
Phenelzine and Amoxapine Inhibit Tyramine and d-Glucuronic Acid Catabolism in Clinically Significant Salmonella in A Serotype-Independent Manner



Abstract
:1. Introduction
2. Results and Discussion
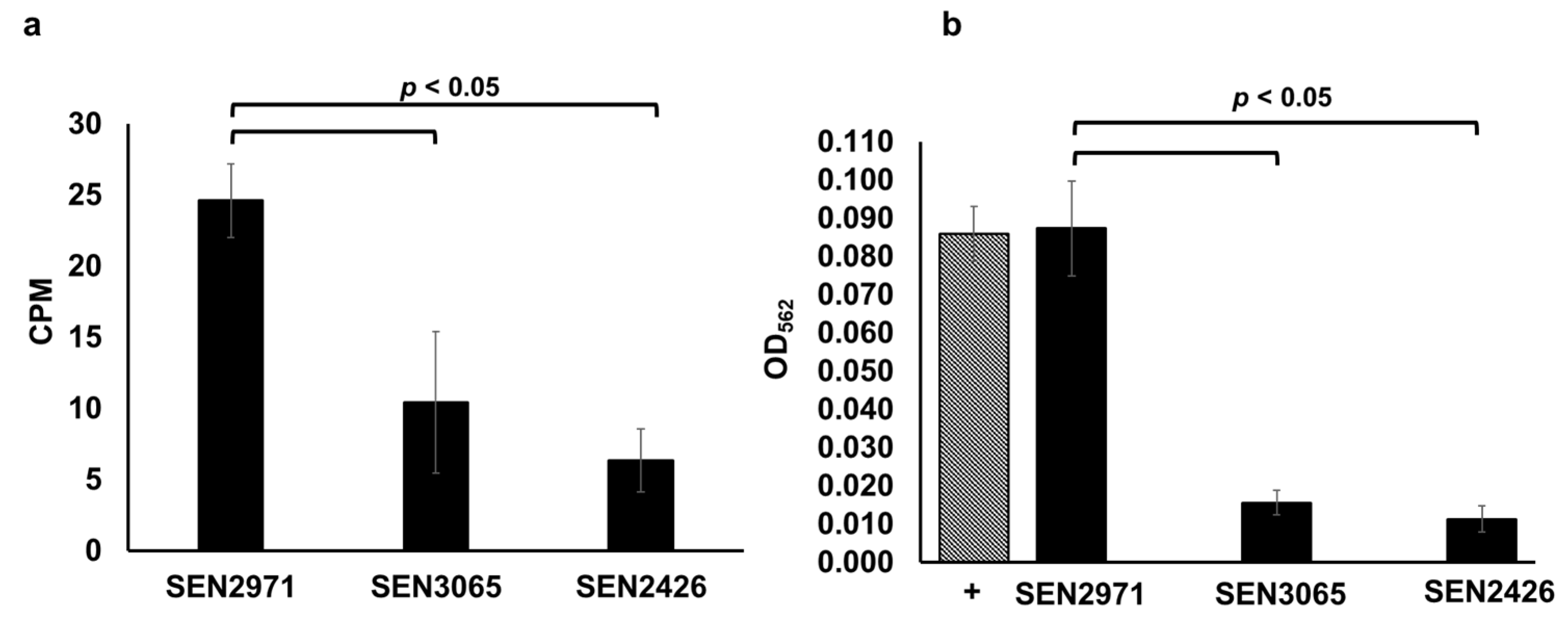
2.1. SEN2971, SEN3065, and SEN2426 Are TYR Oxidoreductases
2.2. Phenelzine Inhibits the TYR Oxidoreductase Activity of Recombinant SEN2971, SEN3065, and SEN2426
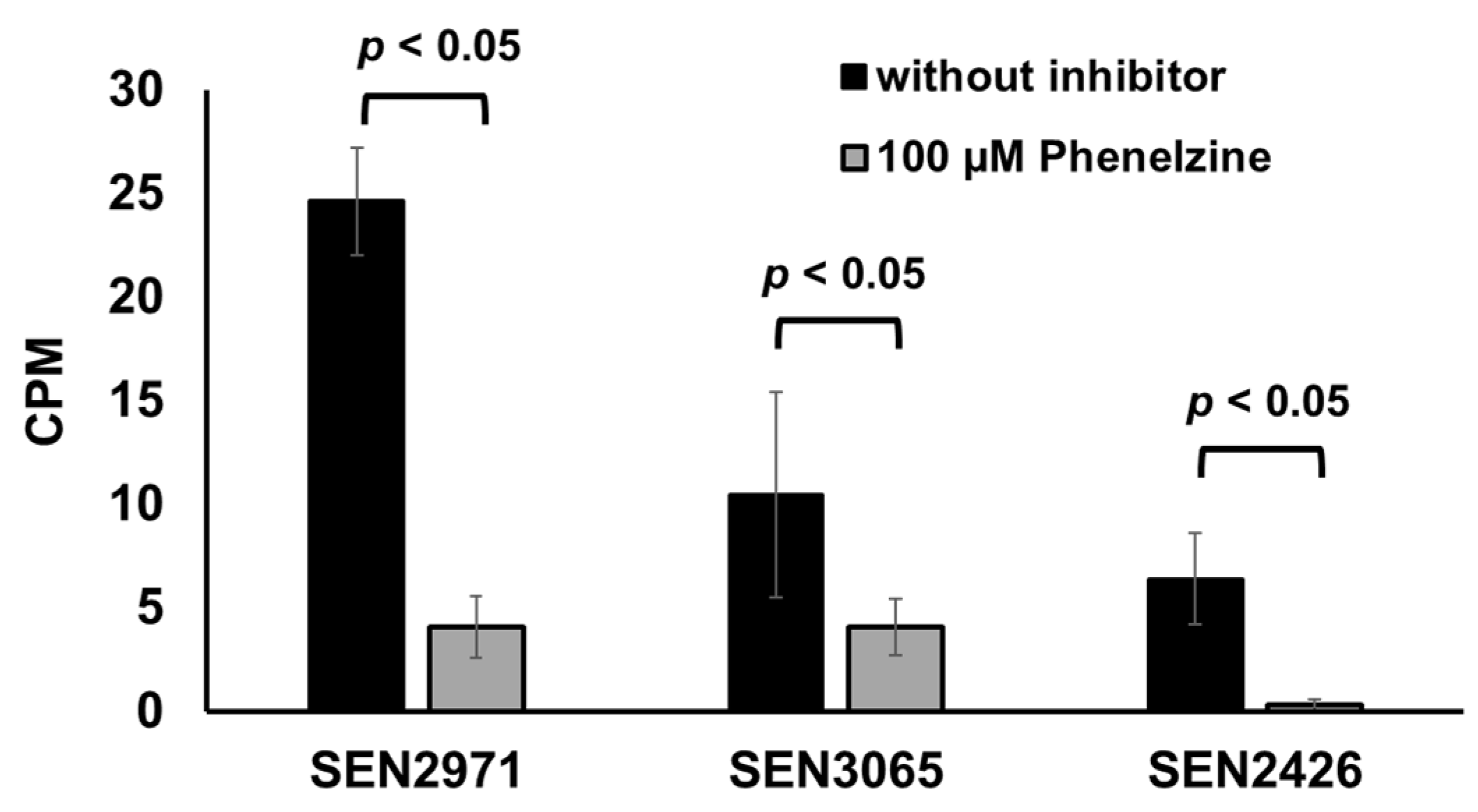
2.3. Phenelzine Inhibits the Growth of S. Enteritidis in the Presence of TYR as the Sole Energy Source
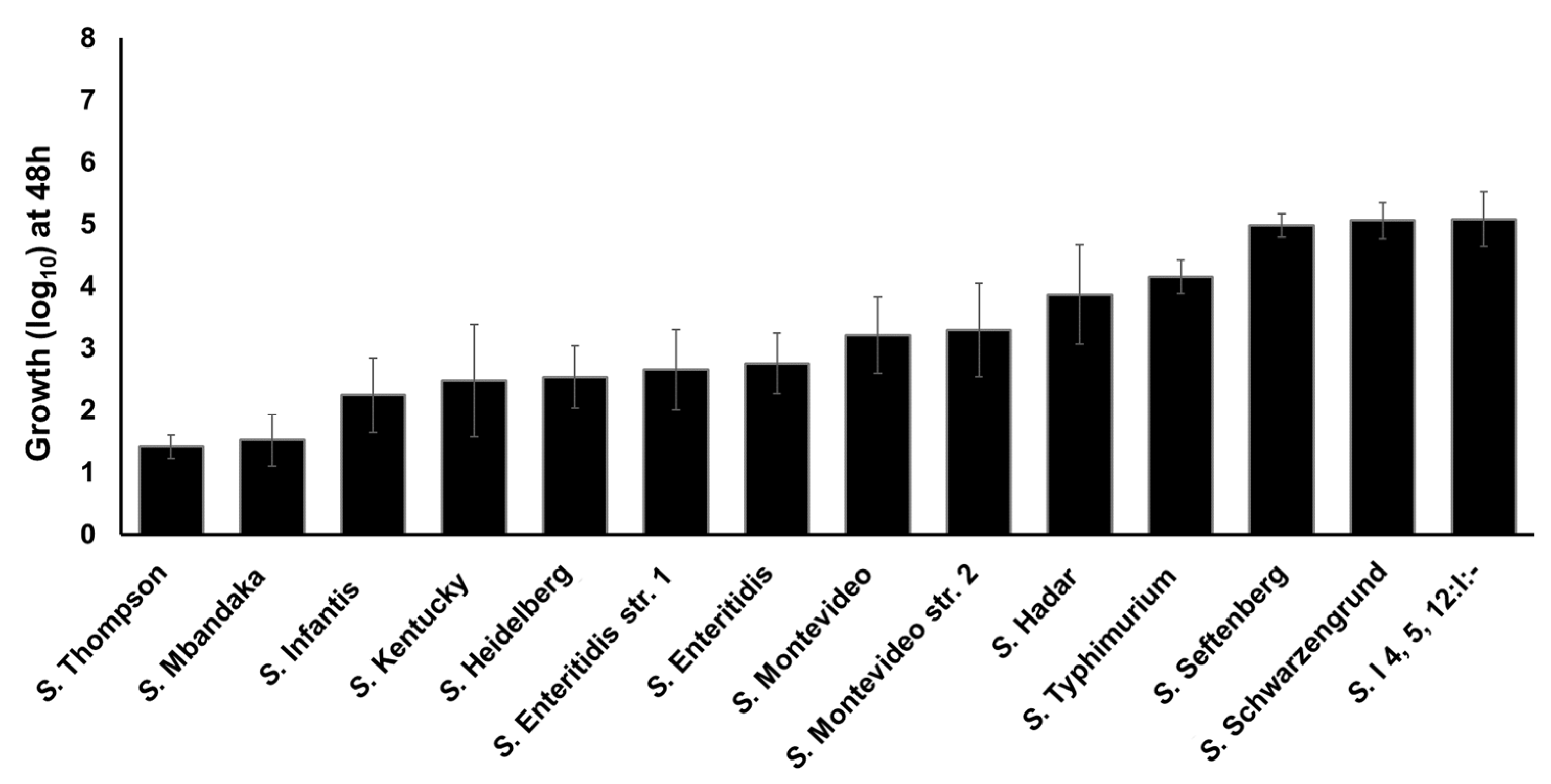
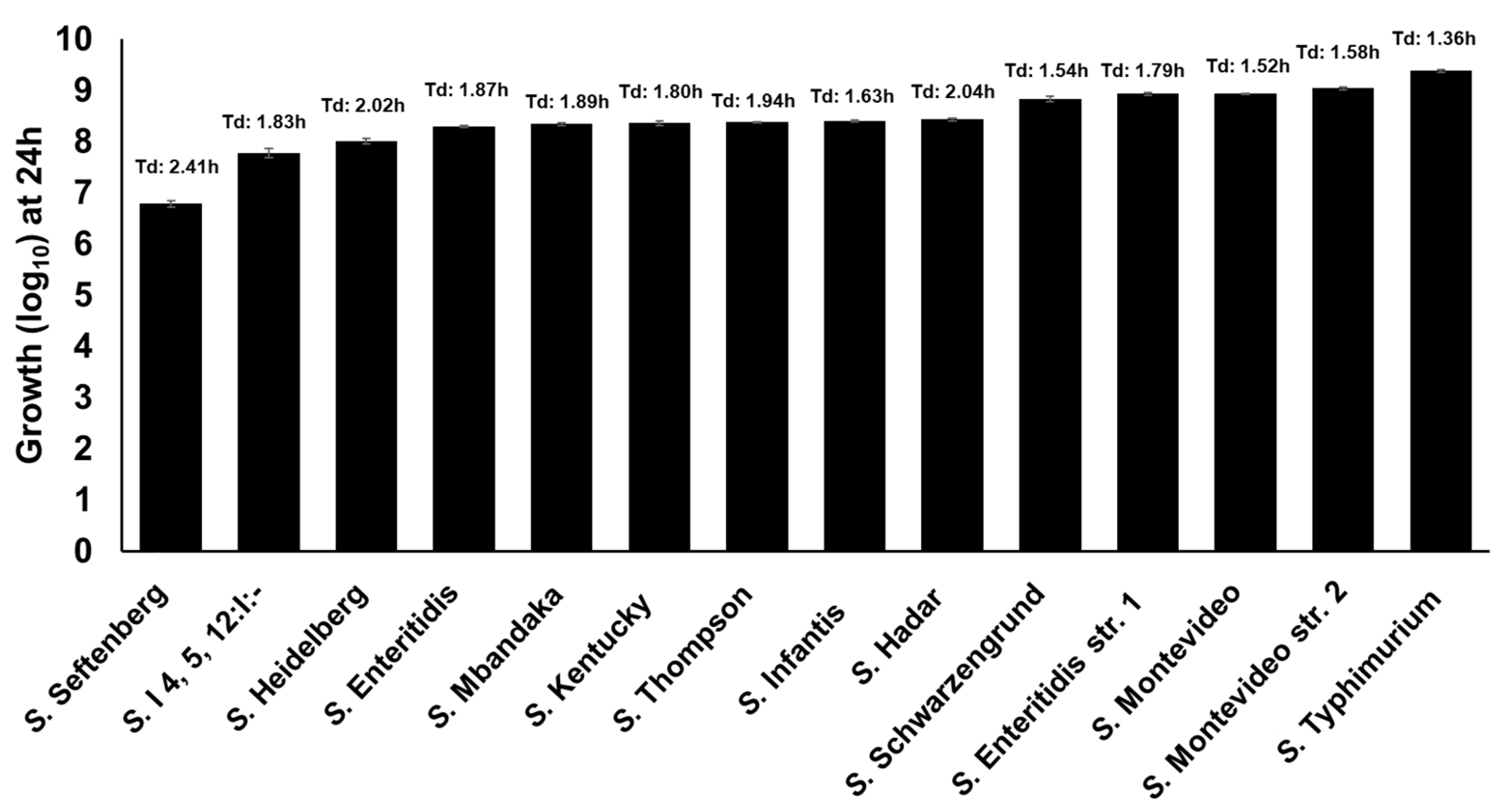
2.4. Clinically Significant NTS Utilize TYR as a Source of Energy
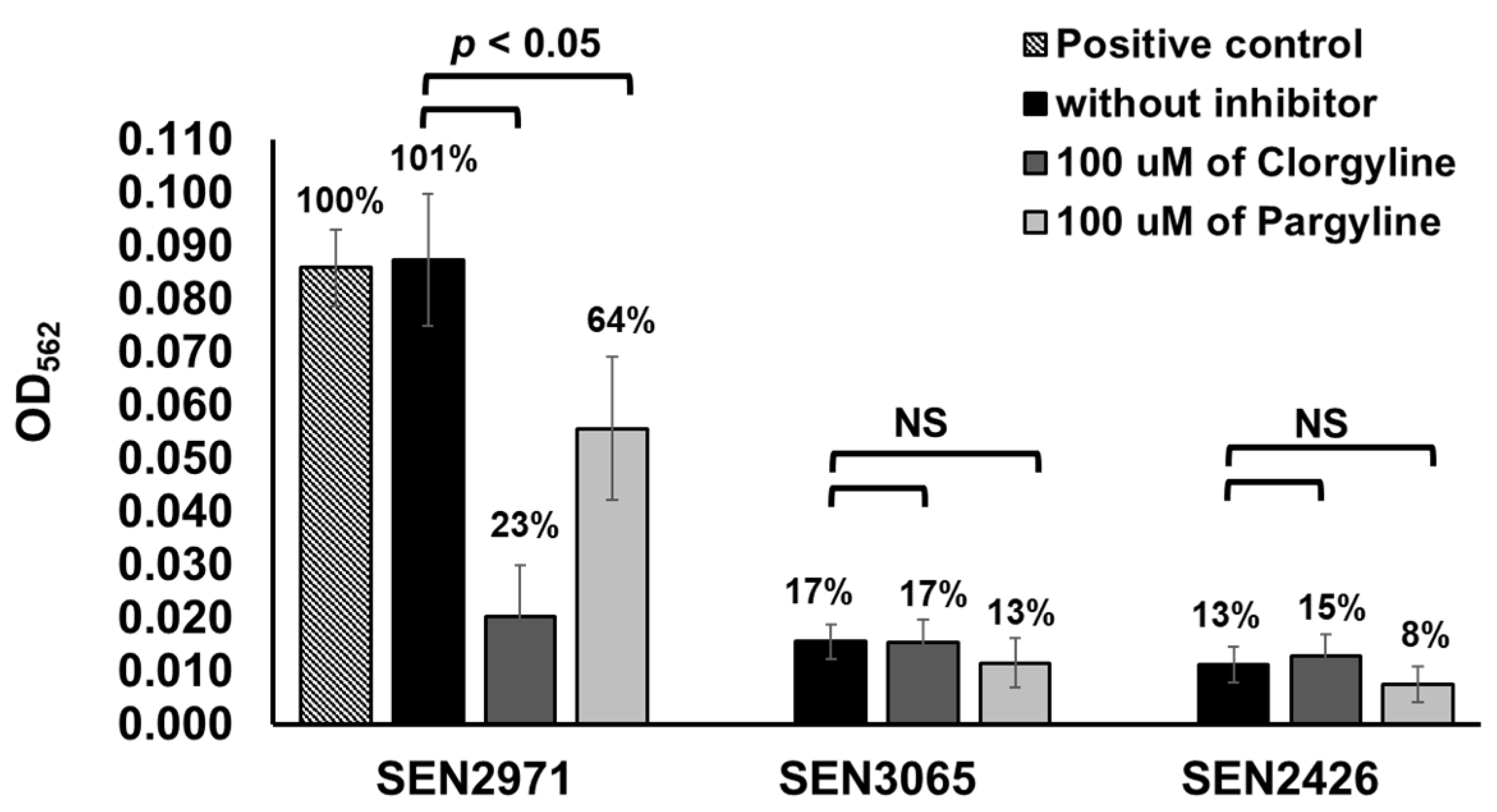
2.5. Phenelzine Inhibits the Ability of NTS to Utilize TYR as a Sole Energy Source in a Serotype-Independent Manner
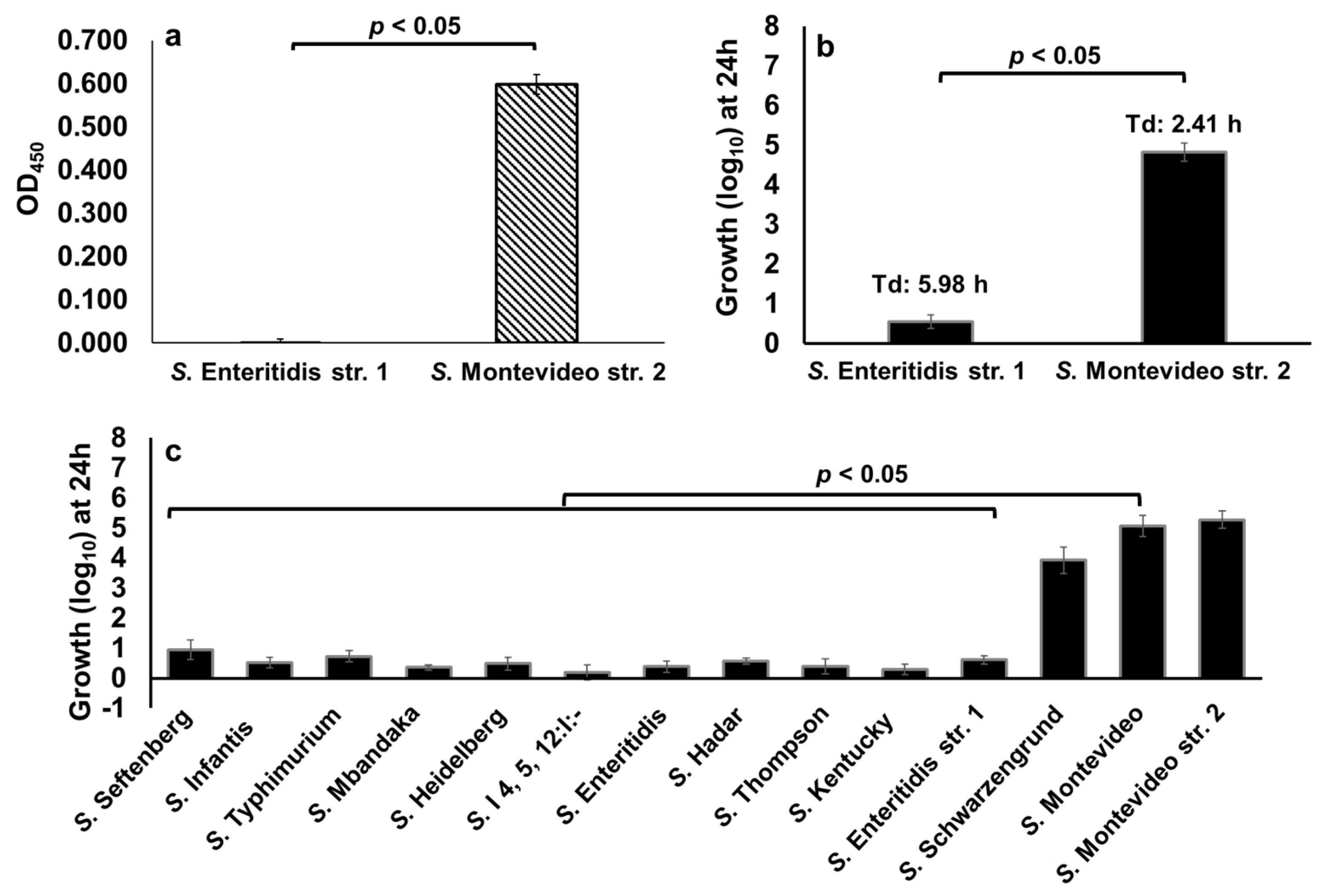
2.6. Clinically Significant NTS Utilize Free-DGA as a Sole Source of Energy for Growth in a Serotype-Independent Manner
2.7. NTS Hydrolyze d-Glucuronide in a Serotype-Dependent Manner
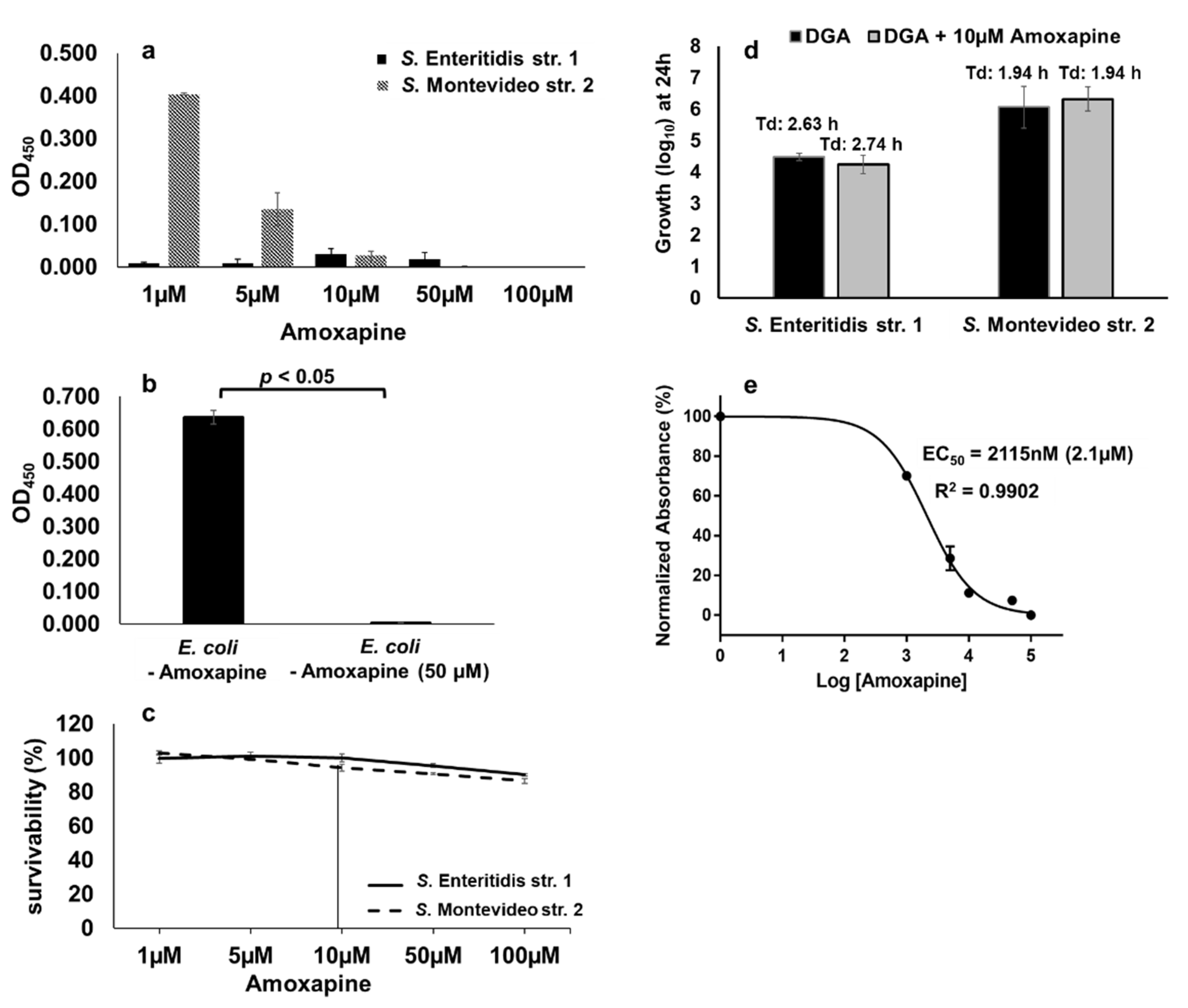
2.8. Amoxapine Inhibits the GUS-Mediated Hydrolysis of d-Glucuronide PNPG by S. Montevideo
2.9. Amoxapine Inhibits the Ability of GUS-Positive NTS Serotypes to Hydrolyze d-Glucuronide AA-Gluc in a Serotype-Independent Manner
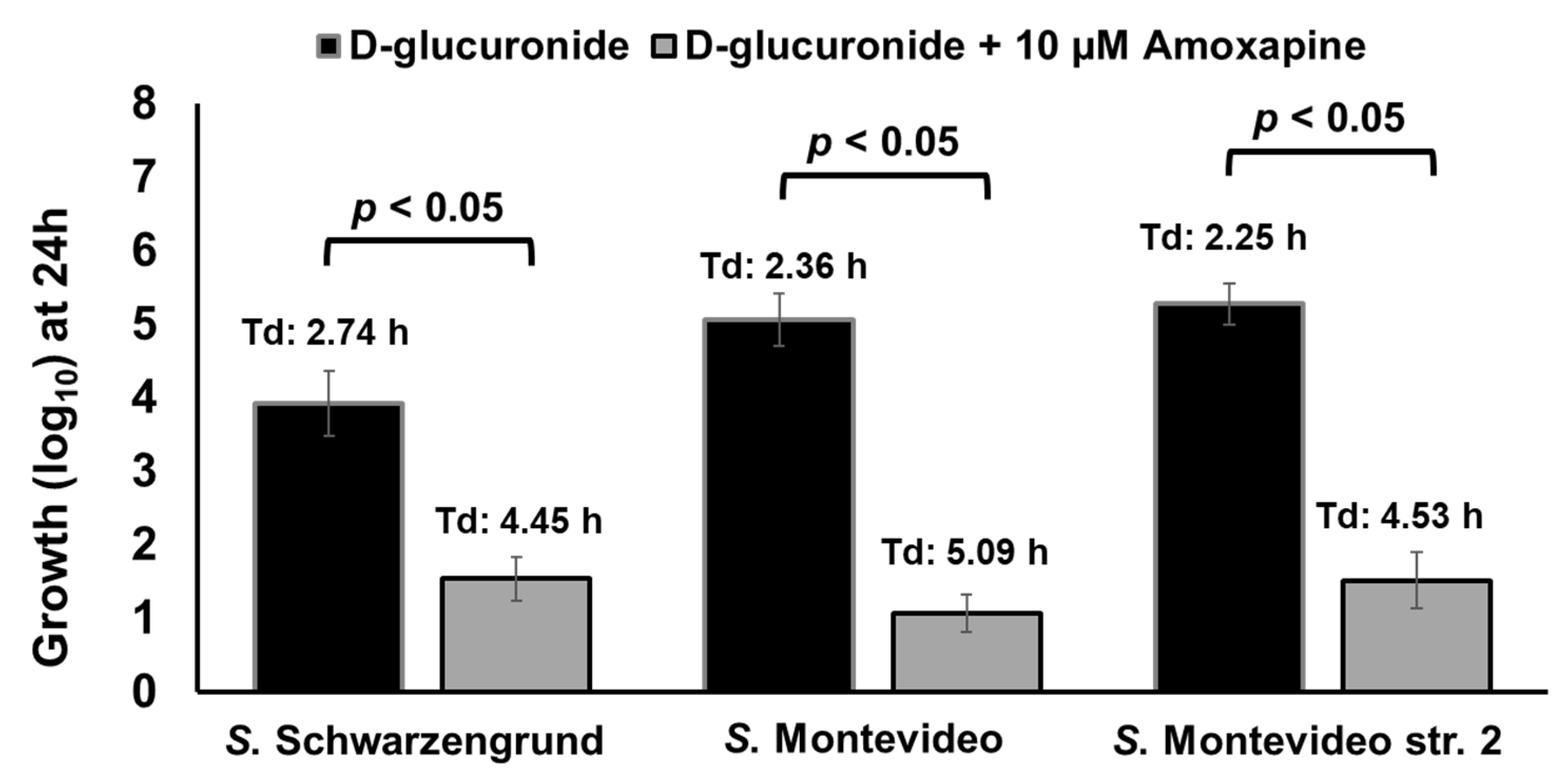
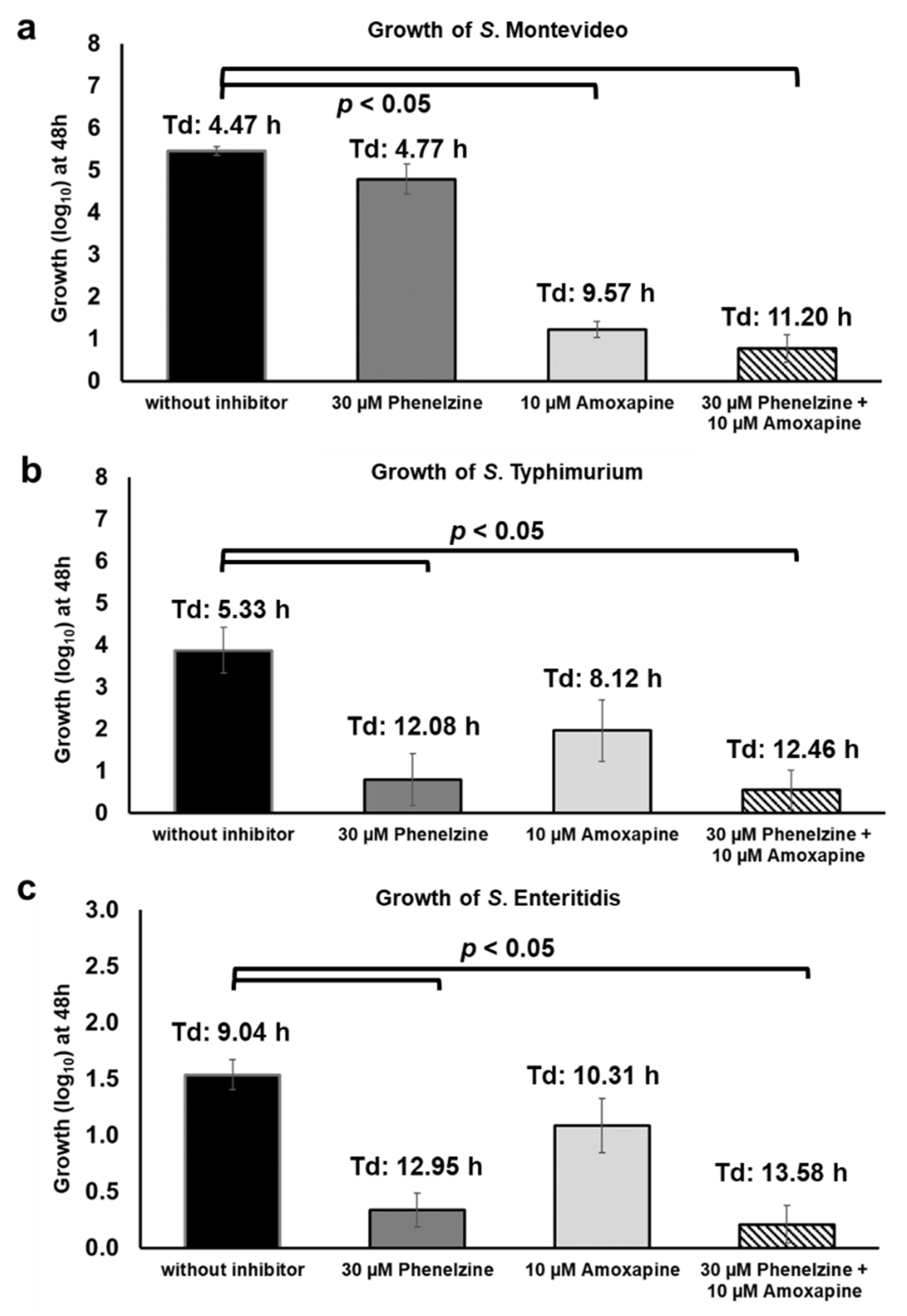
2.10. Combination of Phenelzine and Amoxapine Inhibits the Ability of NTS Serotypes to Derive Energy for Growth from D-Glucuronide and TYR
3. Materials and Methods
3.1. Bacterial Strains
