
WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
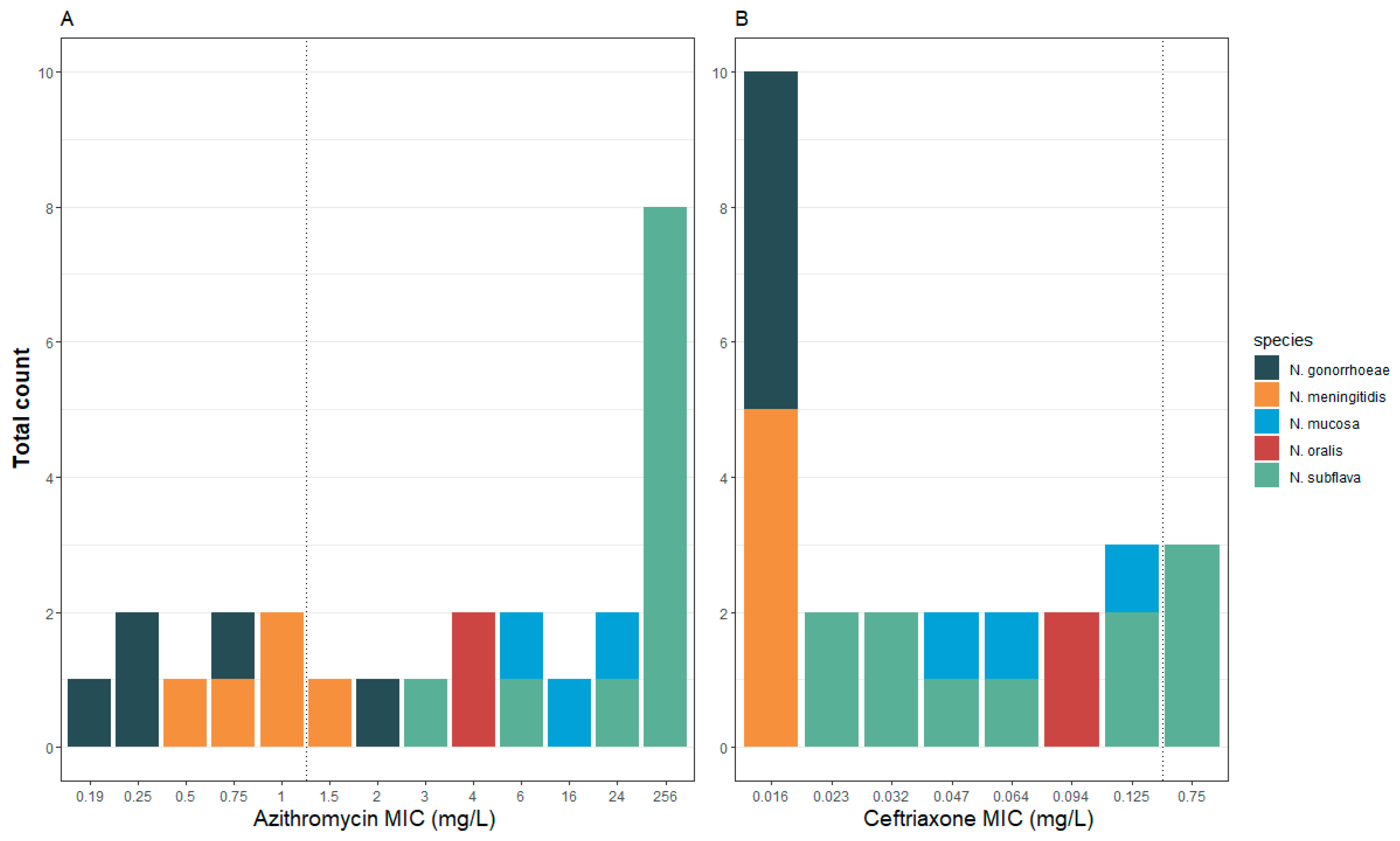
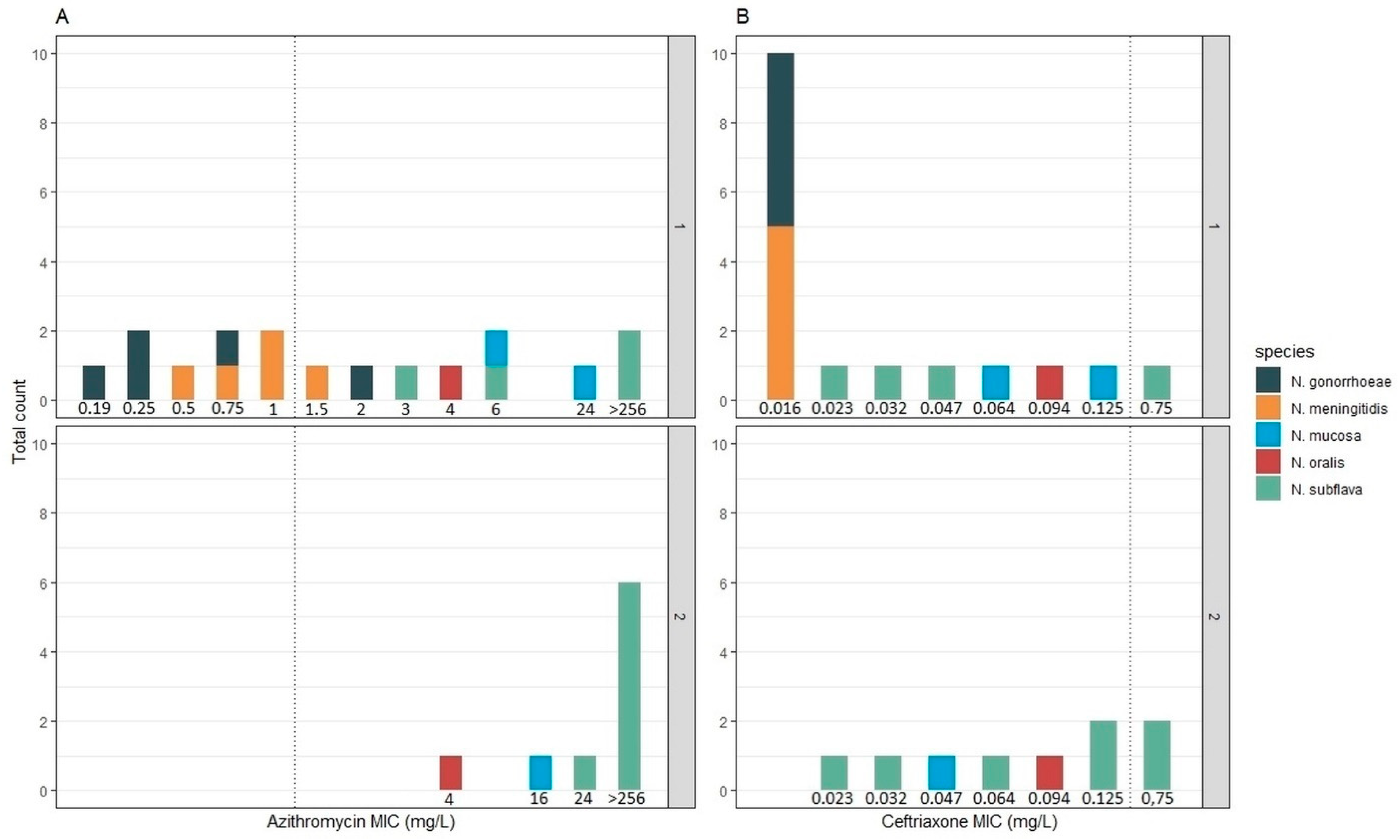
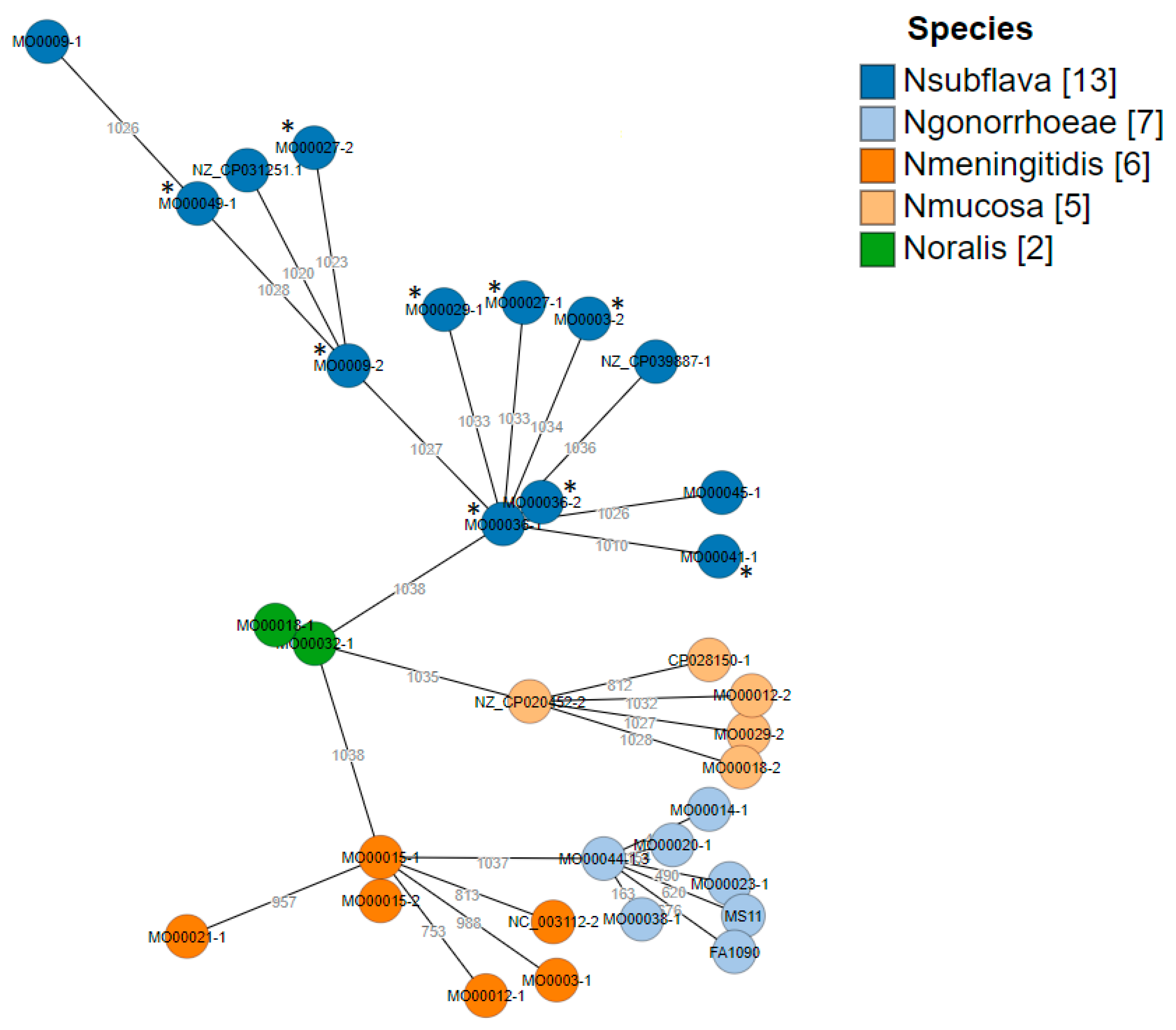
2.1. Characterization of Strain Collection
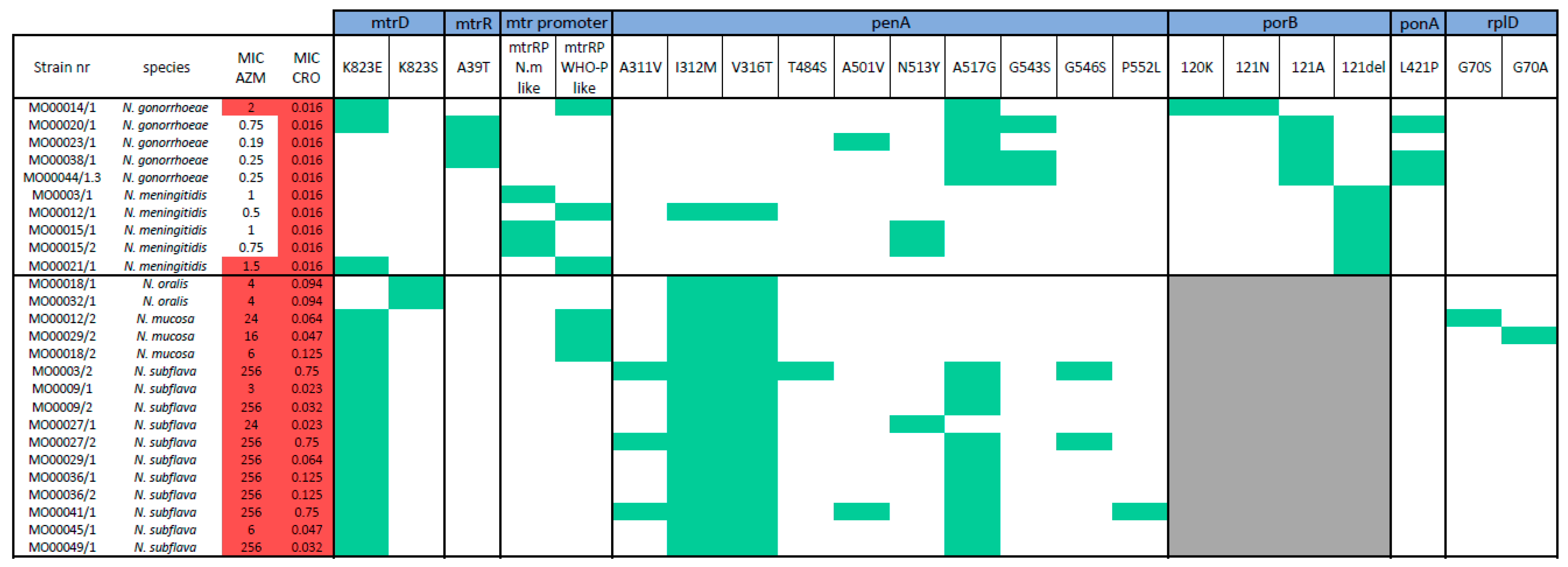
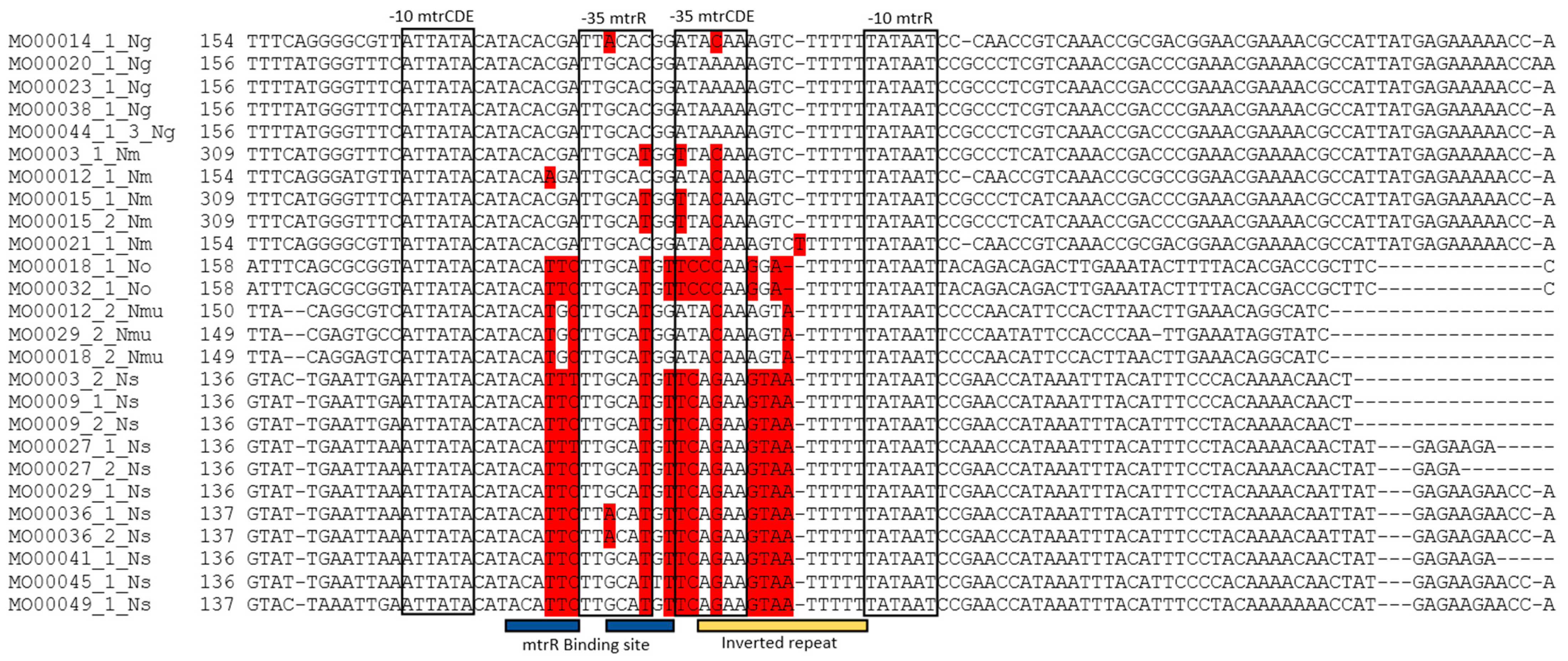
2.2. SNP Determination
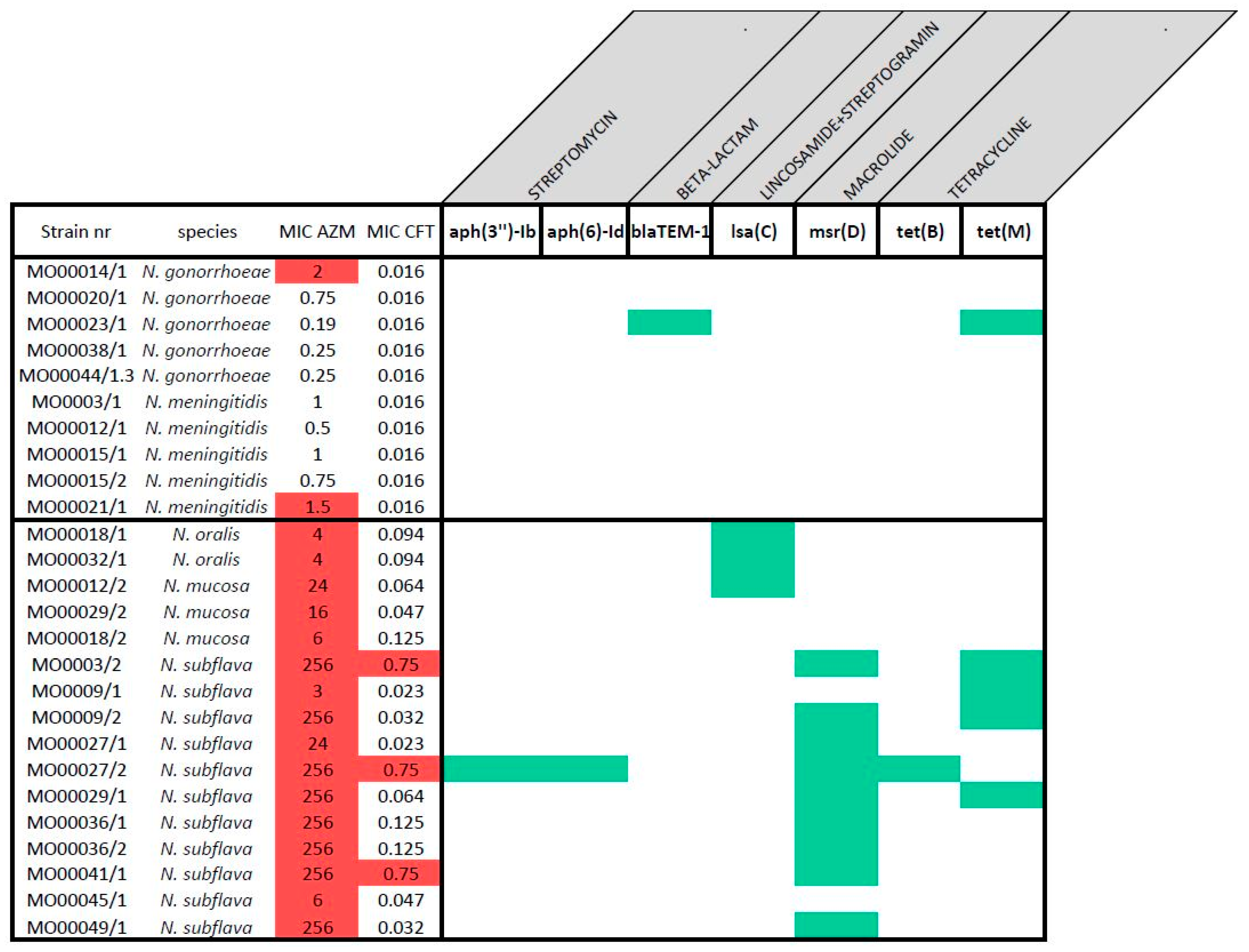
2.3. AMR Gene Screening
2.4. Core Genome (cg) MLST
3. Discussion
4. Materials and Methods
4.1. Strain Collection and MIC Determination
4.2. Whole Genome Sequencing
4.3. RAM Analysis and AMR Gene Screening
4.4. Multiple Linear Regression Models of Azithromycin MIC Prediction in Neisseria
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Unemo, M.; Bradshaw, C.S.; Hocking, J.S.; de Vries, H.J.C.; Francis, S.C.; Mabey, D.; Marrazzo, J.M.; Sonder, G.J.B.; Schwebke, J.R.; Hoornenborg, E.; et al. Sexually transmitted infections: Challenges ahead. Lancet Infect. Dis. 2017, 17, e235–e279. [Google Scholar] [CrossRef]
- Eyre, D.W.; Sanderson, N.D.; Lord, E.; Regisford-Reimmer, N.; Chau, K.; Barker, L.; Morgan, M.; Newnham, R.; Golparian, D.; Unemo, M.; et al. Gonorrhoea treatment failure caused by a Neisseria gonorrhoeae strain with combined ceftriaxone and high-level azithromycin resistance, England, February 2018. Eurosurveillance 2018, 23, 1800323. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.; Sawatzky, P.; Liu, G.; Allen, V.; Lefebvre, B.; Hoang, L.; Drews, S.; Horsman, G.; Wylie, J.; Haldane, D.; et al. Decline in Decreased Cephalosporin Susceptibility and Increase in Azithromycin Resistance in Neisseria gonorrhoeae, Canada. Emerg. Infect. Dis. 2016, 22, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Xue, J.; Chen, Y.; Chen, S.; Wang, Q.; Zhang, C.; Wu, S.; Lv, H.; Yu, Y.; van der Veen, S. Increasing prevalence of Neisseria gonorrhoeae with decreased susceptibility to ceftriaxone and resistance to azithromycin in Hangzhou, China (2015–17). J. Antimicrob. Chemother. 