
Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus


 , , and
, , and
Abstract
:1. Introduction
2. Results
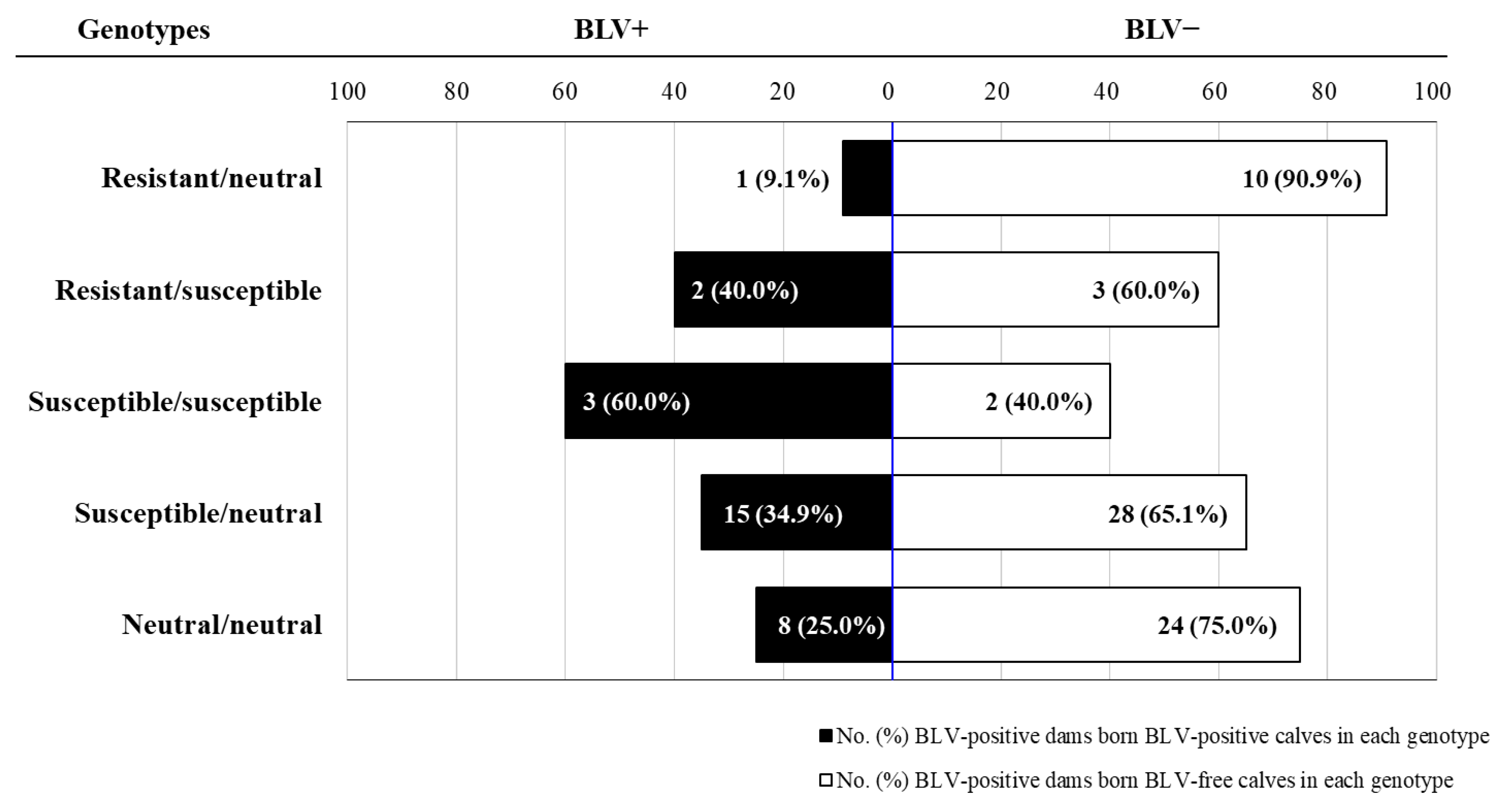
2.1. Risk of Perinatal BLV Transmission in Dams with Different BoLA-DRB3 Genotypes
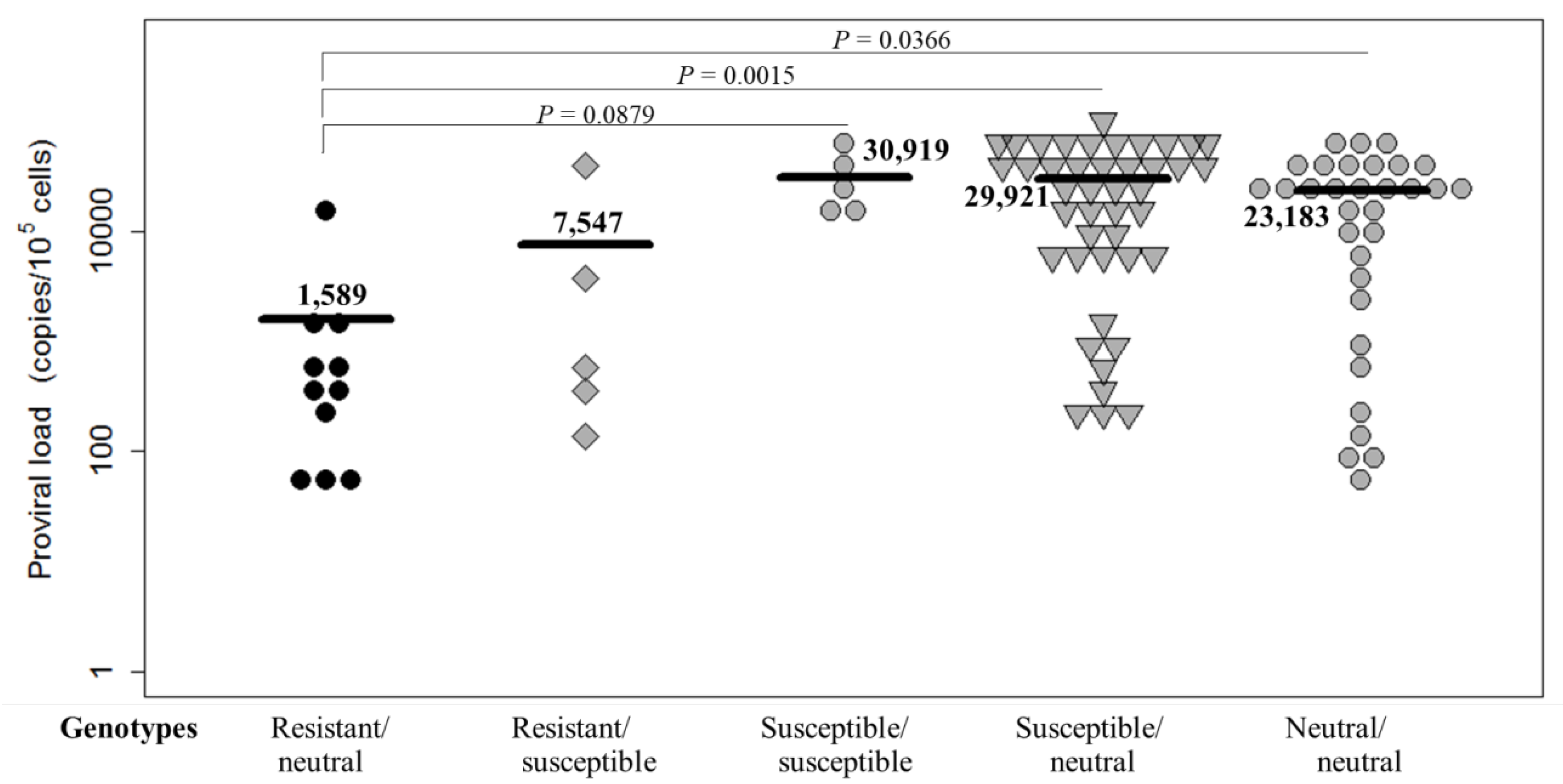
2.2. Distribution of PVLs in Dams with Different BoLA-DRB3 Genotypes
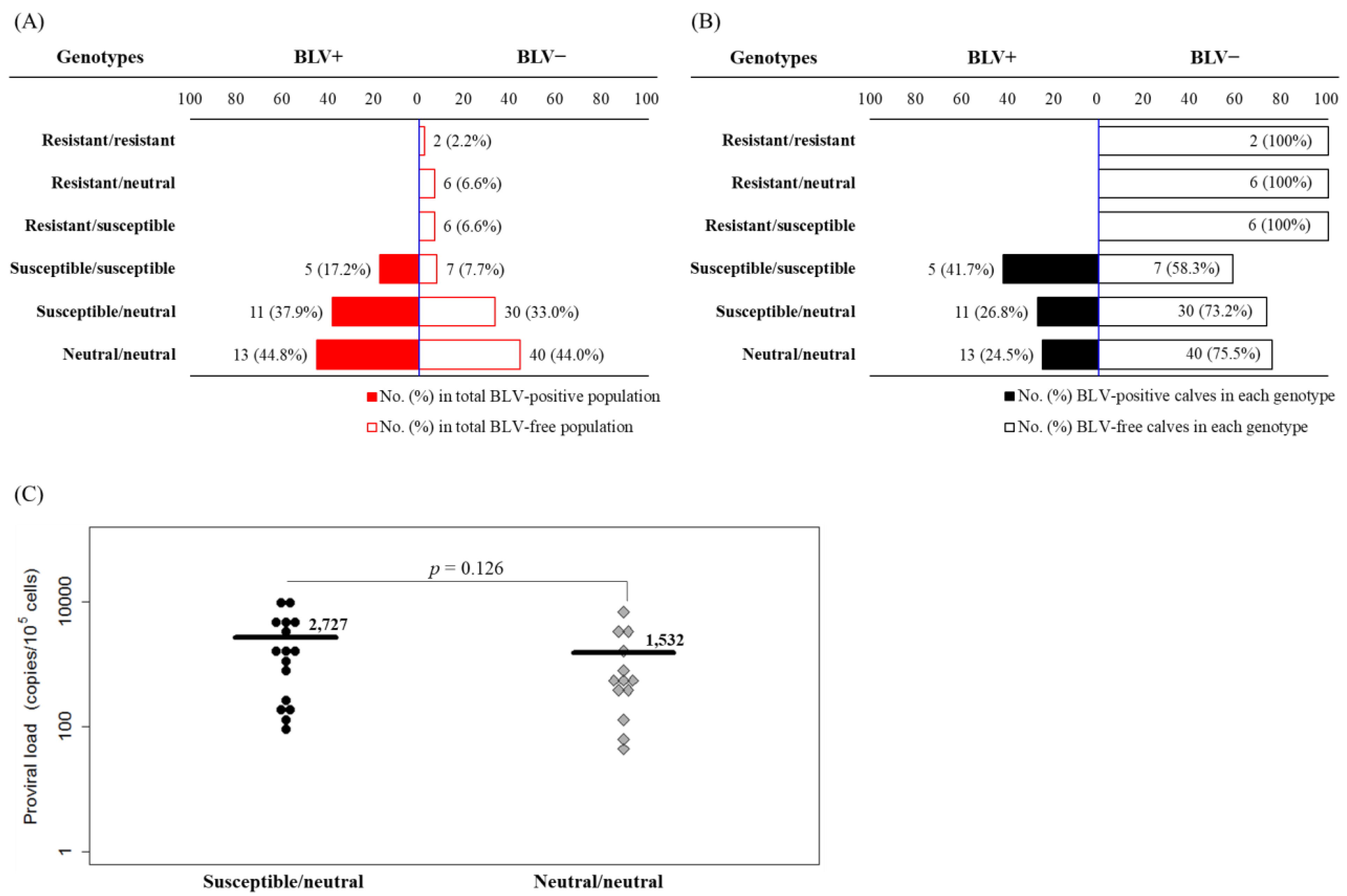
2.3. Frequencies of BLV Provirus in Calves with Different BoLA-DRB3 Genotypes
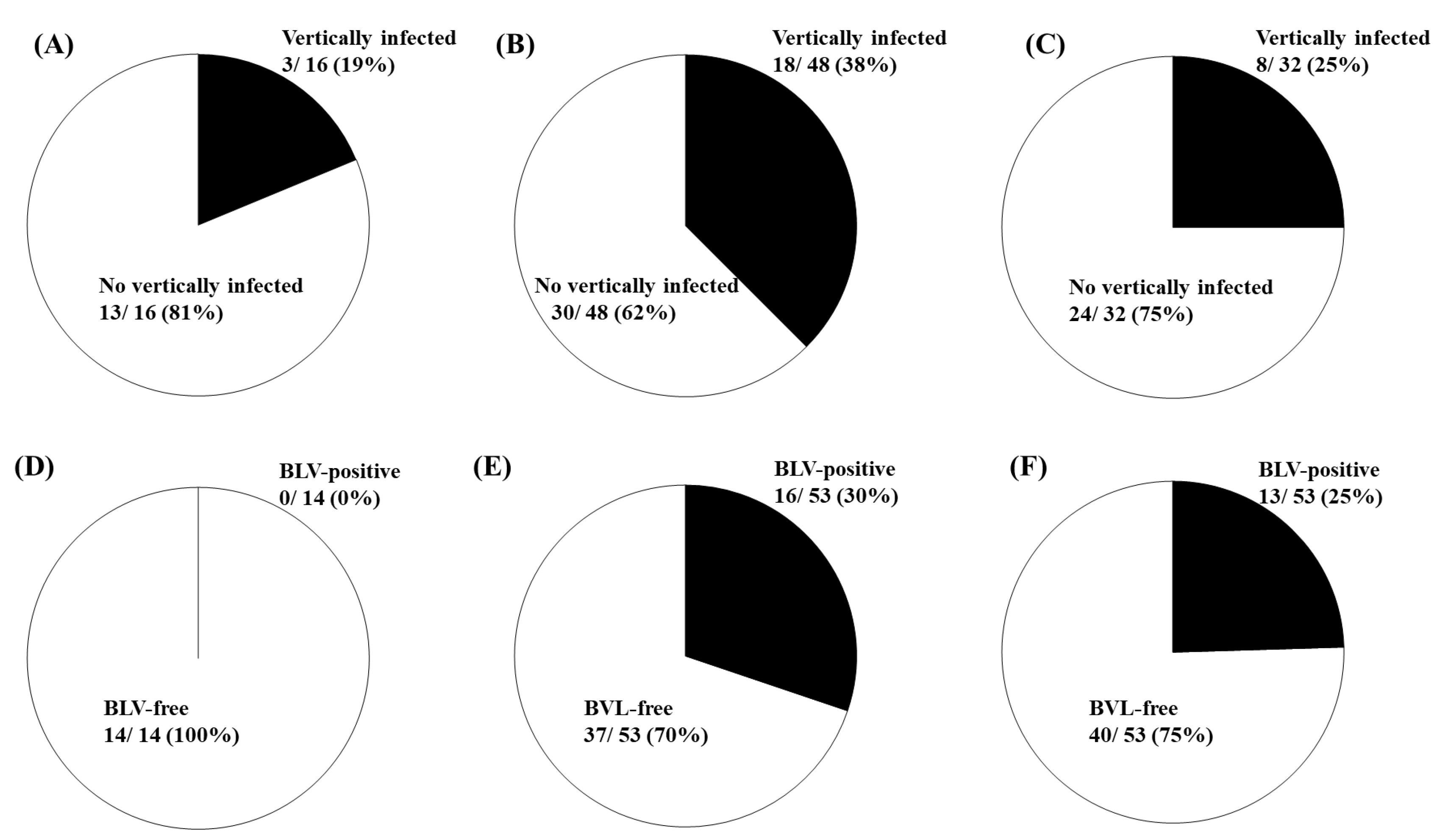
2.4. Differential Risk of BLV Vertical Transmission in Dams and Calves with Different BoLA-DRB3 Genotypes
3. Discussion
4. Materials and Methods
4.1. Clinical Animals
4.2. Ethics Approval