3.2. Cloning, Expression, and Purification of Recombinant SEN2971, SEN3065, and SEN2426
3.3. Determination of Oxidoreductase Activity of Recombinant SEN2971, SEN3065, and SEN2426
3.3.1. Radiometric Assay
3.3.2. Colorimetric Assay
3.4. Cell-Based Oxidoreductase Inhibition Assays
3.5. Cell-Based GUS Activity and Inhibition Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, E.; Sperandio, V. Frenemies: Signaling and nutritional integration in pathogen- microbiota-host interactions. Cell Host Microbe 2015, 18, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Fabich, A.J.; Jones, S.A.; Chowdhury, F.Z.; Cernosek, A.; Anderson, A.; Smalley, D.; Mchargue, J.W.; Hightower, G.A.; Smith, J.T.; Autieri, S.M.; et al. Comparison of Carbon Nutrition for Pathogenic and Commensal Escherichia coli Strains in the Mouse Intestine. Infect. Immun. 2008, 76, 1143–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumler, A.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Staib, L.; Fuchs, T.M. From food to cell: Nutrient exploitation strategies of enteropathogens. Microbiology 2014, 160, 1020–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, C.M.A. The Dynamic Interactions between Salmonella and the Microbiota, within the Challenging Niche of the Gastrointestinal Tract. Int. Sch. Res. Not. 2014, 2014, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, S.; Woodmansey, E.J.; Macfarlane, G.T. Colonization of Mucin by Human Intestinal Bacteria and Establishment of Biofilm Communities in a Two-Stage Continuous Culture System. Appl. Environ. Microbiol. 2005, 71, 7483–7492. