A.A.; Sappington, K.J.; Nisapakultorn, K.; Chazin, W.J.; Dietrich, E.A.; Ross, K.F.; Herzberg, M.C. Subversion of antimicrobial calprotectin (S100A8/S100A9 complex) in the cytoplasm of TR146 epithelial cells after invasion by Listeria monocytogenes. Mucosal. Immunol. 2009, 2, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grim, K.P.; Francisco, B.S.; Radin, J.N.; Brazel, E.B.; Kelliher, J.L.; Solórzano, P.K.P.; Kim, P.C.; McDevitt, C.A.; Kehl-Fie, T.E. The metallophore staphylopine enables Staphylococcus aureus to compete with the host for zinc and overcome nutritional immunity. MBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Z.; Jellbauer, S.; Poe, A.J.; Ton, V.; Pesciaroli, M.; Kehl-Fie, T.E.; Restrepo, N.A.; Hosking, M.P.; Edwards, R.A.; Battistoni, A.; et al. Zinc sequestration by the neutrophil protein calprotectin enhances Salmonella growth in the inflamed gut. Cell Host Microbe 2012, 11, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, M.R.; Ducret, V.; Leoni, S.; Perron, K. Pseudomonas aeruginosa zinc homeostasis: Key issues for an opportunistic pathogen. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 722–733. [Google Scholar] [CrossRef]
- Ammendola, S.; Pasquali, P.; Pistoia, C.; Petrucci, P.; Petrarca, P.; Rotilio, G.; Battistoni, A. High-affinity Zn2+ uptake system ZnuABC is required for bacterial zinc homeostasis in intracellular environments and contributes to the virulence of Salmonella enterica. Infect. Immun. 2007, 75, 5867–5876. [Google Scholar] [CrossRef] [Green Version]
- Campoy, S.; Jara, M.; Busquets, N.; De Rozas, A.M.P.; Badiola, I.; Barbé, J. Role of the high-affinity zinc uptake znuABC system in Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2002, 70, 4721–4725. [Google Scholar] [CrossRef] [Green Version]
- Garrido, M.E.; Bosch, M.; Medina, R.; Llagostera, M.; De Rozas, A.M.P.; Badiola, I.; Barbé, J. The high-affinity zinc-uptake system znuACB is under control of the iron-uptake regulator (fur) gene in the animal pathogen Pasteurella multocida. FEMS Microbiol. Lett. 2003, 221, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Watanabe, K.; Shirahata, T.; Watarai, M. Zinc uptake system (znuA locus) of Brucella abortus is essential for intracellular survival and virulence in mice. J. Veter Med. Sci. 2004, 66, 1059–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobrov, A.G.; Kirillina, O.; Fetherston, J.D.; Miller, M.C.; Burlison, J.A.; Perry, R. The Yersinia pestis siderophore, yersiniabactin, and the ZnuABC system both contribute to zinc acquisition and the development of lethal septicaemic plague in mice. Mol. Microbiol. 2014, 93, 759–775. [Google Scholar] [CrossRef] [Green Version]
- Graham, A.I.; Hunt, S.; Stokes, S.L.; Bramall, N.; Bunch, J.; Cox, A.G.; McLeod, C.W.; Poole, R.K.; Oka, M.; Sumita, N.; et al. Severe zinc depletion of Escherichia coli roles for high affinity zinc binding by ZinT, zinc transport and zinc-independent proteins. J. Biol. Chem. 2009, 284, 18377–18389. [Google Scholar] [CrossRef] [Green Version]
- Ilari, A.; Alaleona, F.; Tria, G.; Petrarca, P.; Battistoni, A.; Zamparelli, C.; Verzili, D.; Falconi, M.; Chiancone, E. The Salmonella enterica ZinT structure, zinc affinity and interaction with the high-affinity uptake protein ZnuA provide insight into the management of periplasmic zinc. Biochim. Biophys. Acta 2014, 1840, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Panina, E.M.; Mironov, A.A.; Gelfand, M.S. Comparative genomics of bacterial zinc regulons: Enhanced ion transport, pathogenesis, and rearrangement of ribosomal proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 9912–9917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrarca, P.; Ammendola, S.; Pasquali, P.; Battistoni, A. The Zur-regulated ZinT protein is an auxiliary component of the high-affinity ZnuABC zinc transporter that facilitates metal recruitment during severe zinc shortage. J. Bacteriol. 