2018, 74, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.A.; Raisman, J.C.; Wong, N.H.; Hudson, A.O.; Wadsworth, C.B. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics 2020, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Higashi, D.; Goytia, M.; Rendón, M.A.; Pilligua-Lucas, M.; Bronnimann, M.; McLean, J.A.; Duncan, J.; Trees, D.; Jerse, A.E.; et al. Commensal Neisseria Kill Neisseria gonorrhoeae through a DNA-Dependent Mechanism. Cell Host Microbe 2019, 26, 228–239.e8. [Google Scholar] [CrossRef]
- Ito, M.; Deguchi, T.; Mizutani, K.-S.; Yasuda, M.; Yokoi, S.; Ito, S.-I.; Takahashi, Y.; Ishihara, S.; Kawamura, Y.; Ezaki, T. Emergence and Spread of Neisseria gonorrhoeae Clinical Isolates Harboring Mosaic-Like Structure of Penicillin-Binding Protein 2 in Central Japan. Antimicrob. Agents Chemother. 2005, 49, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, C.B.; Arnold, B.J.; Sater, M.R.A.; Grad, Y.H. Azithromycin Resistance through Interspecific Acquisition of an Epistasis-Dependent Efflux Pump Component and Transcriptional Regulator in Neisseria gonorrhoeae. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igawa, G.; Yamagishi, Y.; Lee, K.-I.; Dorin, M.; Shimuta, K.; Suematsu, H.; Nakayama, S.-I.; Mikamo, H.; Unemo, M.; Ohnishi, M. Neisseria cinerea with High Ceftriaxone MIC Is a Source of Ceftriaxone and Cefixime Resistance-Mediating penA Sequences in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2018, 62, e02069-17. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.V.; Pham, L.Q.; Nguyen, H.T.; Nguyen, M.X.B.; Nguyen, T.V.; May, F.; Le, G.M.; Klausner, J.D. Decreased Cephalosporin Susceptibility of Oropharyngeal Neisseria Species in Antibiotic-using Men Who Have Sex with Men in Hanoi, Vietnam. Clin. Infect. Dis. 2020, 70, 1169–1175. [Google Scholar] [CrossRef]
- Kenyon, C.; Buyze, J.; Wi, T. Antimicrobial Consumption and Susceptibility of Neisseria gonorrhoeae: A Global Ecological Analysis. Front. Med. 2018, 5, 329. [Google Scholar] [CrossRef]