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.M.; Ferreyra, J.A.; Higginbottom, S.K.; Lynch, J.B.; Kashyap, P.C.; Gopinath, S.; Naidu, N.; Choudhury, B.; Bart, C.; Monack, D.M.; et al. Microbiota-liberated host sugars facilitate post-antibiotic expansion of enteric pathogens. Nature 2014, 502, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, A.R.; Sperandio, V. Enteric Pathogens Exploit the Microbiota-generated Nutritional Environment of the Gut. Microbiol. Spectr. 2016, 3, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Chaves, F.; Winter, S.E.; Lopez, C.A.; Xavier, M.N.; Winter, M.G.; Nuccio, S.; Russell, J.M.; Laughlin, R.C.; Lawhon, S.D.; Sterzenbach, T.; et al. Salmonella Uses Energy Taxis to Benefit from Intestinal Inflammation. PLoS Pathog. 2013, 9, 1–13. [Google Scholar] [CrossRef]
- Steeb, B.; Claudi, B.; Burton, N.A.; Tienz, P.; Schmidt, A.; Farhan, H.; Bumann, D.; Maze, A. Parallel Exploitation of Diverse Host Nutrients Enhances Salmonella Virulence. PLoS Pathog. 2013, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Elder, J.R.; Paul, N.C.; Burin, R.; Guard, J.; Shah, D.H. Genomic organization and role of SPI-13 in nutritional fitness of Salmonella. Int. J. Med. Microbiol. 2018, 308, 1043–1052. [Google Scholar] [CrossRef]
- Marcobal, A.; Las Rivas, B.D.; Landete, J.M.; Tabera, L.; Ia, M.A.R. Tyramine and Phenylethylamine Biosynthesis by Food Bacteria. Food Sci. Nutr. 2012, 448–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palencia, P.F.; Fernández, M.; Mohedano, M.L.; Ladero, V.; Quevedo, C.; Alvarez, M.A.; López, P. Role of tyramine synthesis by food-borne enterococcus durans in adaptation to the gastrointestinal tract environment. Appl. Environ. Microbiol. 