2010, 192, 1553–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerasi, M.; Liu, J.Z.; Ammendola, S.; Poe, A.J.; Petrarca, P.; Pesciaroli, M.; Pasquali, P.; Raffatellu, M.; Battistoni, A. The ZupT transporter plays an important role in zinc homeostasis and contributes to Salmonella enterica virulence. Metallomics 2014, 6, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Grass, G.; Wong, M.D.; Rosen, B.P.; Smith, R.L.; Rensing, C. ZupT is a Zn (II) uptake system in Escherichia coli. J. Bacteriol. 2002, 184, 864–866. [Google Scholar] [CrossRef] [Green Version]
- Sabri, M.; Houle, S.; Dozois, C.M. Roles of the extraintestinal pathogenic Escherichia coli ZnuACB and ZupT zinc transporters during urinary tract infection. Infection and immunity 2009, 77, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Rosadini, C.V.; Gawronski, J.D.; Raimunda, D.; Argüello, J.M.; Akerley, B.J. A novel zinc binding system, ZevAB, is critical for survival of nontypeable Haemophilus influenzae in a murine lung infection model. Infection and immunity 2011, 79, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Corbett, D.; Wang, J.; Schuler, S.; Lopez-Castejon, G.; Glenn, S.; Brough, D.; Andrew, P.W.; Cavet, J.S.; Roberts, I.S. Two zinc uptake systems contribute to the full virulence of Listeria monocytogenes during growth in vitro and in vivo. Infection and immunity 2012, 80, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Moody, S.C.; Hider, R.C.; Lei, L.; Kelly, S.L.; Waterman, M.R.; Lamb, D.C. Structural analysis of cytochrome P450 105N1 involved in the biosynthesis of the zincophore, coelibactin. Int. J. Mol. Sci. 2012, 13, 8500–8513. [Google Scholar] [CrossRef] [Green Version]
- Si, M.; Wang, Y.; Zhang, B.; Zhao, C.; Kang, Y.; Bai, H.; Wei, D.; Zhu, L.; Zhang, L.; Dong, T.G.; et al. The Type VI Secretion System Engages a Redox-Regulated Dual-Functional Heme Transporter for Zinc Acquisition. Cell Rep. 2017, 20, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Asmis, K.R.; Pivonka, N.L.; Santambrogio, G.; Brümmer, M.; Kaposta, C.; Neumark, D.M.; Wöste, L. Passage of heme-iron across the envelope of Staphylococcus aureus. Science 2003, 299, 906–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stork, M.; Grijpstra, J.; Bos, M.P.; Torres, C.M.; Devos, N.; Poolman, J.T.; Chazin, W.J.; Tommassen, J. Zinc piracy as a mechanism of Neisseria meningitidis for evasion of nutritional immunity. PLOS Pathog. 2013, 9, e1003733. [Google Scholar] [CrossRef]
- Capdevila, D.A.; Wang, J.; Giedroc, D.P. Bacterial Strategies to Maintain Zinc Metallostasis at the Host-Pathogen Interface. J. Biol. Chem. 2016, 291, 20858–20868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wątły, J.; Potocki, S.; Rowińska-Żyrek, M. Zinc Homeostasis at the Bacteria/Host Interface-From Coordination Chemistry to Nutritional Immunity. Chemistry 2016, 22, 15992–16010. [Google Scholar] [CrossRef]
- Assunção, A.G.L.; Herrero, E.; Lin, Y.-F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; Van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [Green Version]
- Brugnera, E.; Georgiev, O.; Radtke, F.; Heuchel, R.; Baker, E.; Sutherland, G.R.; Schaffner, W. Cloning, chromosomal mapping and characterization of the human metal-regulatory transcription factor MTF-1. Nucleic Acids Res. 1994, 22, 3167–3173. [Google Scholar] [CrossRef] [Green Version]
