
Microbial Pathogenicity in Space



Abstract
:1. Introduction
1.1. How Do Microorganisms Get to Space?
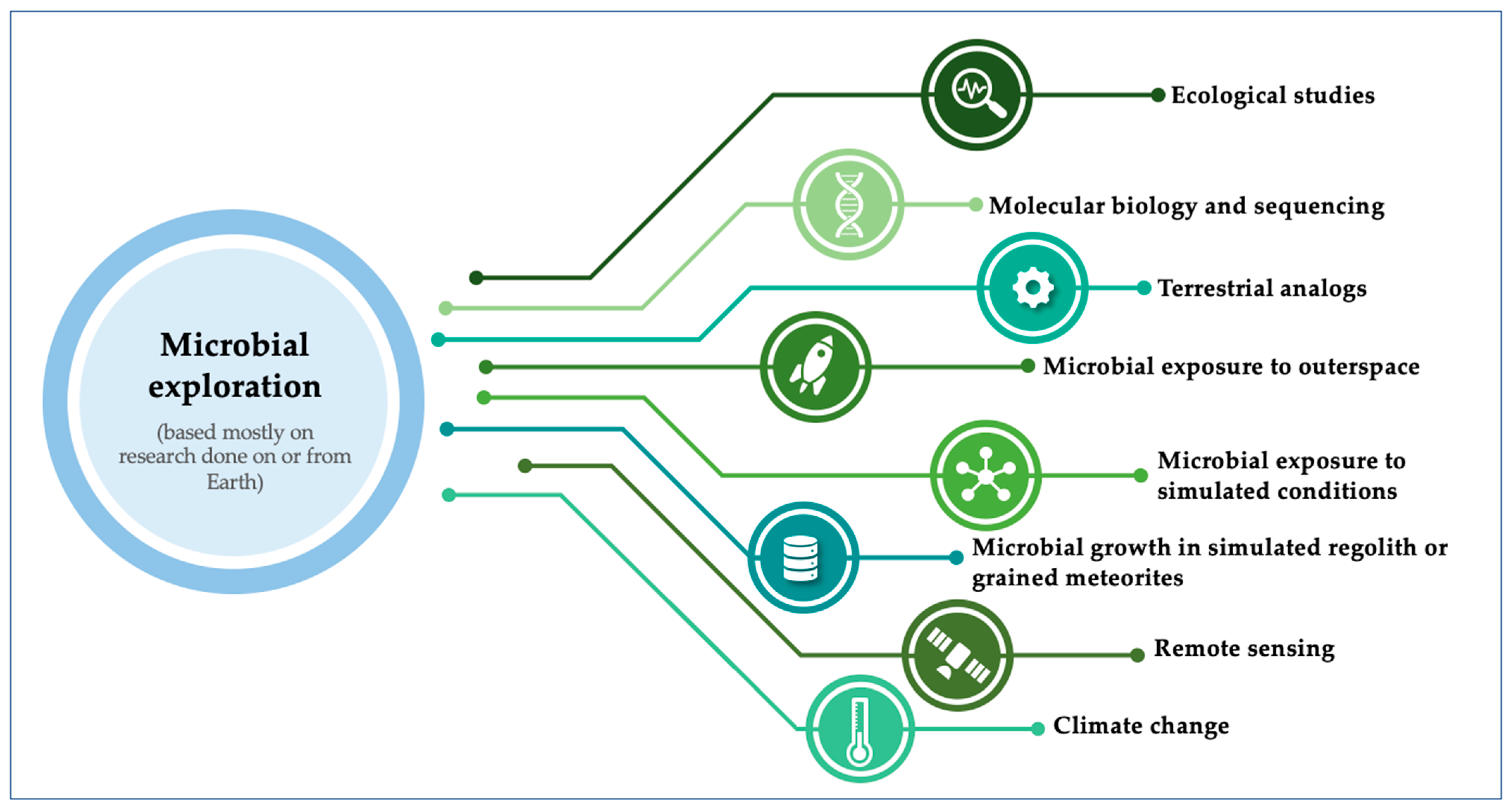
1.2. How Can We Study the Potential Pathogenicity of Microorganisms in Space Exploration?
1.2.1. Ecological Studies
1.2.2. Molecular Biology and Sequencing
1.2.3. Terrestrial Analogues
1.2.4. Microbial Exposure to Outer Space
1.2.5. Microbial Exposure to Simulated Conditions
1.2.6. Microbial Growth in Simulated Regoliths or Grained Meteorites
1.2.7. Remote Sensing
1.2.8. Climate Change
1.3. How do Microorganisms React to Space Conditions?
1.3.1. Bacteria
1.3.2. Fungi
1.3.3. Other Microorganisms
1.3.4. Examples of Studies Made So Far to Understand the Space Adaptability of Microorganisms
1.4. Can There Be Any Dangers of Exposure and Return to Common Environments?
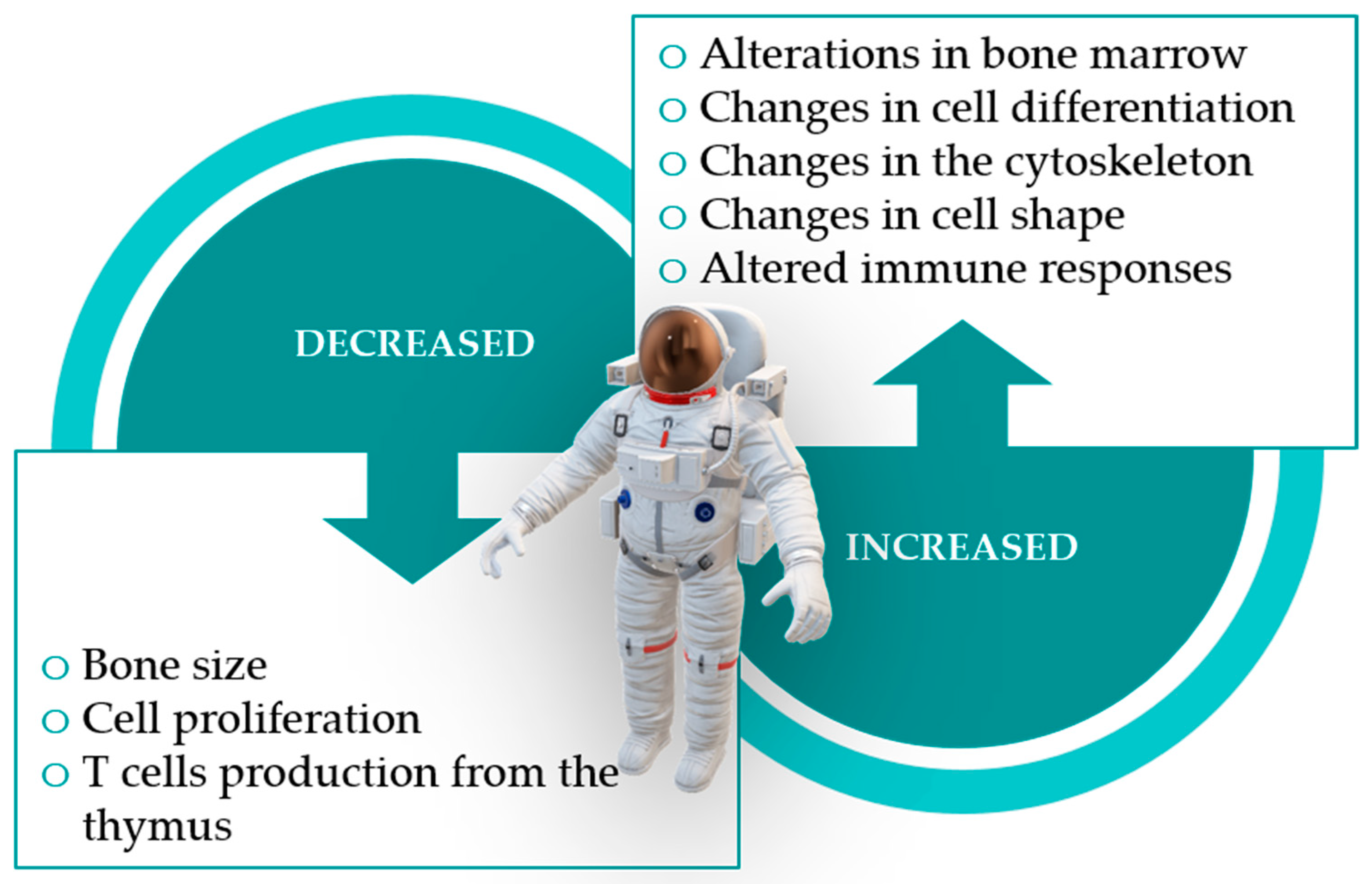
1.5. What Are the Effects and Impact of Space Exposure on Humans?
1.5.1. Development of Acquired Immunity
1.5.2. Immune Cell Responses
1.5.3. Immunity and Onset of Disease
1.5.4. Changes in Microbiome
1.5.5. Limitations and Countermeasures
2. Overview and Future Developments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pedrós-Alió, C.; Simó, R. Studying marine microorganisms from space. Int. Microbiol. 2002, 5, 195–200. [Google Scholar]
- Adam, J.A.; Gulati, S.; Hirsa, A.H.; Bonocora, R.P. Growth of microorganisms in an interfacially driven space bioreactor analog. NPJ Microgravity 2020, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Amalfitano, S.; Levantesi, C.; Copetti, D.; Stefani, F.; Locantore, I.; Guarnieri, V.; Lobascio, C.; Bersani, F.; Giacosa, D.; Detsis, E.; et al. Water and microbial monitoring technologies towards the near future space exploration. Water Res. 2020, 115787. [Google Scholar] [CrossRef]
- Cao, Z.; Liu, H.; Zhao, B.; Pang, Q.; Zhang, X. Extreme environmental stress-induced biological responses in the planarian. BioMed Res. Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, F.; Sueur, C.; Bergouignan, A. Human adaptation to deep space environment: An evolutionary perspective of the foreseen interplanetary exploration. Front. Public Health 2020, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Richter, P.; Vadakedath, N.; Mancinelli, R.; Krüger, M.; Strauch, S.M.; Grimm, D.; Darriet, P.; Chapel, J.P.; Cohen, J.; et al. Exploration of space to achieve scientific breakthroughs. Biotechnol. Adv. 2020, 43, 107572. [Google Scholar] [CrossRef] [PubMed]
- Mahnert, A.; Verseux, C.; Schwendner, P.; Koskinen, K.; Kumpitsch, C.; Blohs, M.; Wink, L.; Brunner, D.; Goessler, T.; Billi, D.; et al. Microbiome dynamics during the HI-SEAS IV mission, and implications for future crewed missions beyond Earth. Microbiome 2021, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Shibuya, M.; Kinoshita, I.; Yatabe, J.; Narumi, I.; Shibata, H.; Hayashi, R.; Fujiwara, D.; Murano, Y.; Hashimoto, H.; et al. DNA Damage and Survival Time Course of Deinococcal Cell Pellets During 3 Years of Exposure to Outer Space. Front. Microbiol. 2020, 11, 2050. [Google Scholar] [CrossRef]
- Afshinnekoo, E.; Scott, R.T.; MacKay, M.J.; Pariset, E.; Cekanaviciute, E.; Barker, R.; Gilroy, S.; Hassane, D.; Smith, S.M.; Zwart, S.R.; et al. Fundamental Biological Features of Spaceflight: Advancing the Field to Enable Deep-Space Exploration. Cell 2020, 183, 1162–1184. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, S.; Wood, J.M.; Sielaff, A.C.; Mora, M.; Duller, S.; Singh, N.K.; Karouia, F.; Moissl-Eichinger, C.; Venkateswaran, K. Assessing the risk of transfer of microorganisms at the international space station due to cargo delivery by commercial resupply vehicles. Front. Microbiol. 2020, 11, 2729. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Ott, C.M.; Pierson, D.L. Changes in monocyte functions of astronauts. Brain Behav. Immun. 2005, 19, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Effect of microgravity & space radiation on microbes. Future Microbiol. 2018, 13, 831–847. [Google Scholar] [PubMed]