2011, 77, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Bargossi, E.; Gardini, F.; Gatto, V.; Montanari, C.; Torriani, S.; Tabanelli, G. The capability of tyramine production and correlation between phenotypic and genetic characteristics of Enterococcus faecium and Enterococcus faecalis strains. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pircher, A.; Bauer, F.; Paulsen, P. Formation of cadaverine, histamine, putrescine and tyramine by bacteria isolated from meat, fermented sausages and cheeses. Eur. Food Res. Technol. 2007, 226, 225–231. [Google Scholar] [CrossRef]
- Leisner, J.J.; Laursen, B.G.; Prévost, H.; Drider, D.; Dalgaard, P. Carnobacterium: Positive and negative effects in the environment and in foods. FEMS Microbiol. Rev. 2007, 31, 592–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, G.V.; Choi, K.; Klemashevich, C.; Wu, C.; Prabakaran, D.; Pan, L.B.; Steinmeyer, S.; Mueller, C.; Yousofshahi, M.; Alaniz, R.C.; et al. Prediction and quantification of bioactive microbiota metabolites in the mouse gut. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Philips, S.R.; Durden, D.A.; Boulton, A. Identification and distribution of p-tyramine in the rat. Can. J. Biochem. 1974, 366–377. [Google Scholar] [CrossRef]
- Díaz, E.; Ferrández, A.; Prieto, M.A.; García, J.L. Biodegradation of aromatic compounds by Escherichia coli. Microbiol. Mol. Biol. Rev. 2001, 65, 523–569. [Google Scholar] [CrossRef] [Green Version]