- Corkins, M.E.; May, M.; Ehrensberger, K.M.; Hu, Y.-M.; Liu, Y.-H.; Bloor, S.D.; Jenkins, B.; Runge, K.W.; Bird, A.J. Zinc finger protein Loz1 is required for zinc-responsive regulation of gene expression in fission yeast. Proc. Natl. Acad. Sci. USA 2013, 110, 15371–15376. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Bird, A.J. Zinc’ing sensibly: Controlling zinc homeostasis at the transcriptional level. Metallomics 2014, 6, 1198–1215. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Lee, K.-L.; Shin, J.-H.; Cho, Y.-B.; Cha, S.-S.; Roe, J.-H. Zinc-dependent regulation of zinc import and export genes by Zur. Nat. Commun. 2017, 8, 15812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fillat, M.F. The FUR (ferric uptake regulator) superfamily: Diversity and versatility of key transcriptional regulators. Arch. Biochem. Biophys. 2014, 546, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Escolar, L.; Pérez-Martín, J.; De Lorenzo, V. Opening the iron box: Transcriptional metalloregulation by the Fur protein. J. Bacteriol. 1999, 181, 6223–6229. [Google Scholar] [CrossRef] [Green Version]
- Hantke, K. Members of the Fur protein family regulate iron and zinc transport in E. coli and characteristics of the Fur-regulated fhuF protein. J. Mol. Microbiol. Biotechnol. 2002, 4, 217–222. [Google Scholar]
- Lee, J.-W.; Helmann, J.D. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature 2006, 440, 363–367. [Google Scholar] [CrossRef]
- Verneuil, N.; Rincé, A.; Sanguinetti, M.; Posteraro, B.; Fadda, G.; Auffray, Y.; Hartke, A.; Giard, J.-C. Contribution of a PerR-like regulator to the oxidative-stress response and virulence of Enterococcus faecalis. Microbiology 2005, 151 Pt 12, 3997–4004. [Google Scholar] [CrossRef] [Green Version]
- Ahn, B.E.; Cha, J.; Lee, E.J.; Han, A.R.; Thompson, C.J.; Roe, J.H. Nur, a nickel-responsive regulator of the Fur family, regulates superoxide dismutases and nickel transport in Streptomyces coelicolor. Mol. Microbiol. 2006, 59, 1848–1858. [Google Scholar] [CrossRef]
- Díaz-Mireles, E.; Wexler, M.; Sawers, G.; Bellini, D.; Todd, J.D.; Johnston, A.W.B. The Fur-like protein Mur of Rhizobium leguminosarum is a Mn2+-responsive transcriptional regulator. Microbiology 2004, 150 Pt 5, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Platero, R.; De Lorenzo, V.; Garat, B.; Fabiano, E. Sinorhizobium meliloti fur-like (Mur) protein binds a fur box-like sequence present in the mntA promoter in a manganese-responsive manner. Appl. Environ. Microbiol. 2007, 73, 4832–4838. [Google Scholar] [CrossRef] [Green Version]
- Patzer, S.I.; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol. 1998, 28, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Gaballa, A.; Helmann, J.D. Identification of a zinc-specific metalloregulatory protein, Zur, controlling zinc transport operons in Bacillus subtilis. J. Bacteriol. 1998, 180, 5815–5821. [Google Scholar] [CrossRef] [Green Version]
- Prestel, E.; Noirot, P.; Auger, S. Genome-wide identification of Bacillus subtilis Zur-binding sites associated with a Zur box expands its known regulatory network. BMC Microbiol. 2015, 15, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.-H.; Helmann, J.D. Molecular logic of the Zur-regulated zinc deprivation response in Bacillus subtilis. Nat. Commun. 2016, 7, 12612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalet, K.; Gouin, E.; Cenatiempo, Y.; Cossart, P.; Héchard, Y. Characterisation of a new operon encoding a Zur-like protein and an associated ABC zinc permease in Listeria monocytogenes. FEMS Microbiol. Lett. 1999, 174, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.-J.; Li, X.-J.; He, Y.