- DasSarma, P.; Antunes, A.; Simões, M.F.; DasSarma, S. Earth’s Stratosphere and Microbial Life. Curr. Issues Mol. Biol. 2020, 38, 197–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alekhova, T.A.; Zakharchuk, L.M.; Tatarinova, N.Y. Diversity of bacteria of the genus Bacillus on board of international space station. Dokl. Biochem. Biophys. 2015, 465, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Blachowicz, A.; Chiang, A.J.; Romsdahl, J.; Kalkum, M.; Wang, C.C.; Venkateswaran, K. Proteomic characterization of Aspergillus fumigatus isolated from air and surfaces of the International Space Station. Fungal Genet. Biol. 2019, 124, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Sielaff, A.C.; Urbaniak, C.; Mohan, G.B.M.; Stepanov, V.G.; Tran, Q.; Wood, J.M.; Minich, J.; McDonald, D.; Mayer, T.; Knight, R.; et al. Characterization of the total and viable bacterial and fungal communities associated with the International Space Station surfaces. Microbiome 2019, 7, 1–21. [Google Scholar]
- Gilbert, R.; Torres, M.; Clemens, R.; Hateley, S.; Hosamani, R.; Wade, W.; Bhattacharya, S. Spaceflight and simulated microgravity conditions increase virulence of Serratia marcescens in the Drosophila melanogaster infection model. NPJ Microgravity 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Romsdahl, J.; Blachowicz, A.; Chiang, Y.M.; Venkateswaran, K.; Wang, C.C. Metabolomic analysis of Aspergillus niger isolated from the international space station reveals enhanced production levels of the antioxidant pyranonigrin A. Front. Microbiol. 2020, 11, 931. [Google Scholar] [CrossRef]
- Horneck, G.; Rettberg, P.; Reitz, G.; Panitz, C.; Rabbow, E. Studies on microorganisms in space: A contribution to the discussion on panspermia, search for life on Mars and interaction of life with its environment. Proc. First Eur. Workshop Exo-/Astro Biol. 2001, 496, 105–112. [Google Scholar]
- Rettberg, P.; Eschweiler, U.; Strauch, K.; Reitz, G.; Horneck, G.; Wänke, H.; Brack, A.; Barbier, B. Survival of microorganisms in space protected by meteorite material: Results of the experiment ‘EXOBIOLOGIE’ of the PERSEUS mission. Adv. Space Res. 2002, 30, 1539–1545. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Yang, Y.; Kawashiri, N.; Shiraishi, K.; Takasu, M.; Narumi, I.; Satoh, K.; Hashimoto, H.; Nakagawa, K.; Tanigawa, Y.; et al. The possible interplanetary transfer of microbes: Assessing the viability of Deinococcus spp. under the ISS environmental conditions for performing exposure experiments of microbes in the Tanpopo mission. Orig. Life Evol. Biosph. 2013, 43, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Kozakiewicz, C.P.; Burridge, C.P.; Funk, W.C.; VandeWoude, S.; Craft, M.E.; Crooks, K.R.; Ernest, H.B.; Fountain-Jones, N.M.; Carver, S. Pathogens in space: Advancing understanding of pathogen dynamics and disease ecology through landscape genetics. Evol. Appl. 2018, 11, 1763–1778. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; DiRuggiero, J. Endolithic microbial communities as model systems for ecology and astrobiology. In Model Ecosystems in Extreme Environments—A Volume in Astrobiology Exploring Life on Earth and Beyond, 2nd ed.; Seckbach, J., Rampelotto, P., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 145–168. [Google Scholar]