- Tsoumanis, A.; Hens, N.; Kenyon, C.R. Is Screening for Chlamydia and Gonorrhea in Men Who Have Sex with Men Associated with Reduction of the Prevalence of these Infections? A Systematic Review of Observational Studies. Sex. Transm. Dis. 2018, 45, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Dorey, R.B.; Theodosiou, A.A.; Read, R.C.; Jones, C.E. The nonpathogenic commensal Neisseria. Curr. Opin. Infect. Dis. 2019, 32, 490–496. [Google Scholar] [CrossRef]
- Kenyon, C.R.; Schwartz, I.S. Effects of Sexual Network Connectivity and Antimicrobial Drug Use on Antimicrobial Resistance in Neisseria gonorrhoeae. Emerg. Infect. Dis. 2018, 24, 1195–1203. [Google Scholar] [CrossRef] [Green Version]
- Laumen, J.G.E.; van Dijck, C.; Abdellati, S.; Manoharan-Basil, S.S.; de Baetselier, I.; Martiny, D.; Crucitti, T.; Kenyon, C. Markedly Reduced Azithromycin and Ceftriaxone Susceptibility in Commensal Neisseria Species in Clinical Samples from Belgian Men Who Have Sex with Men. Clin. Infect. Dis. 2021, 72, 363–364. [Google Scholar] [CrossRef]
- Lewis, D.A. The role of core groups in the emergence and dissemination of antimicrobial-resistant N. gonorrhoeae. Sex. Transm. Infect. 2013, 89, iv47–iv51. [Google Scholar] [CrossRef]
- De Baetselier, I.; Kenyon, C.; Berghe, W.V.; Smet, H.; Wouters, K.; Bossche, D.V.D.; Vuylsteke, B.; Crucitti, T. An alarming high prevalence of resistance-associated mutations to macrolides and fluoroquinolones in Mycoplasma genitalium in Belgium: Results from samples collected between 2015 and 2018. Sex. Transm. Infect. 2020, 1–7. [Google Scholar] [CrossRef]
- Kenyon, C. Prevalence of macrolide resistance in Treponema pallidum is associated with macrolide consumption. J. Med Microbiol. 2019, 68, 119–123. [Google Scholar] [CrossRef]
- Vuylsteke, B.; Reyniers, T.; de Baetselier, I.; Nöstlinger, C.; Crucitti, T.; Buyze, J.; Kenyon, C.; Wouters, K.; Laga, M. Daily and event-driven pre-exposure prophylaxis for men who have sex with men in Belgium: Results of a prospective cohort measuring adherence, sexual behaviour and STI incidence. J. Int. AIDS Soc. 2019, 22, e25407. [Google Scholar] [CrossRef] [PubMed]