-Q.; Feng, J.-X.; Chen, B.; Tang, J.-L. The zinc uptake regulator Zur is essential for the full virulence of Xanthomonas campestris pv. campestris. Mol. Plant-Microbe Interact. 2005, 18, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Li, M.; Zhang, H.; Zheng, B.; Han, H.; Wang, C.; Yan, J.; Tang, J.; Gao, G.F. Functional definition and global regulation of Zur, a zinc uptake regulator in a Streptococcus suis serotype 2 strain causing streptococcal toxic shock syndrome. J. Bacteriol. 2008, 190, 7567–7578. [Google Scholar] [CrossRef] [Green Version]
- Latorre, M.; Low, M.; Gárate, E.; Reyes-Jara, A.; Murray, B.E.; Cambiazo, V.; González, M. Interplay between copper and zinc homeostasis through the transcriptional regulator Zur in Enterococcus faecalis. Metallomics 2015, 7, 1137–1145. [Google Scholar] [CrossRef]
- Maciag, A.; Dainese, E.; Rodriguez, G.M.; Milano, A.; Provvedi, R.; Pasca, M.R.; Smith, I.; Palù, G.; Riccardi, G.; Manganelli, R. Global analysis of the Mycobacterium tuberculosis Zur (FurB) regulon. J. Bacteriol. 2007, 189, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hurst, T.K.; Thompson, R.B.; Fierke, C.A. Genetically encoded ratiometric biosensors to measure intracellular exchangeable zinc in Escherichia coli. J. Biomed. Opt. 2011, 16, 087011. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Jung, H.J.; An, Y.J.; Cho, Y.-B.; Cha, S.-S.; Roe, J.-H. Graded expression of zinc-responsive genes through two regulatory zinc-binding sites in Zur. Proc. Natl. Acad. Sci. USA 2011, 108, 5045–5050. [Google Scholar] [CrossRef] [Green Version]
- Gradassi, M.; Pesciaroli, M.; Martinelli, N.; Ruggeri, J.; Petrucci, P.; Hassan, W.H.; Raffatellu, M.; Scaglione, F.E.; Ammendola, S.; Battistoni, A.; et al. Attenuated Salmonella enterica serovar Typhimurium lacking the ZnuABC transporter: An efficacious orally-administered mucosal vaccine against salmonellosis in pigs. Vaccine 2013, 31, 3695–3701. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Gradassi, M.; Martinelli, N.; Ruggeri, J.; Pistoia, C.; Raffatellu, M.; Magistrali, C.; Battistoni, A.; Pasquali, P.; Alborali, G. Salmonella Typhimurium lacking the Znuabc transporter is attenuated and immunogenic in pigs. Vaccine 2013, 31, 2868–2873. [Google Scholar] [CrossRef]
- Lucarelli, D.; Russo, S.; Garman, E.; Milano, A.; Meyer-Klaucke, W.; Pohl, E. Crystal structure and function of the zinc uptake regulator FurB from Mycobacterium tuberculosis. J. Biol. Chem. 2007, 282, 9914–9922. [Google Scholar] [CrossRef] [Green Version]
- Gilston, B.A.; Wang, S.; Marcus, M.D.; Canalizo-Hernández, M.A.; Swindell, E.P.; Xue, Y.; Mondragón, A.; O’Halloran, T.V. Structural and mechanistic basis of zinc regulation across the E. coli Zur regulon. PLoS Biol. 2014, 12, e1001987. [Google Scholar] [CrossRef]
- D’Autréaux, B.; Pecqueur, L.; De Peredo, A.G.; Diederix, R.E.M.; Caux-Thang, C.; Tabet, L.; Bersch, B.; Forest, E.; Michaud-Soret, I. Reversible redox- and zinc-dependent dimerization of the Escherichia coli fur protein. Biochemistry 2007, 46, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Helmann, J.D. Biochemical characterization of the structural Zn2+ site in the Bacillus subtilis peroxide sensor PerR. J. Biol. Chem. 2006, 281, 23567–23578. [Google Scholar] [CrossRef] [Green Version]
- Pecqueur, L.; D’Autréaux, B.; Dupuy, J.; Nicolet, Y.; Jacquamet, L.; Brutscher, B.; Michaud-Soret, I.; Bersch, B. Structural changes of Escherichia coli ferric uptake regulator during metal-dependent dimerization and activation explored by NMR and X-ray crystallography. J. Biol. Chem. 2006, 281, 21286–21295. [Google Scholar] [CrossRef] [Green Version]