- Méndez, A.; Rivera-Valentín, E.G.; Schulze-Makuch, D.; Filiberto, J.; Ramírez, R.; Wood, T.E.; Dávila, A.; McKay, C.; Ceballos, K.O.; Jusino-Maldonado, M.; et al. Habitability Models for Astrobiology. Astrobiology 2021, in press. [Google Scholar]
- Epps, C.W.; Keyghobadi, N. Landscape genetics in a changing world: Disentangling historical and contemporary influences and inferring change. Mol. Ecol. 2015, 24, 6021–6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterman, W.E.; Pope, N.S. The use and misuse of regression models in landscape genetic analyses. Mol. Ecol. 2021, 30, 37–47. [Google Scholar] [CrossRef]
- Shirk, A.J.; Landguth, E.L.; Cushman, S.A. The effect of gene flow from unsampled demes in landscape genetic analysis. Mol. Ecol. Resour. 2021, 21, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Preston, L.J.; Dartnell, L.R. Planetary habitability: Lessons learned from terrestrial analogues. Int. J. Astrobiol. 2014, 13, 81–98. [Google Scholar] [CrossRef] [Green Version]
- Martins, Z.; Cottin, H.; Kotler, J.M.; Carrasco, N.; Cockell, C.S.; de la Torre Noetzel, R.; Demets, R.; De Vera, J.P.; d’Hendecourt, L.; Ehrenfreund, P.; et al. Earth as a tool for astrobiology—A European perspective. Space Sci. Rev. 2017, 209, 43–81. [Google Scholar] [CrossRef]
- Antunes, A.; Olsson-Francis, K.; McGenity, T.J. Exploring deep-sea brines as potential terrestrial analogues of oceans in the icy moons of the outer solar system. Curr. Issues Mol. Biol. 2020, 38, 123–162. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Holt, J.; Campbell, J.; Groseman, H.; Josset, J.L.; Bontognali, T.R.; Phelps, A.; Hakobyan, L.; Kuretn, L.; Beattie, A.; et al. Subsurface scientific exploration of extraterrestrial environments (MINAR 5): Analogue science, technology and education in the Boulby Mine, UK. Int. J. Astrobiol. 2019, 18, 157–182. [Google Scholar] [CrossRef] [Green Version]
- Cortesão, M.; Schütze, T.; Marx, R.; Moeller, R.; Meyer, V. Fungal Biotechnology in Space: Why and How? In Grand Challenges in Fungal Biotechnology; Grand Challenges in Biology and Biotechnology; Nevalainen, H., Ed.; Springer: Cham, Switzerland, 2020; pp. 501–535. [Google Scholar]
- Radtke, A.L.; Herbst-Kralovetz, M.M. Culturing and applications of rotating wall vessel bioreactor derived 3D epithelial cell models. J. Vis. Exp. 2012, 62, e3868. [Google Scholar] [CrossRef] [Green Version]
- Ott, C.M.; Crabbé, A.; Wilson, J.W.; Barrila, J.; Castro-Wallace, S.L.; Nickerson, C.A. Microbial Stress: Spaceflight-Induced Alterations in Microbial Virulence and Infectious Disease Risks for the Crew. In Stress Challenges and Immunity in Space; Choukèr, A., Ed.; Springer: Cham, Switzerland, 2020; pp. 327–355. [Google Scholar]
- Mastroleo, F.; Van Houdt, R.; Leroy, B.; Benotmane, M.A.; Janssen, A.; Mergeay, M.; Vanhavere, F.; Hendrickx, L.; Wattiez, R.; Leys, N. Experimental design and environmental parameters affect Rhodospirillum rubrum S1H response to space flight. ISME J. 2009, 3, 1402–1419. [Google Scholar] [CrossRef] [Green Version]
