
Scanning Electron Microscopic Findings on Respiratory Organs of Some Naturally Infected Dromedary Camels with the Lineage-B of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Saudi Arabia—2018


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Identification of Some MERS-CoV Field Infected Animals
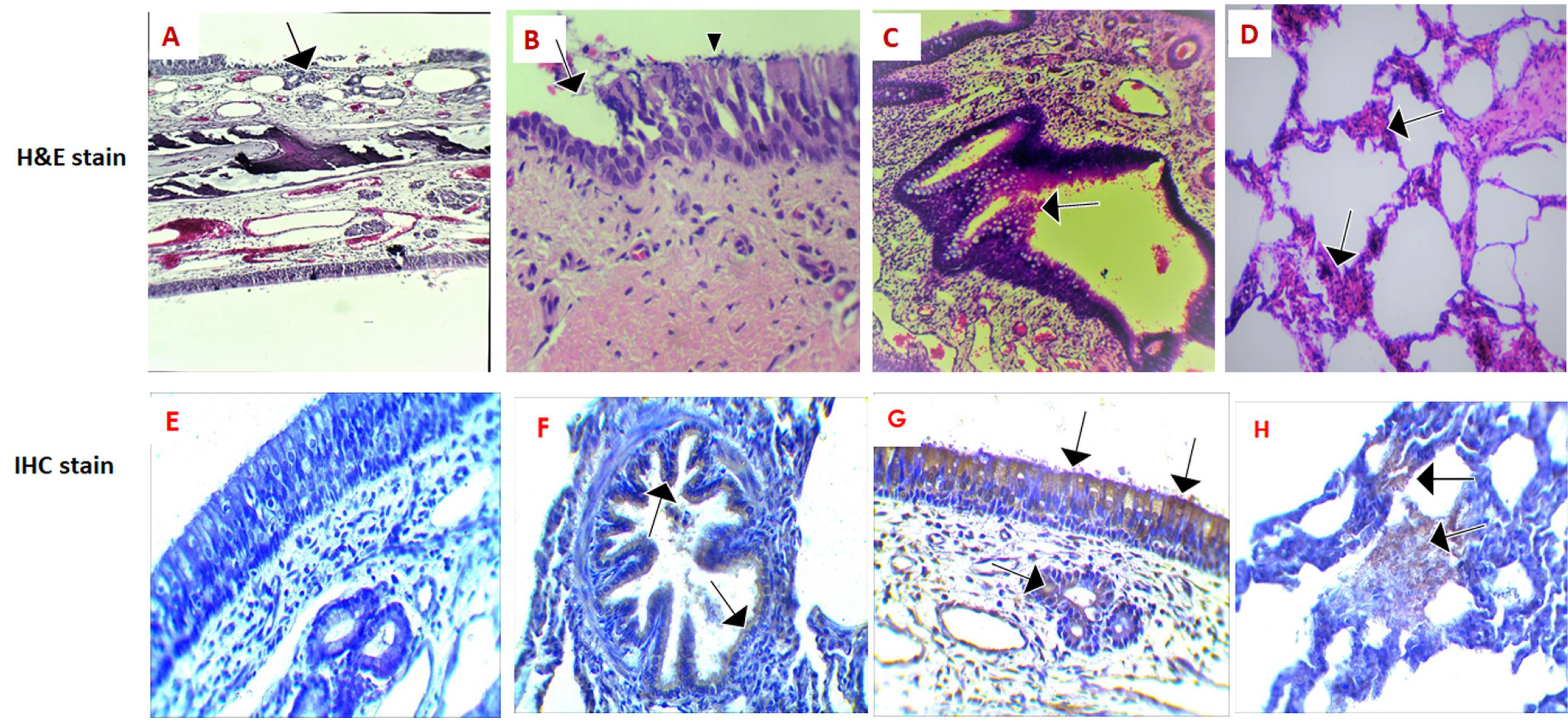
2.2. Histopathological and Immunohistochemical Changes in Some Respiratory Organs Obtained from MERS-CoV Field Infected Animals Using the Light Microscope
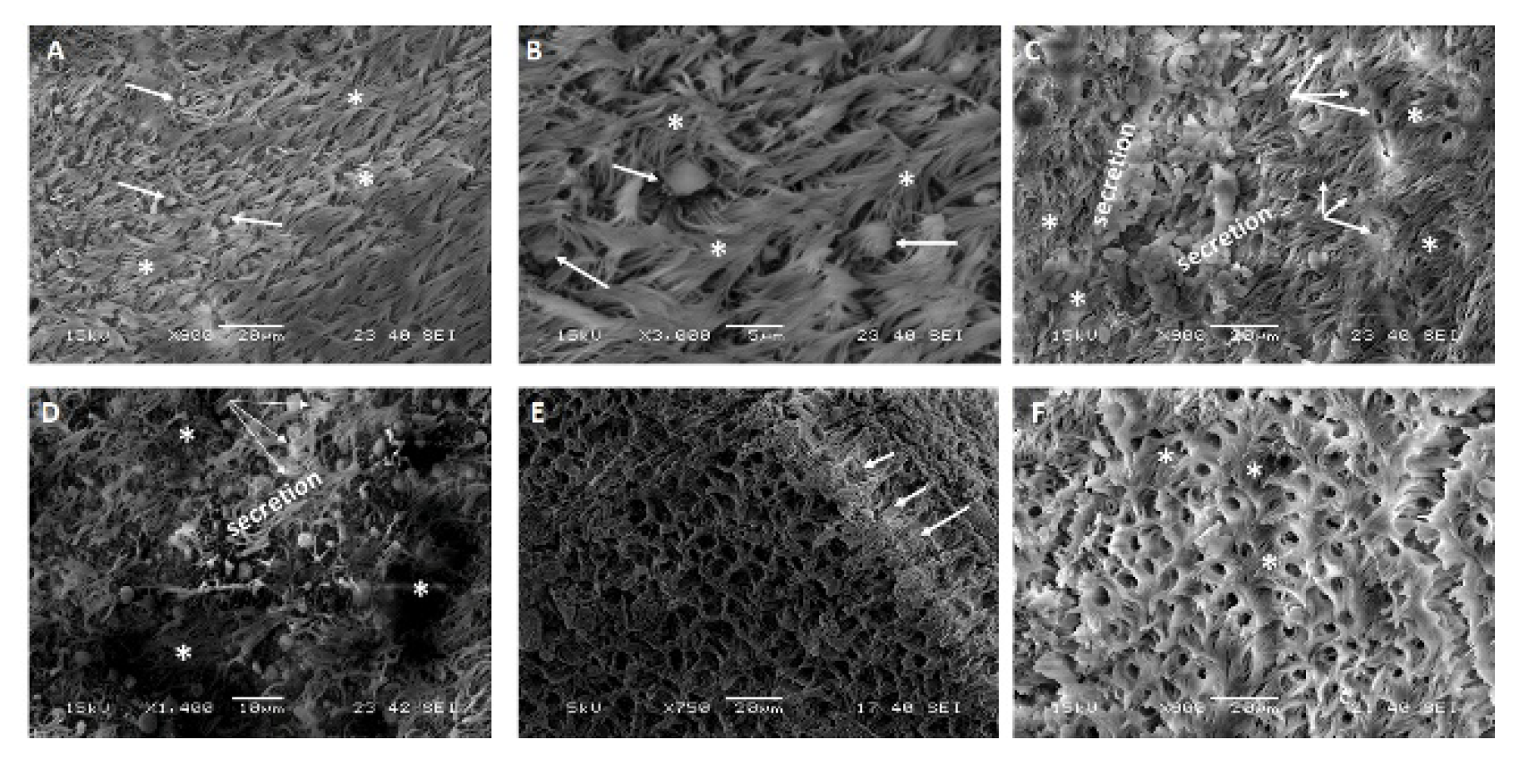
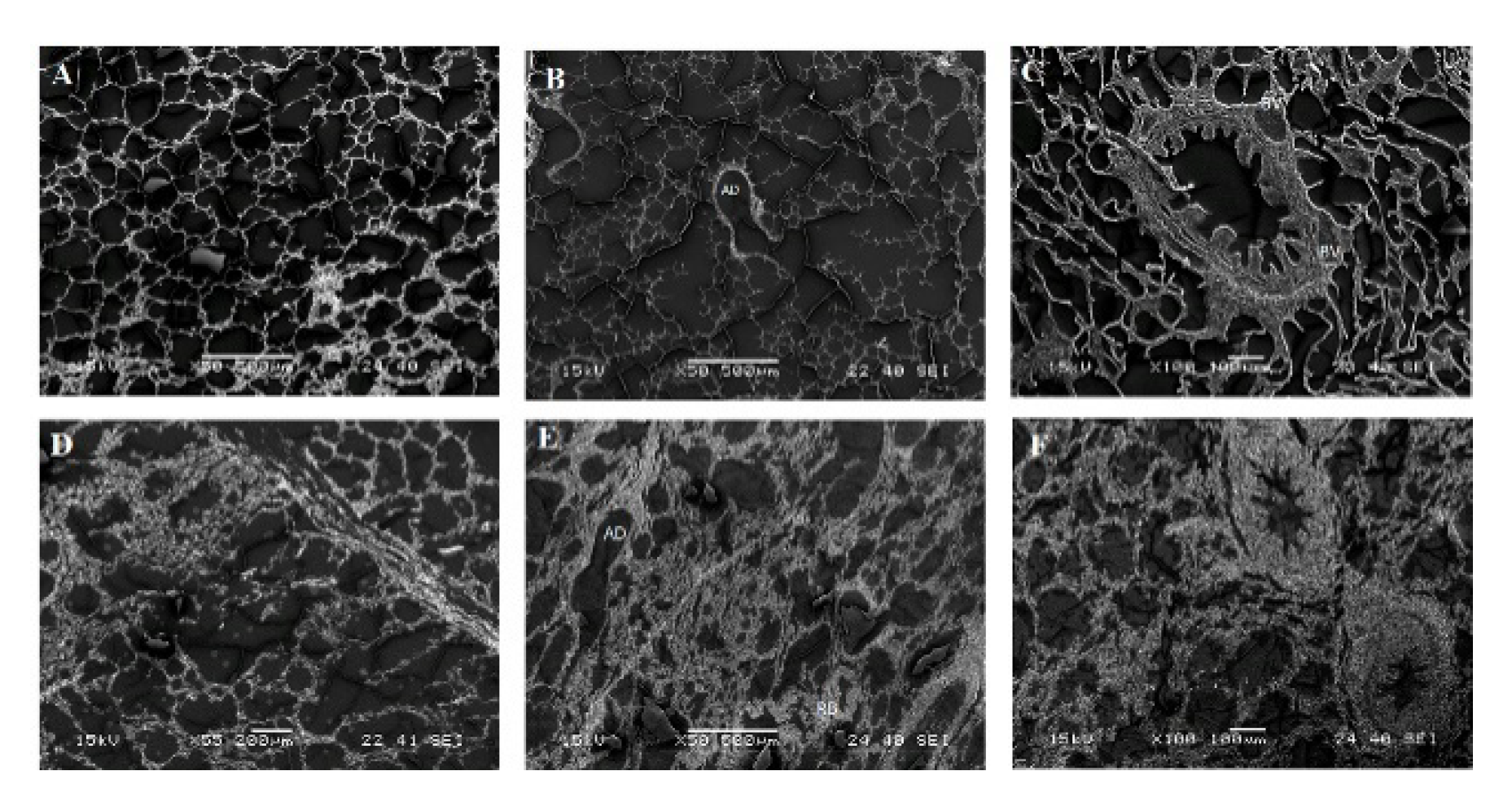
2.3. Scanning Electron Microscopy (SEM) on Respiratory Organs Obtained from MERS-CoV Field Infected Animals
2.3.1. Lesion Scoring and Pathological Alterations in the Nasal Turbinate of Some Field Infected Animals by the SEM
2.3.2. Lesion Scoring and Pathological Alterations in the Trachea of Some Field Infected Animals by the SEM
2.3.3. Alterations and some Pathological Findings in the Lung Tissues of Field Infected MERS-CoV Dromedary Camels by SEM
2.4. Identification of the MERS-CoV Lineage in Animals Admitted to This Abattoir
3. Discussion
4. Materials and Methods
4.1. Animal Selection
4.2. Histopathological Examination
4.3. Immunohistochemistry
4.4. RNA Extraction, Real-Time PCR, and Sequencing of MERS-CoV-S Gene
4.5. Scanning Electron Microscope
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Reusken, C.B.; Haagmans, B.L.; Muller, M.A.; Gutierrez, C.; Godeke, G.J.; Meyer, B.; Muth, D.; Raj, V.S.; Smits-De Vries, L.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Hemida, M.G.; Chu, D.K.; Poon, L.L.; Perera, R.A.; Alhammadi, M.A.; Ng, H.Y.; Siu, L.Y.; Guan, Y.; Alnaeem, A.; Peiris, M. MERS coronavirus in dromedary camel herd, Saudi Arabia. Emerg. Infect. Dis. 2014, 20, 1231–1234. [Google Scholar] [CrossRef] [Green Version]
- Hemida, M.G.; Perera, R.A.; Al Jassim, R.A.; Kayali, G.; Siu, L.Y.; Wang, P.; Chu, K.W.; Perlman, S.; Ali, M.A.; Alnaeem, A.; et al. Seroepidemiology of Middle East respiratory syndrome (MERS) coronavirus in Saudi Arabia (1993) and Australia (2014) and characterisation of assay specificity. Eurosurveillance 2014, 19, 20828. [Google Scholar] [CrossRef]