- Elovaara, H.; Huusko, T.; Maksimow, M.; Elima, K.; Yegutkin, G.G.; Skurnik, M.; Dobrindt, U.; Siitonen, A.; McPherson, M.J.; Salmi, M.; et al. Primary amine oxidase of Escherichia coli is a metabolic enzyme that can use a human leukocyte molecule as a substrate. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Rankin, L.D.; Bodenmiller, D.M.; Partridge, J.D.; Nishino, S.F.; Spiro, S. Escherichia coli NsrR Regulates a Pathway for the Oxidation of 3-Nitrotyramine to 4-Hydroxy-3-Nitrophenyllacetate. J. Bacteriol. 2008, 190, 6170–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Spiro, S. Finely tuned regulation of the aromatic amine degradation pathway in Escherichia coli. J. Bacteriol. 2013, 195, 5141–5150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burin, R.; Shah, D.H. Global transcriptional profiling of tyramine and D -glucuronic acid catabolism in Salmonella. Int. J. Med. Microbiol. 2020, 310, 151452. [Google Scholar] [CrossRef]
- Finberg, J.P.M.; Rabey, J.M.; Gillman, K. Inhibitors of MAO-A and MAO-B in Psychiatry and Neurology. Front. Pharmacol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bortolato, M.; Chen, K.; Shih, J.C. Monoamine oxidase inactivation: From pathophysiology to therapeutics. Adv. Drug Deliv. Rev. 2008, 60, 1527–1533. [Google Scholar] [CrossRef] [Green Version]
- Rothe, M.; Alpert, C.; Loh, G.; Blaut, M. Novel Insights into E. coli’ s Hexuronate Metabolism: KduI Facilitates the Conversion of Galacturonate and Glucuronate under Osmotic Stress Conditions. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellock, S.J.; Redinbo, M.R. Glucuronides in the gut: Sugar-driven symbioses between microbe and host. J. Biol. Chem. 2017, 292, 