- Villa, A.; Versari, S.; Maier, J.A.M.; Bradamante, S. Cell behavior in simulated microgravity: A comparison of results obtained with RWV and RPM. Gravit. Space Res. 2007, 18, 89–90. [Google Scholar]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Growth of Lactobacillus reuteri DSM17938 Under Two Simulated Microgravity Systems: Changes in Reuterin Production, Gastrointestinal Passage Resistance, and Stress Genes Expression Response. Astrobiology 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Taubner, R.S.; Olsson-Francis, K.; Vance, S.D.; Ramkissoon, N.K.; Postberg, F.; de Vera, J.P.; Antunes, A.; Casas, E.C.; Sekine, Y.; Noack, L.; et al. Experimental and simulation efforts in the astrobiological exploration of exooceans. Space Sci. Rev. 2020, 216, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson-Francis, K.; Ramkissoon, N.K.; Macey, M.C.; Pearson, V.K.; Schwenzer, S.P.; Johnson, D.N. Simulating microbial processes in extraterrestrial, aqueous environments. J. Microbiol. Methods 2020, 172, 105883. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Cockell, C.S. PELS (Planetary Environmental Liquid Simulator): A new type of simulation facility to study extraterrestrial aqueous environments. Astrobiology 2015, 15, 111–118. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Mancinelli, R.L.; Kern, R.G.; Rothschild, L.J.; McKay, C.P. Survival of endospores of Bacillus subtilis on spacecraft surfaces under simulated Martian environments: Implications for the forward contamination of Mars. Icarus 2003, 165, 253–276. [Google Scholar] [CrossRef]
- Berry, B.J.; Jenkins, D.G.; Schuerger, A.C. Effects of simulated Mars conditions on the survival and growth of Escherichia coli and Serratia liquefaciens. Appl. Environ. Microbiol. 2010, 76, 2377–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, K.M.; Britt, D.T.; Smith, T.M.; Fritsche, R.F.; Batcheldor, D. Mars global simulant MGS-1: A Rocknest-based open standard for basaltic Martian regolith simulants. Icarus 2019, 317, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Ramkissoon, N.K.; Pearson, V.K.; Schwenzer, S.P.; Schröder, C.; Kirnbauer, T.; Wood, D.; Seidel, R.G.; Miller, M.A.; Olsson-Francis, K. New simulants for Martian regolith: Controlling iron variability. Planet. Space Sci. 2019, 179, 104722. [Google Scholar] [CrossRef]
- Rettberg, P.; Rabbow, E.; Panitz, C.; Horneck, G. Biological space experiments for the simulation of Martian conditions: UV radiation and Martian soil analogues. Adv. Space Res. 2004, 33, 1294–1301. [Google Scholar] [CrossRef]
- Polgári, M.; Gyollai, I.; Bérczi, S. Microbially mediated transformation inside the Kaba meteorite? Acta Geosci. Debr. 2018, 1, 55–69. [Google Scholar]
- Perrone, G.; Ferrara, M.; Medina, A.; Pascale, M.; Magan, N. Toxigenic fungi and mycotoxins in a climate change scenario: Ecology, genomics, distribution, prediction and prevention of the risk. Microorganisms 2020, 8, 1496. [Google Scholar] [CrossRef]
- Váry, Z.; Mullins, E.; McElwain, J.C.; Doohan, F.M. The severity of wheat diseases increases when plants and pathogens are acclimatized to elevated carbon dioxide. Glob. Chang. Biol. 2015, 21, 2661–2669. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Zhou, S.; Chen, L.; Xu, B. Impact of climate change on human infectious diseases: Empirical evidence and human adaptation. Environ. Int. 2016, 86, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Epstein, P.R.; Burke, T.A.; Balbus, J.M. Global climate change and emerging infectious diseases. JAMA 1996, 275, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Boodram, L.L.G.; Paz, S.; Ibarra, A.M.S.; Wasserheit, J.N.; Lowe, R. Strengthening the global response to climate change and infectious disease threats. BMJ 2020, 371, m3081. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Gibson, A. OECD Space Forum, Directorate for Science, Technology and Innovation. 2014. Available online: www.oecd.org/futures/space-technologies-and-climate-change.pdf (accessed on 1 March 2021).