- Hemida, M.G.; Perera, R.A.; Wang, P.; Alhammadi, M.A.; Siu, L.Y.; Li, M.; Poon, L.L.; Saif, L.; Alnaeem, A.; Peiris, M. Middle East Respiratory Syndrome (MERS) coronavirus seroprevalence in domestic livestock in Saudi Arabia, 2010 to 2013. Eurosurveillance 2013, 18, 20659. [Google Scholar] [CrossRef]
- Khalafalla, A.I.; Lu, X.; Al-Mubarak, A.I.; Dalab, A.H.; Al-Busadah, K.A.; Erdman, D.D. MERS-CoV in Upper Respiratory Tract and Lungs of Dromedary Camels, Saudi Arabia, 2013–2014. Emerg. Infect. Dis. 2015, 21, 1153–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusof, M.F.; Eltahir, Y.M.; Serhan, W.S.; Hashem, F.M.; Elsayed, E.A.; Marzoug, B.A.; Abdelazim, A.S.; Bensalah, O.K.; Al Muhairi, S.S. Prevalence of Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels in Abu Dhabi Emirate, United Arab Emirates. Virus Genes 2015, 50, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirato, K.; Melaku, S.K.; Kawachi, K.; Nao, N.; Iwata-Yoshikawa, N.; Kawase, M.; Kamitani, W.; Matsuyama, S.; Tessema, T.S.; Sentsui, H. Middle East Respiratory Syndrome Coronavirus in Dromedaries in Ethiopia Is Antigenically Different From the Middle East Isolate EMC. Front. Microbiol. 2019, 10, 1326. [Google Scholar] [CrossRef] [PubMed]
- Crameri, G.; Durr, P.A.; Klein, R.; Foord, A.; Yu, M.; Riddell, S.; Haining, J.; Johnson, D.; Hemida, M.G.; Barr, J.; et al. Experimental Infection and Response to Rechallenge of Alpacas with Middle East Respiratory Syndrome Coronavirus. Emerg. Infect. Dis. 2016, 22, 1071–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodon, J.; Okba, N.M.A.; Te, N.; van Dieren, B.; Bosch, B.J.; Bensaid, A.; Segales, J.; Haagmans, B.L.; Vergara-Alert, J. Blocking transmission of Middle East respiratory syndrome coronavirus (MERS-CoV) in llamas by vaccination with a recombinant spike protein. Emerg. Microbes Infect. 