- Vitale, S.; Fauquant, C.; Lascoux, D.; Schauer, K.; Saint-Pierre, C.; Michaud-Soret, I. A ZnS4 structural zinc site in the Helicobacter pylori ferric uptake regulator. Biochemistry 2009, 48, 5582–5591. [Google Scholar] [CrossRef]
- Osman, D.; Cavet, J.S. Metal sensing in Salmonella: Implications for pathogenesis. Adv. Microb. Physiol. 2011, 58, 175–232. [Google Scholar] [CrossRef]
- Kandari, D.; Joshi, H.; Tanwar, N.; Munde, M.; Bhatnagar, R. Delineation of the Residues of Bacillus anthracis Zinc Uptake Regulator Protein Directly Involved in Its Interaction with Cognate DNA. Biol. Trace Element Res. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kandari, D.; Gopalani, M.; Gupta, M.; Joshi, H.; Bhatnagar, S.; Bhatnagar, R. Identification, Functional Characterization, and Regulon Prediction of the Zinc Uptake Regulator (zur) of Bacillus anthracis—An insight into the Zinc Homeostasis of the Pathogen. Front. Microbiol. 2019, 9, 3314. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.; Sengupta, K.; Wendel, B.M.; Helmann, J.D.; Chen, P. Biphasic unbinding of a metalloregulator from DNA for transcription (de) repression in Live Bacteria. Nucleic Acids Res. 2020, 48, 2199–2208. [Google Scholar] [CrossRef]
- Huang, D.-L.; Tang, D.-J.; Liao, Q.; Li, H.-C.; Chen, Q.; He, Y.-Q.; Feng, J.-X.; Jiang, B.-L.; Lu, G.-T.; Chen, B.; et al. The Zur of Xanthomonas campestris functions as a repressor and an activator of putative zinc homeostasis genes via recognizing two distinct sequences within its target promoters. Nucleic Acids Res. 2008, 36, 4295–4309. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Chua, T.T.; Chew, C.Y.; Bryant, D.A. Fur-type transcriptional repressors and metal homeostasis in the cyanobacterium Synechococcus sp. PCC 7002. Front. Microbiol. 2015, 6, 1217. [Google Scholar] [CrossRef] [Green Version]
- Neupane, D.P.; Jacquez, B.; Sundararajan, A.; Ramaraj, T.; Schilkey, F.D.; Yukl, E.T. Zinc-dependent transcriptional regulation in Paracoccus denitrificans. Front. Microbiol. 2017, 8, 569. [Google Scholar] [CrossRef] [Green Version]
- Bhubhanil, S.; Sittipo, P.; Chaoprasid, P.; Nookabkaew, S.; Sukchawalit, R.; Mongkolsuk, S. Control of zinc homeostasis in Agrobacterium tumefaciens via zur and the zinc uptake genes znuABC and zinT. Microbiology 2014, 160, 2452–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaoprasid, P.; Dokpikul, T.; Johnrod, J.; Sirirakphaisarn, S.; Nookabkaew, S.; Sukchawalit, R.; Mongkolsuk, S. Agrobacterium tumefaciens Zur regulates the high-affinity zinc uptake system TroCBA and the putative metal chaperone YciC, along with ZinT and ZnuABC, for survival under zinc-limiting conditions. Appl. Environ. Microbiol. 2016, 82, 3503–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, B.L.; Rathi, S.; Chazin, W.J.; Skaar, E.P. Acinetobacter baumannii response to host-mediated zinc limitation requires the transcriptional regulator Zur. J. Bacteriol. 2014, 196, 2616–2626. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.K.; Hassan, K.A.; Penesyan, A.; Loper, J.E.; Paulsen, I.T. The effect of zinc limitation on the transcriptome of Pseudomonas protegens Pf-5. Environ. Microbiol. 2013, 15, 702–715. [Google Scholar] [CrossRef] [Green Version]
- Ellison, M.L.; Farrow, J.M.; Parrish, W.; Danell, A.S.; Pesci, E.C. The transcriptional regulator Np20 is the zinc uptake regulator in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e75389. [Google Scholar] [CrossRef]
- Lhospice, S.; Gomez, N.O.; Ouerdane, L.; Brutesco, C.; Ghssein, G.; Hajjar, C.; Liratni, A.; Wang, S.; Richaud, P.; Bleves, S.; et al. Pseudomonas aeruginosa zinc uptake in chelating environment is primarily mediated by the metallophore pseudopaline. Sci. Rep. 2017, 7, 17132. [Google Scholar] [CrossRef] [Green Version]