- Novikova, N.; Deshevaya, E.; Levinskikh, M.; Polikarpov, N.; Poddubko, S.; Gusev, O.; Sychev, V. Study of the effects of the outer space environment on dormant forms of microorganisms, fungi and plants in the ‘Expose-R’ experiment. Int. J. Astrobiol. 2015, 14, 137–142. [Google Scholar] [CrossRef]
- Alarico, S.; Nunes-Costa, D.; Silva, A.; Costa, M.; Macedo-Ribeiro, S.; Empadinhas, N. A genuine mycobacterial thermophile: Mycobacterium hassiacum growth, survival and GpgS stability at near-pasteurization temperatures. Microbiology 2020, 166, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Thirsk, R.; Kuipers, A.; Mukai, C.; Williams, D. The space-flight environment: The International Space Station and beyond. Can. Med Assoc. J. 2009, 180, 1216–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo-Cavazos, P.; Nicholson, W.L. Mechanotransduction in Prokaryotes: A Possible Mechanism of Spaceflight Adaptation. Life 2021, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Gilman, R.T.; Nuismer, S.L.; Jhwueng, D.C. Coevolution in multidimensional trait space favours escape from parasites and pathogens. Nature 2012, 483, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Yao, L.; Zhu, J.; Shi, J.; Jin, M.; Huang, Q.; Yang, H. Simulated microgravity affects some biological characteristics of Lactobacillus acidophilus. Appl. Microbiol. Biotechnol. 2017, 101, 3439–3449. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, A.; Cleenwerck, I.; Iversen, C.; De Vos, P.; Stephan, R. Pantoea gaviniae sp. nov. and Pantoea calida sp. nov., isolated from infant formula and an infant formula production environment. Int. J. Syst. Evol. Microbiol. 2010, 60, 2786–2792. [Google Scholar] [CrossRef]
- Brady, C.L.; Cleenwerck, I.; Venter, S.N.; Engelbeen, K.; De Vos, P.; Coutinho, T.A. Emended description of the genus Pantoea, description of four species from human clinical samples, Pantoea septica sp. nov., Pantoea eucrina sp. nov., Pantoea brenneri sp. nov. and Pantoea conspicua sp. nov., and transfer of Pectobacterium cypripedii (Hori 1911) Brenner et al. 1973 emend. Hauben et al. 1998 to the genus as Pantoea cypripedii comb. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 2430–2440. [Google Scholar] [PubMed] [Green Version]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef] [Green Version]
- Solomon, S.A.; Bharadwaj, A.R.; Singh, N.K.; Wood, J.M.; Debieu, M.; O’Hara, N.B.; Mason, C.E.; Venkateswaran, K. Draft Genome Sequences of Klebsiella Species Isolated from the International Space Station. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, A.; Crombé, F.; Bosmans, P.; Cools, F.; Piérard, D.; Wybo, I. Serratia marcescens outbreak in a neonatal intensive care unit and the potential of whole-genome sequencing. J. Hosp. Infect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Géry, A.; Mouet, A.; Gravey, F.; Fines-Guyon, M.; Guerin, F.; Ethuin, F.; Borgey, F.; Lubrano, J.; Le Hello, S. Investigation of Serratia marcescens surgical site infection outbreak associated with peroperative ultrasonography probe. J. Hosp. Infect. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Grebennikova, T.V.; Syroeshkin, A.V.; Shubralova, E.V.; Eliseeva, O.V.; Kostina, L.V.; Kulikova, N.Y.; Latyshev, O.E.; Morozova, M.A.; Yuzhakov, A.G.; Zlatskiy, I.A.; et al. The DNA of bacteria of the World Ocean and the Earth in cosmic dust at the International Space Station. Sci. World J. 2018, 2018, 7360147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, M.F.; Ottoni, C.A.; Antunes, A. Mycogenic metal nanoparticles for the treatment of mycobacterioses. Antibiotics 2020, 9, 569. [Google Scholar] [CrossRef]
- Klintworth, R.; Reher, H.J.; Viktorov, A.N.; Bohle, D. Biological induced corrosion of materials II: New test methods and experiences from MIR station. Acta Astronaut. 1999, 44, 569–578. [Google Scholar] [CrossRef]
- Deinhardt-Emmer, S.; Höring, S.; Mura, C.; Hillemann, D.; Hermann, B.; Sachse, S.; Bohnert, J.; Löffler, B. First time isolation of Mycobacterium hassiacum from a respiratory sample. Clin. Med. Insights Circ. Respir. Pulm. Med. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Abshire, C.F.; Prasai, K.; Soto, I.; Shi, R.; Concha, M.; Baddoo, M.; Flemington, E.K.; Ennis, D.G.; Scott, R.S.; Harrison, L. Exposure of Mycobacterium marinum to low-shear modeled microgravity: Effect on growth, the transcriptome and survival under stress. NPJ Microgravity 2016, 2, 1–14. [Google Scholar] [CrossRef]
- Kovács, Á.T. Bacillus subtilis. Trends Microbiol. 2019, 27, 724–725. [Google Scholar] [CrossRef]
- Aunins, T.R.; Erickson, K.E.; Prasad, N.; Levy, S.E.; Jones, A.; Shrestha, S.; Mastracchio, R.; Stodieck, L.; Klaus, D.; Zea, L.; et al. Spaceflight modifies Escherichia coli gene expression in response to antibiotic exposure and reveals role of oxidative stress response. Front. Microbiol. 2018, 9, 310. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Perlin, T.; Mouriño-Pérez, R.R.; Sánchez-Carbente, M.D.R.; Folch-Mallol, J.L.; Pérez-Llano, Y.; Gunde-Cimerman, N.; Sánchez, N.D.C.; Batista-García, R.A. Haloadaptative Responses of Aspergillus sydowii to extreme water deprivation: Morphology, compatible solutes, and oxidative stress at NaCl saturation. J. Fungi 2020, 6, 316. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Gilbert, M.K.; Mack, B.M.; OBrian, G.R.; Rodriguez, A.; Bhatnagar, D.; Payne, G.; Magan, N. Interactions between water activity and temperature on the Aspergillus flavus transcriptome and aflatoxin B1 production. Int. J. Food Microbiol. 2017, 256, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Where Aspergillosis Comes From. Available online: www.cdc.gov/fungal/diseases/aspergillosis/causes.html (accessed on 22 February 2021).