2019, 8, 1593–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusken, C.B.; Schilp, C.; Raj, V.S.; De Bruin, E.; Kohl, R.H.; Farag, E.A.; Haagmans, B.L.; Al-Romaihi, H.; Le Grange, F.; Bosch, B.J.; et al. MERS-CoV Infection of Alpaca in a Region Where MERS-CoV is Endemic. Emerg. Infect. Dis. 2016, 22, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Adney, D.R.; van Doremalen, N.; Brown, V.R.; Bushmaker, T.; Scott, D.; de Wit, E.; Bowen, R.A.; Munster, V.J. Replication and shedding of MERS-CoV in upper respiratory tract of inoculated dromedary camels. Emerg. Infect. Dis. 2014, 20, 1999–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adney, D.R.; Bielefeldt-Ohmann, H.; Hartwig, A.E.; Bowen, R.A. Infection, Replication, and Transmission of Middle East Respiratory Syndrome Coronavirus in Alpacas. Emerg. Infect. Dis. 2016, 22, 1031–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, R.W.; Hemida, M.G.; Kayali, G.; Chu, D.K.; Poon, L.L.; Alnaeem, A.; Ali, M.A.; Tao, K.P.; Ng, H.Y.; Chan, M.C.; et al. Tropism and replication of Middle East respiratory syndrome coronavirus from dromedary camels in the human respiratory tract: An in-vitro and ex-vivo study. Lancet Respir. Med. 2014, 2, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, M.H.; Yoo, D.; Ojkic, D.; Barta, J.R. Virus shedding and serum antibody responses during experimental turkey coronavirus infections in young turkey poults. Avian Pathol. 2009, 38, 181–186. [Google Scholar] [CrossRef]
- Lin, Y.; Yan, X.; Cao, W.; Wang, C.; Feng, J.; Duan, J.; Xie, S. Probing the structure of the SARS coronavirus using scanning electron microscopy. Antivir. Ther. 2004, 9, 287–289. [Google Scholar]
- Zhang, Q.F.; Cui, J.M.; Huang, X.J.; Lin, W.; Tan, D.Y.; Xu, J.W.; Yang, Y.F.; Zhang, J.Q.; Zhang, X.; Li, H.; et al. Morphology and morphogenesis of severe acute respiratory syndrome (SARS)-associated virus. Acta Bioch. Bioph. Sin. 2003, 35, 587–591. [Google Scholar]
- Pesaresi, M.; Pirani, F.; Tagliabracci, A.; Valsecchi, M.; Procopio, A.D.; Busardo, F.P.; Graciotti, L. SARS-CoV-2 identification in lungs, heart and kidney specimens by transmission and scanning electron microscopy. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5186–5188. [Google Scholar] [CrossRef]