- Furuya, R.; Tanaka, M.; Onoye, Y.; Kanayama, A.; Saika, T.; Iyoda, T.; Tatewaki, M.; Matsuzaki, K.; Kobayashi, I. Antimicrobial resistance in clinical isolates of Neisseria subflava from the oral cavities of a Japanese population. J. Infect. Chemother. 2007, 13, 302–304. [Google Scholar] [CrossRef]
- Bennett, J.S.; Watkins, E.R.; Jolley, K.A.; Harrison, O.B.; Maiden, M.C.J. Identifying Neisseria Species by Use of the 50S Ribosomal Protein L6 (rplF) Gene. J. Clin. Microbiol. 2014, 52, 1375–1381. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing Breakpoint tables for Interpretation of MICs and Zone Diameters. Version 11.0. 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_11.0_Breakpoint_Tables.pdf (accessed on 11 January 2021).
- Olesky, M.; Hobbs, M.; Nicholas, R.A. Identification and Analysis of Amino Acid Mutations in Porin IB That Mediate Intermediate-Level Resistance to Penicillin and Tetracycline in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2002, 46, 2811–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyre, D.W.; de Silva, D.; Cole, K.; Peters, J.; Cole, M.J.; Grad, Y.H.; Demczuk, W.; Martin, I.; Mulvey, M.R.; Crook, D.W.; et al. WGS to predict antibiotic MICs for Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2017, 72, 1937–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.-K.; Martin, I.; Liu, G.; Bryden, L. Mutation in 23S rRNA Associated with Macrolide Resistance in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2002, 46, 3020–3025. [Google Scholar] [CrossRef] [Green Version]
- Allen, V.G.; Farrell, D.J.; Rebbapragada, A.; Tan, J.; Tijet, N.; Perusini, S.J.; Towns, L.; Lo, S.; Low, D.E.; Melano, R.G. Molecular Analysis of Antimicrobial Resistance Mechanisms in Neisseria gonorrhoeae Isolates from Ontario, Canada. Antimicrob. Agents Chemother. 