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Vesper, S.J.; Wong, W.; Kuo, C.M.; Pierson, D.L. Mold species in dust from the International Space Station identified and quantified by mold-specific quantitative PCR. Res. Microbiol. 2008, 159, 432–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, B.P.; Blachowicz, A.; Palmer, J.M.; Romsdahl, J.; Huttenlocher, A.; Wang, C.C.; Keller, N.P.; Venkateswaran, K. Characterization of Aspergillus fumigatus isolates from air and surfaces of the International Space Station. mSphere 2016, 1, e00227-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, A.; Fernández-Molina, J.V.; Bikandi, J.; Ramírez, A.; Margareto, J.; Sendino, J.; Hernando, F.L.; Pontón, J.; Garaizar, J.; Rementeria, A. What makes Aspergillus fumigatus a successful pathogen? Genes and molecules involved in invasive aspergillosis. Rev. Iberoam. Micol. 2010, 27, 155–182. [Google Scholar] [CrossRef] [PubMed]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An emerging pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465717. [Google Scholar] [CrossRef] [Green Version]
- Geltner, C.; Lass-Flörl, C.; Bonatti, H.; Müller, L.; Stelzmüller, I. Invasive pulmonary mycosis due to Penicillium chrysogenum: A new invasive pathogen. Transplantation 2013, 95, e21–e23. [Google Scholar] [CrossRef]
- Person, A.K.; Chudgar, S.M.; Norton, B.L.; Tong, B.C.; Stout, J.E. Aspergillus niger: An unusual cause of invasive pulmonary aspergillosis. J. Med. Microbiol. 2010, 59, 834. [Google Scholar] [CrossRef] [Green Version]
- Bai, F.; Wei, C.; Zhang, L. Analysis of the mold resistance of the non-metal materials on space station and suggestions of improvement measures. In Proceedings of the 14th International Conference on Man-Machine-Environment System Engineering; Long, S., Dhillon, B.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 197–202. [Google Scholar]
- Sobisch, L.Y.; Rogowski, K.M.; Fuchs, J.; Schmieder, W.; Vaishampayan, A.; Oles, P.; Novikova, N.; Grohmann, E. Biofilm forming antibiotic resistant gram-positive pathogens isolated from surfaces on the international space station. Front. Microbiol. 2019, 10, 543. [Google Scholar] [CrossRef]
- Rettberg, P.; Antunes, A.; Brucato, J.; Cabezas, P.; Collins, G.; Haddaji, A.; Kminek, G.; Leuko, S.; McKenna-Lawlor, S.; Moissl-Eichinger, C.; et al. Biological contamination prevention for outer solar system moons of astrobiological interest: What do we need to know? Astrobiology 2019, 19, 951–974. [Google Scholar] [CrossRef] [PubMed]
- De Vera, J.P.; Alawi, M.; Backhaus, T.; Baqué, M.; Billi, D.; Böttger, U.; Berger, T.; Bohmeier, M.; Cockell, C.; Demets, R.; et al. Limits of life and the habitability of Mars: The ESA space experiment BIOMEX on the ISS. Astrobiology 2019, 19, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuko, S.; Rettberg, P.; Pontifex, A.L.; Burns, B.P. On the response of halophilic archaea to space conditions. Life 2014, 4, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuko, S.; Domingos, C.; Parpart, A.; Reitz, G.; Rettberg, P. The survival and resistance of Halobacterium salinarum NRC-1, Halococcus hamelinensis, and Halococcus morrhuae to simulated outer space solar radiation. Astrobiology 2015, 15, 987–997. [Google Scholar] [CrossRef]
- Dornmayr-Pfaffenhuemer, M.; Legat, A.; Schwimbersky, K.; Fendrihan, S.; Stan-Lotter, H. Responses of haloarchaea to simulated microgravity. Astrobiology 2011, 11, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moissl-Eichinger, C.; Pausan, M.; Taffner, J.; Berg, G.; Bang, C.; Schmitz, R.A. Archaea are interactive components of complex microbiomes. Trends Microbiol. 2018, 26, 70–85. [Google Scholar] [CrossRef]
- Griffin, D.W. The quest for extraterrestrial life: What about the viruses? Astrobiology 2013, 13, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Berliner, A.J.; Mochizuki, T.; Stedman, K.M. Astrovirology: Viruses at large in the universe. Astrobiology 2018, 18, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bérces, A.; Egyeki, M.; Fekete, A.; Horneck, G.; Kovács, G.; Panitz, C.; Rontó, G. The PUR experiment on the EXPOSE-R facility: Biological dosimetry of solar extraterrestrial UV radiation. Int. J. Astrobiol. 2015, 14, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Lindeboom, R.E.F.; Ilgrande, C.; Carvajal-Arroyo, J.M.; Coninx, I.; Van Hoey, O.; Roume, H.; Morozova, J.; Udert, K.M.; Sas, B.; Paille, C.; et al. Nitrogen cycle microorganisms can be reactivated after Space exposure. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Zammuto, V.; Rizzo, M.G.; De Plano, L.M.; Franco, D.; Guglielmino, S.; Caccamo, M.T.; Magazù, S.; Fujimori, A.; Giudice, A.L.; Guglielmin, M.; et al. Effects of heavy ion particle irradiation on spore germination of Bacillus spp. from extremely hot and cold environments. Life 2020, 10, 264. [Google Scholar] [CrossRef]
- Santomartino, R.; Waajen, A.C.; De Wit, W.; Nicholson, N.; Parmitano, L.; Loudon, C.M.; Moeller, R.; Rettberg, P.; Fuchs, F.M.; Van Houdt, R.; et al. No effect of microgravity and simulated Mars gravity on final bacterial cell concentrations on the International Space Station: Applications to space bioproduction. Front. Microbiol. 2020, 11, 579156. [Google Scholar] [CrossRef]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. Evaluation of acquired antibiotic resistance in Escherichia coli exposed to long-term low-shear modeled microgravity and background antibiotic exposure. Mbio 2019, 10, e02637-18. [Google Scholar] [CrossRef] [Green Version]
- Lam, K.S.; Mamber, S.W.; Pack, E.J.; Forenza, S.; Fernandes, P.B.; Klaus, D.M. The effects of space flight on the production of monorden by Humicola fuscoatra WC5157 in solid-state fermentation. Appl. Microbiol. Biotechnol. 1998, 49, 579–583. [Google Scholar] [CrossRef]