- Dougbag, A.E.; Berg, R.; Kassem, A.; Hemmoda, A.; Osman, A. Electron microscopic study on the prenatal development of the tracheal epithelium in the one-humped camel (Camelus dromedarius). Z. Mikrosk. Anat. Forsch. 1984, 98, 832–844. [Google Scholar]
- Raji, A.R.; Naserpour, M. Light and electron microscopic studies of the trachea in the one-humped camel (Camelus dromedarius). Anat. Histol. Embryol. 2007, 36, 10–13. [Google Scholar] [CrossRef]
- Haverkamp, A.K.; Lehmbecker, A.; Spitzbarth, I.; Widagdo, W.; Haagmans, B.L.; Segales, J.; Vergara-Alert, J.; Bensaid, A.; van den Brand, J.M.A.; Osterhaus, A.; et al. Experimental infection of dromedaries with Middle East respiratory syndrome-Coronavirus is accompanied by massive ciliary loss and depletion of the cell surface receptor dipeptidyl peptidase 4. Sci. Rep. 2018, 8, 9778. [Google Scholar] [CrossRef]
- Alnaeem, A.; Kasem, S.; Qasim, I.; Al-Doweriej, A.; Al-Houfufi, A.; Alwazan, A.; Albadrani, A.; Alshaammari, K.; Refaat, M.; Al-Shabebi, A.; et al. Some clinical and pathological observations on the naturally infected dromedary camels with the Middle East respiratory syndrome coronavirus in Saudi Arabia 2018–2019. Vet. Q. 2020, 40, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Ba Abduallah, M.M.; Hemida, M.G. Comparative analysis of the genome structure and organization of the Middle East respiratory syndrome coronavirus (MERS-CoV) 2012 to 2019 revealing evidence for virus strain barcoding, zoonotic transmission, and selection pressure. Rev. Med. Virol. 2021, 31, 1–12. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Hui, K.P.Y.; Perera, R.; Miguel, E.; Niemeyer, D.; Zhao, J.; Channappanavar, R.; Dudas, G.; Oladipo, J.O.; Traore, A.; et al. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc. Natl. Acad. Sci. USA 2018, 115, 3144–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemida, M.G.; Chu, D.K.W.; Chor, Y.Y.; Cheng, S.M.S.; Poon, L.L.M.; Alnaeem, A.; Peiris, M. Phylogenetic Analysis of MERS-CoV in a Camel Abattoir, Saudi Arabia, 2016–2018. Emerg. Infect. Dis. 2020, 26, 3089–3091. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Poon, L.L.; Gomaa, M.M.; Shehata, M.M.; Perera, R.A.; Abu Zeid, D.; El Rifay, A.S.; Siu, L.Y.; Guan, Y.; Webby, R.J.; et al. MERS coronaviruses in dromedary camels, Egypt. Emerg. Infect. Dis. 2014, 20, 1049–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeil, A.; Gomaa, M.; Nageh, A.; Shehata, M.M.; Kayed, A.E.; Sabir, J.S.M.; Abiadh, A.; Jrijer, J.; Amr, Z.; Said, M.A.; et al. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Dromedary Camels in Africa and Middle East. Viruses 2019, 11, 717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adney, D.R.; Letko, M.; Ragan, I.K.; Scott, D.; van Doremalen, N.; Bowen, R.A.; Munster, V.J. Bactrian camels shed large quantities of Middle East respiratory syndrome coronavirus (MERS-CoV) after experimental infection. Emerg. Microbes Infect. 2019, 8, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Iovannitti, B.; Pirie, H.M.; Wright, N.G. Scanning electron microscopic study of the lower respiratory tract in calves and adult cattle. Res. Vet. Sci. 1985, 38, 80–87. [Google Scholar] [CrossRef]
- Blyth, D.I.; Pedrick, M.S.; Savage, T.J.; Bright, H.; Beesley, J.E.; Sanjar, S. Induction, duration, and resolution of airway goblet cell hyperplasia in a murine model of atopic asthma: Effect of concurrent infection with respiratory syncytial virus and response to dexamethasone. Am. J. Respir. Cell Mol. Biol. 1998, 19, 38–54. [Google Scholar] [CrossRef]
- Argyropoulos, K.V.; Serrano, A.; Hu, J.; Black, M.; Feng, X.; Shen, G.; Call, M.; Kim, M.J.; Lytle, A.; Belovarac, B.; et al. Association of Initial Viral Load in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Patients with Outcome and Symptoms. Am. J. Pathol. 2020, 190, 1881–1887. [Google Scholar] [CrossRef] [PubMed]
- Alnaeem, A.; Kasem, S.; Qasim, I.; Al-Doweriej, A.; Refaat, M.; Al-Shabebi, A.; Hemida, M.G. The dipeptidyl peptidase-4 expression in some MERS-CoV naturally infected dromedary camels in Saudi Arabia 2018–2019. VirusDisease 2020, 31, 200–203. [Google Scholar] [CrossRef]
- Haverkos, H.W. Polymicrobial Diseases. Emerg. Infect. Dis. 2003, 9, 141. [Google Scholar] [CrossRef]