4.3. Collection of Blood Samples, Extraction of Genomic DNA, and Separation of Serum or Plasma
4.4. Enzyme-Linked Immunosorbent Assay (ELISA) for Anti-Env gp51 Antibody
4.5. Quantification of BLV PVL Using the BLV-CoCoMo-qPCR-2 Assay
4.6. BoLA-DRB3 Genotyping
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aida, Y.; Murakami, H.; Takahashi, M.; Takeshima, S.N. Mechanisms of pathogenesis induced by bovine leukemia virus as a model for human T-cell leukemia virus. Front. Microbiol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Panel, E.; Health, A. Enzootic bovine leukosis EFSA Panel on Animal Health and Welfare (AHAW). EFSA J. 2015, 13, 4188. [Google Scholar]
- Gillet, N.; Florins, A.; Boxus, M.; Burteau, C.; Nigro, A.; Vandermeers, F.; Balon, H.; Bouzar, A.B.; Defoiche, J.; Burny, A.; et al. Mechanisms of leukemogenesis induced by bovine leukemia virus: Prospects for novel anti-retroviral therapies in human. Retrovirology 2007, 4, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.I.; Tsutsui, T. Nationwide survey of bovine leukemia virus infection among dairy and beef breeding cattle in Japan from 2009–2011. J. Vet. Med. Sci. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, A.; Takeshima, S.N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res. 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Nekouei, O.; VanLeeuwen, J.; Stryhn, H.; Kelton, D.; Keefe, G. Lifetime