2010, 55, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, L.K.R.; Edwards, T.A.; O’Neill, A.J. ABC-F Proteins Mediate Antibiotic Resistance through Ribosomal Protection. mBio 2016, 7, e01975-15. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N. The ABC of Ribosome-Related Antibiotic Resistance. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef]
- Banks, D.J.; Porcella, S.F.; Barbian, K.D.; Martin, J.M.; Musser, J.M. Structure and Distribution of an Unusual Chimeric Genetic Element Encoding Macrolide Resistance in Phylogenetically Diverse Clones of Group A. Streptococcus. J. Infect. Dis. 2003, 188, 1898–1908. [Google Scholar] [CrossRef]
- Del Grosso, M.; Camilli, R.; Iannelli, F.; Pozzi, G.; Pantosti, A. The mef(E)-Carrying Genetic Element (mega) of Streptococcus pneumoniae: Insertion Sites and Association with Other Genetic Elements. Antimicrob. Agents Chemother. 2006, 50, 3361–3366. [Google Scholar] [CrossRef] [Green Version]
- Gay, K.; Stephens, D.S. Structure and Dissemination of a Chromosomal Insertion Element Encoding Macrolide Efflux in Streptococcus pneumoniae. J. Infect. Dis. 2001, 184, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Fostier, C.R.; Monlezun, L.; Ousalem, F.; Singh, S.; Hunt, J.F.; Boël, G. ABC-F translation factors: From antibiotic resistance to immune response. FEBS Lett. 2020, 595, 675–706. [Google Scholar] [CrossRef]
- Daly, M.M.; Doktor, S.; Flamm, R.; Shortridge, D. Characterization and Prevalence of MefA, MefE, and the Associated msr (D) Gene in Streptococcus pneumoniae Clinical Isolates. J. Clin. Microbiol. 2004, 42, 3570–3574. [Google Scholar] [CrossRef] [Green Version]
- Iannelli, F.; Santoro, F.; Santagati, M.; Docquier, J.-D.; Lazzeri, E.; Pastore, G.; Cassone, M.; Oggioni, M.R.; Rossolini, G.M.; Stefani, S.; et al. Type M Resistance to Macrolides Is Due to a Two-Gene Efflux Transport System of the ATP-Binding Cassette (ABC) Superfamily. Front. Microbiol. 2018, 9, 1670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tatsuno, I.; Okada, R.; Hata, N.; Matsumoto, M.; Isaka, M.; Isobe, K.-I.; Hasegawa, T. Predominant role of msr(D) over mef(A) in macrolide resistance in Streptococcus pyogenes. Microbiology 2016, 162, 46–52. [Google Scholar] [CrossRef]