- Gohy, S.; Carlier, F.M.; Fregimilicka, C.; Detry, B.; Lecocq, M.; Ladjemi, M.Z.; Verleden, S.; Hoton, D.; Weynand, B.; Bouzin, C.; et al. Altered generation of ciliated cells in chronic obstructive pulmonary disease. Sci. Rep. 2019, 9, 17963. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.K.; Satapathy, A.; Naidu, M.M.; Mukhopadhyay, S.; Sharma, S.; Barton, L.M.; Stroberg, E.; Duval, E.J.; Pradhan, D.; Tzankov, A.; et al. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) and coronavirus disease 19 (COVID-19)—Anatomic pathology perspective on current knowledge. Diagn. Pathol. 2020, 15, 103. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, B.; Sharma, G.D.; Singh, S.; Dadhich, R. Incidence and pathology of bronchopneumonia in camel. Indian J. Vet. Pathol. 2006, 30, 39–41. [Google Scholar]
- Kasem, S.; Qasim, I.; Al-Hufofi, A.; Hashim, O.; Alkarar, A.; Abu-Obeida, A.; Gaafer, A.; Elfadil, A.; Zaki, A.; Al-Romaihi, A.; et al. Cross-sectional study of MERS-CoV-specific RNA and antibodies in animals that have had contact with MERS patients in Saudi Arabia. J. Infect. Public Health 2018, 11, 331–338. [Google Scholar] [CrossRef]
- Hemida, M.G.; Ali, A.M.; Alnaeem, A. The Middle East respiratory syndrome coronavirus (MERS-CoV) nucleic acids detected in the saliva and conjunctiva of some naturally infected dromedary camels in Saudi Arabia-2019. Zoonoses Public Health 2021. [Google Scholar] [CrossRef]
- Hemida, M.G.; Ali, M.; Alhammadi, M.; Alnaeem, A. The Middle East respiratory syndrome coronavirus in the breath of some infected dromedary camels (Camelus dromedarius). Epidemiol. Infect. 2020, 148, e247. [Google Scholar] [CrossRef]
- Hemida, M.G.; Waheed, M.; Ali, A.M.; Alnaeem, A. Detection of the Middle East respiratory syndrome coronavirus in dromedary camel’s seminal plasma in Saudi Arabia 2015–2017. Transbound. Emerg. Dis. 2020, 67, 2609–2614. [Google Scholar] [CrossRef]