- Guo, Y.; Li, Y.; Su, L.; Chang, D.; Liu, W.; Wang, T.; Yuan, Y.; Fang, X.; Wang, J.; Li, T.; et al. Comparative genomic analysis of Klebsiella pneumonia (LCT-KP214) and a mutant strain (LCT-KP289) obtained after spaceflight. BMC Genom. 2014, 15, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, P.W.; Leff, L. The effect of simulated microgravity on bacteria from the Mir space station. Microgravity Sci. Technol. 2004, 15, 35–41. [Google Scholar] [CrossRef]
- Mastroleo, F.; Van Houdt, R.; Atkinson, S.; Mergeay, M.; Hendrickx, L.; Wattiez, R.; Leys, N. Modelled microgravity cultivation modulates N-acylhomoserine lactone production in Rhodospirillum rubrum S1H independently of cell density. Microbiology 2013, 159, 2456–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, P.; Zhang, B.; Zhao, X.; Li, D.; Yu, Y.; Zhang, X.; Huang, B.; Liu, C. Decreased metabolism and increased tolerance to extreme environments in Staphylococcus warneri during long-term spaceflight. MicrobiologyOpen 2019, 8, e917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Liu, N.; Rong, X.; Ruan, J.; Huang, Y. Effects of simulated microgravity and spaceflight on morphological differentiation and secondary metabolism of Streptomyces coelicolor A3. Appl. Microb. Cell Physiol. 2015, 99, 4409–4422. [Google Scholar] [CrossRef]
- Lam, K.S.; Gustavson, D.R.; Pirnik, D.L.; Pack, E.; Bulanhagui, C.; Mamber, S.W.; Forenza, S.; Stodieck, L.S.; Klaus, D.M. The effect of space flight on the production of actinomycin D by Streptomyces plicatus. J. Ind. Microbiol. Biotechnol. 2002, 29, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Benoit, M.R.; Li, W.; Stodieck, L.S.; Lam, K.S.; Winther, C.L.; Roane, T.M.; Klaus, D.M. Microbial antibiotic production aboard the International Space Station. Appl. Microbiol. Biotechnol. 2006, 70, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Ryba-White, M.; Nedukha, O.; Hilaire, E.; Guikema, J.A.; Kordyum, E.; Leach, J.E. Growth in microgravity increases susceptibility of soybean to a fungal pathogen. Plant Cell Physiol. 2001, 42, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craven, E.; Winters, M.; Smith, A.L.; Lalime, E.; Mancinelli, R.; Shirey, B.; Schubert, W.; Schuerger, A.; Burgin, M.; Seto, E.P.; et al. Biological safety in the context of backward planetary protection and Mars Sample Return: Conclusions from the Sterilization Working Group. Int. J. Astrobiol. 2021, 20, 1–28. [Google Scholar] [CrossRef]
- Raguse, M.; Fiebrandt, M.; Denis, B.; Stapelmann, K.; Eichenberger, P.; Driks, A.; Eaton, P.; Awakowicz, P.; Moeller, R. Understanding of the importance of the spore coat structure and pigmentation in the Bacillus subtilis spore resistance to low-pressure plasma sterilization. J. Phys. D Appl. Phys. 2016, 49, 285401. [Google Scholar] [CrossRef]
- Hicks, P.M.; Nelson, J. Preliminary Testing of Electrolytic Silver Ion Generation for Spacecraft Potable Water Systems. Int. Conf. Environ. Syst. 2020, 371, 1–10. [Google Scholar]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? NPJ Microgravity 2020, 6, 1–7. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Eleveld, M.; op den Camp, H.J.; van der Meer, J.W.; Gow, N.A.; de Jonge, M.I. Immune recognition of putative alien microbial structures: Host-pathogen interactions in the age of space travel. PLoS Pathog. 2020, 16, e1008153. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, C.; Lu, Z.; Ge, P.; Cui, Y.; Zhao, D.; Yang, X.; Wu, B.; Qiang, L.; Zhang, Y.; et al. Simulated microgravity suppresses MAPK pathway-mediated innate immune response to bacterial infection and induces gut microbiota dysbiosis. FASEB J. 2020, 34, 14631–14644. [Google Scholar] [CrossRef]
- Chen, H.; Lv, K.; Ji, G.; Yuan, Y.; Lu, L.; Liang, F.; Li, K.; Xu, Z.; Xiong, J.; Qu, L.; et al. Physiological acclimatization of the liver to 180-day isolation and the Mars solar day. BioMed Res. Int. 2020, 2020, 2796510. [Google Scholar] [CrossRef]
- Mermel, L.A. Infection prevention and control during prolonged human space travel. Clin. Infect. Dis. 2013, 56, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Lauber, B.A.; Bolshakova, O.; Ullrich, O. Spacecraft microbiology. In The Immune System in Space: Are We Prepared? Choukèr, A., Ullrich, O., Eds.; Springer: Cham, Switzerland, 2016; pp. 77–86. [Google Scholar]
- Green, M.J.; Aylott, J.W.; Williams, P.; Ghaemmaghami, A.M.; Williams, P.M. Immunity in Space: Prokaryote Adaptations and Immune Response in Microgravity. Life 2021, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Nie, L.; Xie, S.; Li, M.; Zhu, C.; Qiu, X.; Kuang, J.; Liu, C.; Lu, C.; Li, W.; et al. Attenuation of Antiviral Immune Response Caused by Perturbation of TRIM25-Mediated RIG-I Activation under Simulated Microgravity. Cell Rep. 2021, 34, 108600. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Babiak-Vazquez, A.; Johnston, S.; Pierson, D.L.; Ott, C.M.; Sams, C. Incidence of clinical symptoms during long-duration orbital spaceflight. Int. J. Gen. Med. 2016, 9, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crucian, B.E.; Choukèr, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune system dysregulation during spaceflight: Potential countermeasures for deep space exploration missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Meerman, M.; Bracco Gartner, T.C.; Buikema, J.W.; Wu, S.M.; Siddiqi, S.; Bouten, C.V.; Grande-Allen, K.J.; Suyker, W.J.; Hjortnaes, J. Myocardial Disease and Long-Distance Space Travel: Solving the Radiation Problem. Front. Cardiovasc. Med. 2021, 8, 631985. [Google Scholar] [CrossRef]
- Josh, R.; Sensinger, M.; Veluri, M. The effect of microgravity on cytokine production in response to antigens. BioTreks 2020, 5, e202009. [Google Scholar]