- Nunez-Samudio, V.; Chesneau, O. Functional interplay between the ATP binding cassette Msr(D) protein and the membrane facilitator superfamily Mef(E) transporter for macrolide resistance in Escherichia coli. Res. Microbiol. 2013, 164, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Rotman, E.; Seifert, H.S. The Genetics of Neisseria Species. Annu. Rev. Genet. 2014, 48, 405–431. [Google Scholar] [CrossRef]
- Knapp, J.S.; Johnson, S.R.; Zenilman, J.M.; Roberts, M.C.; Morse, S.A. High-level tetracycline resistance resulting from TetM in strains of Neisseria spp., Kingella denitrificans, and Eikenella corrodens. Antimicrob. Agents Chemother. 1988, 32, 765–767. [Google Scholar] [CrossRef] [Green Version]
- Morse, S.A.; Johnson, S.R.; Biddle, J.W.; Roberts, M.C. High-level tetracycline resistance in Neisseria gonorrhoeae is result of acquisition of streptococcal tetM determinant. Antimicrob. Agents Chemother. 1986, 30, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C.; Knapp, J.S. Host range of the conjugative 25.2-megadalton tetracycline resistance plasmid from Neisseria gonorrhoeae and related species. Antimicrob. Agents Chemother. 1988, 32, 488–491. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.C.; Mortimer, T.D.; Duckett, M.A.; Hicks, A.L.; Wheeler, N.E.; Sánchez-Busó, L.; Grad, Y.H. Increased power from conditional bacterial genome-wide association identifies macrolide resistance mutations in Neisseria gonorrhoeae. Nat. Commun. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Adriaenssens, N.; Coenen, S.; Versporten, A.; Muller, A.; Minalu, G.; Faes, C.; Vankerckhoven, V.; Aerts, M.; Hens, N.; Molenberghs, G.; et al. European Surveillance of Antimicrobial Consumption (ESAC): Outpatient antibiotic use in Europe (1997–2009). J. Antimicrob. Chemother. 