- Pederick, V.G.; Eijkelkamp, B.A.; Begg, S.L.; Ween, M.P.; McAllister, L.J.; Paton, J.C.; McDevitt, C.A. ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 13139. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.A.; Foster, S.J. zur: A Zn2+-responsive regulatory element of Staphylococcus aureus. Microbiology 2001, 147 Pt 5, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Dowd, G.C.; Casey, P.G.; Begley, M.; Hill, C.; Gahan, C.G. Investigation of the role of ZurR in the physiology and pathogenesis of Listeria monocytogenes. FEMS Microbiol. Lett. 2012, 327, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Desrosiers, D.C.; Bearden, S.W.; Mier, I.; Abney, J.; Paulley, J.T.; Fetherston, J.D.; Salazar, J.C.; Radolf, J.D.; Perry, R.D. Znu is the predominant zinc importer in Yersinia pestis during in vitro growth but is not essential for virulence. Infect. Immun. 2010, 78, 5163–5177. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Klesney-Tait, J.; Lumbley, S.R.; Ward, C.K.; Latimer, J.L.; Ison, C.A.; Hansen, E.J. Identification of the znuA-encoded periplasmic zinc transport protein of Haemophilus ducreyi. Infect. Immun. 1999, 67, 5060–5068. [Google Scholar] [CrossRef] [Green Version]
- Hensley, M.P.; Tierney, D.L.; Crowder, M.W. Zn(II) binding to Escherichia coli 70S ribosomes. Biochemistry 2011, 50, 9937–9939. [Google Scholar] [CrossRef] [Green Version]
- Gaballa, A.; Wang, T.; Ye, R.W.; Helmann, J.D. Functional Analysis of the Bacillus subtilis Zur Regulon. J. Bacteriol. 2002, 184, 6508–6514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastropasqua, M.C.; D’Orazio, M.; Cerasi, M.; Pacello, F.; Gismondi, A.; Canini, A.; Canuti, L.; Consalvo, A.; Ciavardelli, D.; Chirullo, B.; et al. Growth of Pseudomonas aeruginosa in zinc poor environments is promoted by a nicotianamine-related metallophore. Mol. Microbiol. 2017, 106, 543–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobrov, A.G.; Kirillina, O.; Fosso, M.Y.; Fetherston, J.D.; Miller, M.C.; VanCleave, T.T.; Burlison, J.A.; Arnold, W.K.; Lawrenz, M.B.; Garneau-Tsodikova, S.; et al. Zinc transporters YbtX and ZnuABC are required for the virulence of Yersinia pestis in bubonic and pneumonic plague in mice. Metallomics 2017, 9, 757–772. [Google Scholar] [CrossRef]
- Fojcik, C.; Arnoux, P.; Ouerdane, L.; Aigle, M.; Alfonsi, L.; Borezée-Durant, E. Independent and cooperative regulation of staphylopine biosynthesis and trafficking by Fur and Zur. Mol. Microbiol. 2018, 108, 159–177. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, M.-C.; Hubert, K.; Joseph, B.; Claus, H.; Schoen, C.; Vogel, U. The zinc-responsive regulon of Neisseria meningitidis comprises 17 genes under control of a Zur element. J. Bacteriol. 2012, 194, 6594–6603. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, B.; Bonnett, S.A.; Shah, K.; Gabriel, S.; Reddy, R.; Schimmel, P.; Rodionov, D.A.; De Crécy-Lagard, V.; Helmann, J.D.; Iwata-Reuyl, D.; et al. Zinc-Independent Folate Biosynthesis: Genetic, Biochemical, and Structural Investigations Reveal New Metal Dependence for GTP Cyclohydrolase IB. J. Bacteriol. 2009, 191, 6936–6949. [Google Scholar] [CrossRef] [Green Version]