8569–8576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniels, A.E.; Rice, E.W.; Reyes, A.L.; Johnson, C.H.; Haugland, R.A.; Stelma, J. Erratum: Confirmational identification of Escherichia coli, a comparison of genotypic and phenotypic assays for glutamate decarboxylase and β-D- glucuronidase (Applied and Environmental Microbiology (1996) 62:9 (3350- 3354)). Appl. Environ. Microbiol. 1998, 64, 4113. [Google Scholar] [CrossRef] [Green Version]
- Feng, P.C.S.; Hartman, P.A. Fluorogenic assays for immediate confirmation of Escherichia coli. Appl. Environ. Microbiol. 1982, 43, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.W.; Brill, J.; Lum, R. Proportion of beta-D-glucuronidase-negative Escherichia coli in human fecal samples. Appl. Environ. Microbiol. 1989, 55, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Kong, R.; Liu, T.; Zhu, X.; Ahmad, S.; Williams, A.L.; Phan, A.T.; Zhao, H.; Scott, J.E.; Yeh, L.A.; Wong, S.T.C. Old drug new use—Amoxapine and its metabolites as potent bacterial β-glucuronidase inhibitors for alleviating cancer drug toxicity. Clin. Cancer Res. 2014, 20, 3521–3530. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.D.; Wang, H.; Lane, K.T.; Scott, J.E.; Orans, J.; Koo, S.; Venkatesh, M.; Jobin, C.; Yeh, L.; Mani, S.; et al. Alleviating Cancer Drug Toxicity by Inhibiting a Bacterial Enzyme. Natl. Inst. Health 2011, 330, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.D.; Roberts, A.B.; Pollet, R.M.; Mani, S.; Kelly, L.; Redinbo, M.R.; Wallace, B.D.; Roberts, A.B.; Pollet, R.M.; Ingle, J.D.; et al. Structure and Inhibition of Microbiome b-Glucuronidases Essential to the Alleviation of Cancer Drug Toxicity. Chem. Biol. 2015, 22, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Hughes, M.A.; Yeh, L.; Scott, J.E. Potential Repurposing of Known Drugs as Potent Bacterial β -Glucuronidase Inhibitors. J. Biomol. Screen. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S. Functional and Genetic Characterization of Sulfatases in Salmonella Enterica Serovar Typhimurium; University of Wisconsin Milwaukee: Milwaukee, WI, USA, 2014. [Google Scholar]
- Murooka, Y.; Harada, T. Regulation of derepressed synthesis of arylsulfatase by tyramine oxidase in Salmonella typhimurium. J. Bacteriol. 1981, 145, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Murooka, Y.; Higashiura, T.; Harada, T. Genetic mapping of tyramine oxidase and arylsulfatase genes and their regulation in intergeneric hybrids of enteric bacteria. J. Bacteriol. 1978, 136, 714–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, J.H.; Suzuki, H.; Azakami, H.; Yamashita, M.; Murooka, Y.; Kumagai, H. Purification, characterization, and crystallization of monoamine oxidase from Escherichia coli K-12. Biosci. Biotechnol. Biochem. 1994, 58, 1652–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, G.; Marwaha, R. Phenelzine. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554508/ (accessed on 2 November 2020).
- Shah, D.H.; Paul, N.C.; Sischo, W.C.; Crespo, R.; Guard, J. Population dynamics and antimicrobial resistance of the most prevalent poultry-associated Salmonella serotypes. Poult. Sci. 2017, 96, 687–702. [Google Scholar] [CrossRef] [PubMed]
- CDC. An Atlas of Salmonella in the United States, 1968–2011; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2013; Volume 1.