2011, 66, vi3–vi12. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, C.; Buyze, J.; Spiteri, G.; Cole, M.J.; Unemo, M. Population-Level Antimicrobial Consumption Is Associated with Decreased Antimicrobial Susceptibility in Neisseria gonorrhoeae in 24 European Countries: An Ecological Analysis. J. Infect. Dis. 2019, 221, 1107–1116. [Google Scholar] [CrossRef]
- Deschepper, R.; Stichele, R.H.V.; Haaijer-Ruskamp, F.M. Cross-cultural differences in lay attitudes and utilisation of antibiotics in a Belgian and a Dutch city. Patient Educ. Couns. 2002, 48, 161–169. [Google Scholar] [CrossRef]
- Kenyon, C.; Manoharan-Basil, S.S. Cultural Drivers of Antibiotic Consumption in High-Income Countries: A Global Ecological Analysis. Microb. Drug Resist. 2020, 26, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Deschepper, R.; Grigoryan, L.; Lundborg, C.S.; Hofstede, G.; Cohen, J.; van der Kelen, G.; Deliens, L.; Haaijer-Ruskamp, F.M. Are cultural dimensions relevant for explaining cross-national differences in antibiotic use in Europe? BMC Heal. Serv. Res. 2008, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.; Fatti, G. Thank Martin Luther that ciprofloxacin could cure your gonorrhoea? Ecological association between Protestantism and antimicrobial consumption in 30 European countries. F1000Research 2020, 9, 1200. [Google Scholar] [CrossRef]
- Kenyon, C.; de Baetselier, I.; Wouters, K. Screening for STIs in PrEP cohorts results in high levels of antimicrobial consumption. Int. J. STD AIDS 2020, 31, 1215–1218. [Google Scholar] [CrossRef]
- Kenyon, C.; Manoharan-Basil, S.S.; van Dijck, C. Is there a resistance-threshold for macrolide consumption? Positive evidence from an ecological analysis of resistance data from Streptococcus pneumoniae, Treponema pallidum, and Mycoplasma genitalium. Microb. Drug Resist. 2021. [Google Scholar] [CrossRef]
- Seemann, T. Abricate. Available online: https://github.com/tseemann/abricate (accessed on 21 March 2021).