- Blaby-Haas, C.E.; Furman, R.; Rodionov, D.A.; Artsimovitch, I.; De Crécy-Lagard, V. Role of a Zn-independent DksA in Zn homeostasis and stringent response. Mol. Microbiol. 2010, 79, 700–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, C.E.; Rodionov, D.A.; Kropat, J.; Malasarn, D.; Merchant, S.S.; De Crécy-Lagard, V. A subset of the diverse COG0523 family of putative metal chaperones is linked to zinc homeostasis in all kingdoms of life. BMC Genom. 2009, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Blaby-Haas, C.E.; Flood, J.A.; de Crécy-Lagard, V.; Zamble, D.B. YeiR: A metal-binding GTPase from Escherichia coli involved in metal homeostasis. Metallomics 2012, 4, 488–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khil, P.P.; Obmolova, G.; Teplyakov, A.; Howard, A.J.; Gilliland, G.L.; Camerini-Otero, R.D. Crystal structure of the Escherichia coli YjiA protein suggests a GTP-dependent regulatory function. Proteins 2004, 54, 371–374. [Google Scholar] [CrossRef]
- Nairn, B.L.; Lonergan, Z.R.; Wang, J.; Braymer, J.J.; Zhang, Y.; Calcutt, M.W.; Lisher, J.P.; Gilston, B.A.; Chazin, W.J.; De Crécy-Lagard, V.; et al. The Response of Acinetobacter baumannii to Zinc Starvation. Cell Host Microbe 2016, 19, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydor, A.M.; Jost, M.; Ryan, K.S.; Turo, K.E.; Douglas, C.D.; Drennan, C.L.; Zamble, D.B. Metal binding properties of Escherichia coli YjiA, a member of the metal homeostasis-associated COG0523 family of GTPases. Biochemistry 2013, 52, 1788–1801. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.; Jochmann, N.; Rodionov, D.A.; Tauch, A. The Zur regulon of Corynebacterium glutamicum ATCC 13032. BMC Genom. 2010, 11, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Qiu, Y.; Gao, H.; Guo, Z.; Han, Y.; Song, Y.; Du, Z.; Wang, X.; Zhou, D.; Yang, R. Characterization of Zur-dependent genes and direct Zur targets in Yersinia pestis. BMC Microbiol. 2009, 9, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Becker, T.; Walters, N.; Pascual, D.W. Deletion of znuA virulence factor attenuates Brucella abortus and confers protection against wild-type challenge. Infect. Immun. 2006, 74, 3874–3879. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.M.; Kakuda, T.; DiRita, V.J. A Campylobacter jejuni znuA orthologue is essential for growth in low-zinc environments and chick colonization. Journal of bacteriology 2009, 191, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Gabbianelli, R.; Scotti, R.; Ammendola, S.; Petrarca, P.; Nicolini, L.; Battistoni, A. Role of ZnuABC and ZinT in Escherichia coli O157: H7 zinc acquisition and interaction with epithelial cells. BMC microbiology 2011, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.F.; Brauer, A.L.; Kirkham, C.; Johnson, A.; Koszelak-Rosenblum, M.; Malkowski, M.G. Role of the zinc uptake ABC transporter of Moraxella catarrhalis in persistence in the respiratory tract. Infection and immunity 2013, 81, 3406–3413. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, I.; Stevenson, R.M. The ZnuABC operon is important for Yersinia ruckeri infections of rainbow trout, Oncorhynchus mykiss (Walbaum). Journal of fish diseases 2010, 33, 331–340. [Google Scholar] [CrossRef]
- Pasquali, P.; Ammendola, S.; Pistoia, C.; Petrucci, P.; Tarantino, M.; Valente, C.; Marenzoni, M.L.; Rotilio, G.; Battistoni, A. Attenuated Salmonella enterica serovar Typhimurium lacking the ZnuABC transporter confers immune-based protection against challenge infections in mice. Vaccine 2008, 26, 3421–3426. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Aloisio, F.; Ammendola, S.; Pistoia, C.; Petrucci, P.; Tarantino, M.; Francia, M.; Battistoni, A.; Pasquali, P. An attenuated Salmonella enterica serovar Typhimurium strain lacking the ZnuABC transporter induces protection in a mouse intestinal model of Salmonella infection. Vaccine 2011, 29, 1783–1790. [Google Scholar] [CrossRef]