- Harhay, D.; Smith, T.P.L.; Bono, J.; Wang, R.; Arthur, T.M.; Bosilevac, J.M.; Harhay, G.P. Annotation of the Salmonella enterica subspecies enterica USDA-ARS-USMARC-1921 complete genome. Unpublished work. 2017. [Google Scholar]
- da Rosa, T.F.; de Souza Machado, C.; Serafin, M.B.; Bottega, A.; Foletto, V.S.; Coelho, S.S.; Hörner, R. Repositioning or Redirection of Antidepressant Drugs in the Treatment of Bacterial and Fungal Infections. Am. J. Ther. 2020, 27, e528–e532. [Google Scholar] [CrossRef] [PubMed]
- Ait Chait, Y.; Mottawea, W.; Tompkins, T.A.; Hammami, R. Unravelling the antimicrobial action of antidepressants on gut commensal microbes. Sci. Rep. 2020, 10, 17878. [Google Scholar] [CrossRef]
- Macedo, D.; Filho, A.J.M.C.; Soares de Sousa, C.N.; Quevedo, J.; Barichello, T.; Júnior, H.V.N.; Freitas de Lucena, D. Antidepressants, antimicrobials or both? Gut microbiota dysbiosis in depression and possible implications of the antimicrobial effects of antidepressant drugs for antidepressant effectiveness. J. Affect. Disord. 2017, 208, 22–32. [Google Scholar] [CrossRef] [PubMed]
- McGovern, A.S.; Hamlin, A.S.; Winter, G. A review of the antimicrobial side of antidepressants and its putative implications on the gut microbiome. Aust. N. Z. J. Psychiatry 2019, 53, 1151–1166. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Doubling Time (h) | ||
|---|---|---|
| TYR | TYR + 30 µM of Phenelzine | |
| S. Thompson | 9.48 | 13.89 * |
| S. Mbandaka | 9.43 | 13.76 * |
| S. Kentucky | 7.89 | 13.61 * |
| S. Infantis | 8.09 | 13.56 * |
| S. Heidelberg | 4.72 | 14.55 * |
| S.Enteritidis str. 1 | 7.45 | 13.61 * |
| S. Enteritidis | 7.26 | 14.12 * |
| S. Montevideo | 6.88 | 13.94 * |
| S.Montevideo str. 2 | 6.80 | 13.83 * |
| S. Hadar | 6.29 | 13.27 * |
| S. Typhimurium | 5.34 | 13.89 * |
| S. Schwarzengrund | 4.86 | 13.29 * |
| S. Seftenberg | 5.00 | 14.55 * |
| S. I 4, 5, 12:I:- | 4.72 | 14.55 * |
| Strain Number | Salmonella Serotype | Strains Used in the Study | Phenotype | AMR Pattern | Reference |
|---|---|---|---|---|---|
| 1 | S. Enteritidis str. CDC_2010K_0968 | WT and ΔSEN2971 | GUS- | N/A | Allard et al., 2013; Elder et al., 2018 |
| 2 | S. Montevideo str. USDA_ARS_USMARC-1921 | WT | GUS+ | N/A | Harhay et al., 2017 |
| 3 | S. Thompson | WT | GUS- | ACSSuTSxtAmcCaz | Shah et al., 2017 |
| 4 | S. Mbandaka | WT | GUS- | ACSSuTAmcCazK | Shah et al., 2017 |
| 5 | S. Kentucky | WT | GUS- | AAmcCCipKNalSSxtT | Shah et al., 2017 |
| 6 | S. Infantis | WT | GUS- | ACGSSuTSxtAmcCaz | Shah et al., 2017 |
| 7 | S. Heidelberg | WT | GUS- | ACSSuTSxtAmcNalCaz | Shah et al., 2017 |
| 8 | S. Enteritidis | WT | GUS- | ACKSSuTSxtAmcCaz | Shah et al., 2017 |
| 9 | S. Hadar | WT | GUS- | ACKSSuTAmc | Shah et al., 2017 |
| 10 | S. Typhimurium | WT | GUS- | ACKSSuTSxtAmcNalCaz | Shah et al., 2017 |
| 11 | S. I4,4,5,12:I:- | WT | GUS- | ACSSuTAmcCaz | Shah et al., 2017 |
| 12 | S. Seftenberg | WT | GUS- | AKSSuTAmcCaz | Shah et al., 2017 |
| 13 | S. Schwarzengrund | WT | GUS+ | ACKSSuTSxt | Shah et al., 2017 |
| 14 | S. Montevideo | WT | GUS+ | ASSuTSxt | Shah et al., 2017 |
| Gene | Primers | Sequence 5′ to 3′ | Product Size (bp) |
|---|---|---|---|
| SEN2971 | RA01clonSEN2971Fw | CACCATGAATACAAAAATCGAT | 1302 |
| RA01clonSEN2971Rv | TTAATTCCGGCCTTTCCAG | ||
| SEN3065 | RA04clonSEN3065Fw | CACCATGATACGTTTCGCTGTA | 999 |
| RA04clonSEN3065Rv | TTACGCAGTAAGGGGATGA | ||
| SEN2426 | RA03clonSEN2426Fw | CACCATGGGTAAACTCACGGGC | 792 |
| RA03clonSEN2426Rv | TCAGACGCCTACGCTTACG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burin, R.; Shah, D.H. Phenelzine and Amoxapine Inhibit Tyramine and d-Glucuronic Acid Catabolism in Clinically Significant Salmonella in A Serotype-Independent Manner. Pathogens 2021, 10, 469. https://doi.org/10.3390/pathogens10040469
Burin R, Shah DH. Phenelzine and Amoxapine Inhibit Tyramine and d-Glucuronic Acid Catabolism in Clinically Significant Salmonella in A Serotype-Independent Manner. Pathogens. 2021; 10(4):469. https://doi.org/10.3390/pathogens10040469
Chicago/Turabian StyleBurin, Raquel, and Devendra H. Shah. 2021. "Phenelzine and Amoxapine Inhibit Tyramine and d-Glucuronic Acid Catabolism in Clinically Significant Salmonella in A Serotype-Independent Manner" Pathogens 10, no. 4: 469. https://doi.org/10.3390/pathogens10040469
APA StyleBurin, R., & Shah, D. H. (2021). Phenelzine and Amoxapine Inhibit Tyramine and d-Glucuronic Acid Catabolism in Clinically Significant Salmonella in A Serotype-Independent Manner. Pathogens, 10(4), 469. https://doi.org/10.3390/pathogens10040469