- Shi, L.; Tian, H.; Wang, P.; Li, L.; Zhang, Z.; Zhang, J.; Zhao, Y. Spaceflight and simulated microgravity suppresses macrophage development via altered RAS/ERK/NFκB and metabolic pathways. Cell. Mol. Immunol. 2020, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the impact of microgravity at the cellular level: Implications for human disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D. Guest Edited Collection: Gravitational biology and space medicine. Sci. Rep. 2019, 9, 14399. [Google Scholar] [CrossRef]
- Basner, M.; Dinges, D.F.; Mollicone, D.J.; Savelev, I.; Ecker, A.J.; Di Antonio, A.; Jones, C.W.; Hyder, E.C.; Kan, K.; Morukov, B.V.; et al. Psychological and behavioral changes during confinement in a 520-day simulated interplanetary mission to Mars. PLoS ONE 2014, 9, e93298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, R.T.; Baker, E.S. Gynecologic and Reproductive Considerations. In Principles of Clinical Medicine for Space Flight; Barratt, M.R., Baker, E., Pool, S.L., Eds.; Springer: New York, NY, USA, 2019; pp. 747–760. [Google Scholar]
- Nickerson, C.A.; Ott, C.M.; Mister, S.J.; Morrow, B.J.; Burns-Keliher, L.; Pierson, D.L. Microgravity as a novel environmental signal affecting Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2000, 68, 3147–3152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matin, A.; Lynch, S.V.; Benoit, M.R. Increased bacterial resistance and virulence in simulated microgravity and its molecular basis. Gravit. Space Biol. 2006, 19, 31–42. [Google Scholar]



{kind=link}
{kind=link}
| Parameter | Alterations and/or Adaptations | Reference |
|---|---|---|
| Solar UV radiation | Increased microbial mutation rates. DNA damage through double/single-strand breaks, base modifications or pyrimidine dimerization. Oxidative stress which induces the production of key enzymes (e.g., catalase). | [8,12] |
| High vacuum | DNA damage through induction of base deletion and insertion. | [8] |
| High-dose ionizing radiation or desiccation | DNA damage through double-strand breaks (DSBs). | [8] |
| Radiation capable of penetrating the International Space Station (ISS) spacecraft | Generation of reactive oxygen species (ROS) within biological systems, with oxidative stress and consequent DNA damage. | [6,18] |
| Microgravity | Increased growth. Smaller lag phase in bacterial growth curves. Shear forces get reduced, there is no sedimentation, diffusion processes get slower, and no convection without gravity. Hindered access to oxygen, metabolites, and nutrients, affecting mass and heat transfer. | [6,12,60] |
| Organism Exposed | Exposure Experiment | Results and Conclusions Obtained | Reference | |
|---|---|---|---|---|
| Location | Time | |||
| Axenic and mixed cultures of microorganisms capable of essential nitrogen cycle conversions | Low Earth Orbit (LEO, 258–571 km of altitude), on a Foton-M4 flight | 44 days |
| [97] |
| Bacillus sp. (two psychrotolerant strains); Bacillus horneckiae, and Bacillus licheniformis (thermophiles) | HIMAC at the NIRS, Japan, with exposure to HZE particles: He and Fe ions, presence of germinants (Glu, Ala, and Val). | 0–8 h |
| [98] |
| Bacillus subtilis, Cupriavidus metallidurans, and Sphingomonas desiccabilis | Simulated Martian gravity (0.38× g), on the ISS | 21 days, plus 2 days-flight (on a Space X Falcon-9 rocket) |
| [99] |
| Deinococcus aerius and Deinococcus radiodurans, in cell pellets | Outside the ISS | 1–3 years |
| [8] |
| Escherichia coli | Simulated microgravity, HARV | Time needed for the growth of 1000 generations |
| [100] |
| Humicola fuscoatra | Space Shuttle mission STS-77 | 10 days |
| [101] |
| Klebsiella pneumonia | Outer space of the Shenzhou VIII spacecraft | 17 days |
| [102] |
| Lactobacillus acidophilus | Simulated microgravity (RWV) | Up to 36 h |
| [60] |
| Lactobacillus reuteri | Simulated microgravity (RWV) | 18 h |
| [37] |
| Lactobacillus reuteri | Simulated microgravity (RPM) | 18 h |
| [37] |
| Ralstonia pickettii and Sphingobacterium thalpophilium (isolated from water systems of the Mir space station) | Simulated microgravity (STLV on a RCCS) | Up to 14 days |
| [103] |
| Rhodospirillum rubrum | Anaerobiose at ISS | 8 days, plus 2 days flight (in the Soyus carrier rocket) |
| [35] |
| Rhodospirillum rubrum | Simulated microgravity (RPM) and space-ionizing radiation, on Earth | 10 days |
| [35] |
| Rhodospirillum rubrum | Simulated partial microgravity (RWV) conditions, on Earth | 10 days |
| [104] |
| Serratia marcescens | Spaceflight conditions at ISS | n.r. |
| [17] |
| Serratia marcescens | Simulated microgravity (RWV) | n.r. |
| [17] |
| Spores of Aspergillus sydowii, Aspergillus versicolor, Penicillium aurantiogriseum, and Penicillium expansum | Outer surface of ISS | Over 22 months |
| [55] |
| Spores of Bacillus licheniformis, Bacillus pumilus, and Bacillus subtilis | Outer surface of ISS | Over 22 months |
| [55] |
| Spores of Bacillus subtilis | Outside MIR space station. | 3 months |
| [20] |
| Staphylococcus warneri | Space environment aboard Tiangong-2 space laboratory. | 64 days, plus 15 days-flight (on the Shenzhou-10 spacecraft) |
| [105] |
| Streptomyces coelicolor | Shenzhou-8 spacecraft, and simulated microgravity (2D-clinostat) on Earth. | 16.5 days |
| [106] |
| Streptomyces plicatus | Space Shuttle mission STS-80 | 7 and 12 days |
| [107] |
| Streptomyces plicatus | ISS | 8, 12, and 72 days |
| [108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, M.F.; Antunes, A. Microbial Pathogenicity in Space. Pathogens 2021, 10, 450. https://doi.org/10.3390/pathogens10040450
Simões MF, Antunes A. Microbial Pathogenicity in Space. Pathogens. 2021; 10(4):450. https://doi.org/10.3390/pathogens10040450
Chicago/Turabian StyleSimões, Marta Filipa, and André Antunes. 2021. "Microbial Pathogenicity in Space" Pathogens 10, no. 4: 450. https://doi.org/10.3390/pathogens10040450
APA StyleSimões, M. F., & Antunes, A. (2021). Microbial Pathogenicity in Space. Pathogens, 10(4), 450. https://doi.org/10.3390/pathogens10040450