- Henderson, B.; Martin, A. Bacterial moonlighting proteins and bacterial virulence. In Between Pathogenicity and Commensalism; Springer: Berlin/Heidelberg, Germany, 2011; pp. 155–213. [Google Scholar] [CrossRef]
- Iyoda, S.; Kamidoi, T.; Hirose, K.; Kutsukake, K.; Watanabe, H. A flagellar gene fliZ regulates the expression of invasion genes and virulence phenotype in Salmonella enterica serovar Typhimurium. Microb. Pathog. 2001, 30, 81–90. [Google Scholar] [CrossRef]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Orazio, M.; Mastropasqua, M.C.; Cerasi, M.; Pacello, F.; Consalvo, A.; Chirullo, B.; Mortensen, B.; Skaar, E.P.; Ciavardelli, D.; Pasquali, P.; et al. The capability of Pseudomonas aeruginosa to recruit zinc under conditions of limited metal availability is affected by inactivation of the ZnuABC transporter. Metallomics 2015, 7, 1023–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camilli, A.; Mekalanos, J.J. Use of recombinase gene fusions to identify Vibrio cholerae genes induced during infection. Mol. Microbiol. 1995, 18, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, E.; Wang, S.; Sanet, M.; Fernández-Vázquez, J.; Jové, D.; Glaría, E.; Valledor, A.F.; O’Halloran, T.V.; Balsalobre, C. A new role for Zinc limitation in bacterial pathogenicity: Modulation of α-hemolysin from uropathogenic Escherichia coli. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistoni, A.; Ammendola, S.; Chiancone, E.; Ilari, A. A novel antimicrobial approach based on the inhibition of zinc uptake in Salmonella enterica. Futur. Med. Chem. 2017, 9, 899–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, D.J.; Sokol, P.A.; Howell, M.L.; Ma, J.F.; Schweizer, H.T.; Ochsner, U.; Vasil, M.L. Ferric uptake regulator (Fur) mutants of Pseudomonas aeruginosa demonstrate defective siderophore-mediated iron uptake, altered aerobic growth, and decreased superoxide dismutase and catalase activities. J. Bacteriol. 1996, 178, 3996–4003. [Google Scholar] [CrossRef] [Green Version]
- Horsburgh, M.J.; Ingham, E.; Foster, S.J. In Staphylococcus aureus, fur is an interactive regulator with PerR, contributes to virulence, and is necessary for oxidative stress resistance through positive regulation of catalase and iron homeostasis. J. Bacteriol. 2001, 183, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Mey, A.R.; Wyckoff, E.E.; Kanukurthy, V.; Fisher, C.R.; Payne, S.M. Iron and fur regulation in Vibrio cholerae and the role of fur in virulence. Infect. Immun. 2005, 73, 8167–8178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cissé, C.; Mathieu, S.V.; Abeih, M.B.O.; Flanagan, L.; Vitale, S.; Catty, P.; Boturyn, D.; Michaud-Soret, I.; Crouzy, S. Inhibition of the ferric uptake regulator by peptides derived from anti-FUR peptide aptamers: Coupled theoretical and experimental approaches. ACS Chem. Biol. 2014, 9, 2779–2786. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandari, D.; Joshi, H.; Bhatnagar, R. Zur: Zinc-Sensing Transcriptional Regulator in a Diverse Set of Bacterial Species. Pathogens 2021, 10, 344. https://doi.org/10.3390/pathogens10030344
Kandari D, Joshi H, Bhatnagar R. Zur: Zinc-Sensing Transcriptional Regulator in a Diverse Set of Bacterial Species. Pathogens. 2021; 10(3):344. https://doi.org/10.3390/pathogens10030344
Chicago/Turabian StyleKandari, Divya, Hemant Joshi, and Rakesh Bhatnagar. 2021. "Zur: Zinc-Sensing Transcriptional Regulator in a Diverse Set of Bacterial Species" Pathogens 10, no. 3: 344. https://doi.org/10.3390/pathogens10030344
APA StyleKandari, D., Joshi, H., & Bhatnagar, R. (2021). Zur: Zinc-Sensing Transcriptional Regulator in a Diverse Set of Bacterial Species. Pathogens, 10(3), 344. https://doi.org/10.3390/pathogens10030344