- Briese, T.; Mishra, N.; Jain, K.; Zalmout, I.S.; Jabado, O.J.; Karesh, W.B.; Daszak, P.; Mohammed, O.B.; Alagaili, A.N.; Lipkin, W.I. Middle East respiratory syndrome coronavirus quasispecies that include homologues of human isolates revealed through whole-genome analysis and virus cultured from dromedary camels in Saudi Arabia. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Nasal Turbinate | Trachea | ||||
|---|---|---|---|---|---|---|
| Number of animal | 1 | 2 | 3 | 1 | 2 | 3 |
| Ciliary loss | + | +++ | - | - | + | ++ |
| Goblet cell hyperplasia | +++ | ++ | - | - | + | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alnaeem, A.; Kasem, S.; Qasim, I.; Refaat, M.; Alhufufi, A.N.; Al-Doweriej, A.; Al-Shabebi, A.; Hereba, A.-E.R.T.; Hemida, M.G. Scanning Electron Microscopic Findings on Respiratory Organs of Some Naturally Infected Dromedary Camels with the Lineage-B of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Saudi Arabia—2018. Pathogens 2021, 10, 420. https://doi.org/10.3390/pathogens10040420
Alnaeem A, Kasem S, Qasim I, Refaat M, Alhufufi AN, Al-Doweriej A, Al-Shabebi A, Hereba A-ERT, Hemida MG. Scanning Electron Microscopic Findings on Respiratory Organs of Some Naturally Infected Dromedary Camels with the Lineage-B of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Saudi Arabia—2018. Pathogens. 2021; 10(4):420. https://doi.org/10.3390/pathogens10040420
Chicago/Turabian StyleAlnaeem, Abdelmohsen, Samy Kasem, Ibrahim Qasim, Mohamed Refaat, Ali Nasser Alhufufi, Ali Al-Doweriej, Abdulkareem Al-Shabebi, Abd-El Rahman Taha Hereba, and Maged Gomaa Hemida. 2021. "Scanning Electron Microscopic Findings on Respiratory Organs of Some Naturally Infected Dromedary Camels with the Lineage-B of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Saudi Arabia—2018" Pathogens 10, no. 4: 420. https://doi.org/10.3390/pathogens10040420
APA StyleAlnaeem, A., Kasem, S., Qasim, I., Refaat, M., Alhufufi, A. N., Al-Doweriej, A., Al-Shabebi, A., Hereba, A.-E. R. T., & Hemida, M. G. (2021). Scanning Electron Microscopic Findings on Respiratory Organs of Some Naturally Infected Dromedary Camels with the Lineage-B of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Saudi Arabia—2018. Pathogens, 10(4), 420. https://doi.org/10.3390/pathogens10040420