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Wick, R.R. Filtlong. Available online: https://github.com/rrwick (accessed on 21 March 2021).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Seqtk. Available online: https://github.com/lh3 (accessed on 21 March 2021).
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genome Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyre, D.E.; Golparian, D.; Unemo, M. Prediction of Minimum Inhibitory Concentrations of Antimicrobials for Neisseria gonorrhoeae Using Whole-Genome Sequencing. In Neisseria Gonorrhoeae: Methods and Protocols; Christodoulides, M., Ed.; Springer: New York, NY, USA, 2019; ISBN 978-1-4939-9495-3. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain nr | Visit 1 | Sampling site 2 | MIC AZM 3 | AZM S/R 4 | MIC CRO 3 | CRO S/R 4 |
|---|---|---|---|---|---|---|---|
| N. gonorrhoeae | MO00014/1 | 1 | Anal | 2 | R | <0.016 | S |
| N. gonorrhoeae | MO00020/1 | 1 | Anal | 0.75 | S | <0.016 | S |
| N. gonorrhoeae | MO00023/1 | 1 | Anal | 0.19 | S | 0.016 | S |
| N. gonorrhoeae | MO00038/1 | 1 | Anal | 0.25 | S | 0.016 | S |
| N. gonorrhoeae | MO00044/1.3 | 1 | Anal | 0.25 | S | 0.016 | S |
| N. meningitidis | MO0003/1 | 1 | Oral | 1 | S | <0.016 | S |
| N. meningitidis | MO00012/1 | 1 | Oral | 0.5 | S | <0.016 | S |
| N. meningitidis | MO00015/1 | 1 | Oral | 1 | S | <0.016 | S |
| N. meningitidis | MO00015/2 | 1 | Oral | 0.75 | S | <0.016 | S |
| N. meningitidis | MO00021/1 | 1 | Oral | 1.5 | R | <0.016 | S |
| N. oralis | MO00018/1 | 1 | Oral | 4 | R | 0.094 | S |
| N. oralis | MO00032/1 | 2 | Oral | 4 | R | 0.094 | S |
| N. mucosa | MO00012/2 | 1 | Oral | 24 | R | 0.064 | S |
| N. mucosa | MO00029/2 | 2 | Oral | 16 | R | 0.047 | S |
| N. mucosa | MO00018/2 | 1 | Oral | 6 | R | 0.125 | S |
| N. subflava | MO0003/2 | 1 | Oral | >256 | R | 0.75 | R |
| N. subflava | MO0009/1 | 1 | Oral | 3 | R | 0.023 | S |
| N. subflava | MO0009/2 | 1 | Oral | >256 | R | 0.032 | S |
| N. subflava | MO00027/1 | 2 | Oral | 24 | R | 0.023 | S |
| N. subflava | MO00027/2 | 2 | Oral | >256 | R | 0.75 | R |
| N. subflava | MO00029/1 | 2 | Oral | >256 | R | 0.064 | S |
| N. subflava | MO00036/1 | 2 | Oral | >256 | R | 0.125 | S |
| N. subflava | MO00036/2 | 2 | Oral | >256 | R | 0.125 | S |
| N. subflava | MO00041/1 | 2 | Oral | >256 | R | 0.75 | R |
| N. subflava | MO00045/1 | 1 | Oral | 6 | R | 0.047 | S |
| N. subflava | MO00049/1 | 2 | Oral | >256 | R | 0.032 | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Block, T.; Laumen, J.G.E.; Van Dijck, C.; Abdellati, S.; De Baetselier, I.; Manoharan-Basil, S.S.; Van den Bossche, D.; Kenyon, C. WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium. Pathogens 2021, 10, 384. https://doi.org/10.3390/pathogens10030384
de Block T, Laumen JGE, Van Dijck C, Abdellati S, De Baetselier I, Manoharan-Basil SS, Van den Bossche D, Kenyon C. WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium. Pathogens. 2021; 10(3):384. https://doi.org/10.3390/pathogens10030384
Chicago/Turabian Stylede Block, Tessa, Jolein Gyonne Elise Laumen, Christophe Van Dijck, Said Abdellati, Irith De Baetselier, Sheeba Santhini Manoharan-Basil, Dorien Van den Bossche, and Chris Kenyon. 2021. "WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium" Pathogens 10, no. 3: 384. https://doi.org/10.3390/pathogens10030384
APA Stylede Block, T., Laumen, J. G. E., Van Dijck, C., Abdellati, S., De Baetselier, I., Manoharan-Basil, S. S., Van den Bossche, D., & Kenyon, C. (2021). WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium. Pathogens, 10(3), 384. https://doi.org/10.3390/pathogens10030384