effects of infection with bovine leukemia virus on longevity and milk production of dairy cows. Prev. Vet. Med. 2016, 133, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [Green Version]
- Ladronka, R.M.; Ainsworth, S.; Wilkins, M.J.; Norby, B.; Byrem, T.M.; Bartlett, P.C. Prevalence of Bovine Leukemia Virus Antibodies in US Dairy Cattle. Vet. Med. Int. 2018. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.K.; Pelzer, K.D.; Johnson, Y.J. Economic implications of bovine leukemia virus infection in mid-Atlantic dairy herds. J. Am. Vet. Med. Assoc. 2003, 223, 346–352. [Google Scholar] [CrossRef]
- White, T.L.; Moore, D.A. Reasons for whole carcass condemnations of cattle in the United States and implications for producer education and veterinary intervention. J. Am. Vet. Med. Assoc. 2009, 235, 937–941. [Google Scholar] [CrossRef]
- Otta, S.L.; Johnson, R.; Wells, S.J. Association between bovine-leukosis virus seroprevalence and herd-level productivity on US dairy farms. Prev. Vet. Med. 2003, 61, 249–262. [Google Scholar] [CrossRef]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J. Dairy Sci. 2012, 95, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Konnai, S.; Murata, S.; Ohashi, K. Immune exhaustion during chronic infections in cattle. J. Vet. Med. Sci. 2017, 79, 1–5. [Google Scholar] [CrossRef] [Green Version]
- VanLeeuwen, J.A.; Haddad, J.P.; Dohoo, I.R.; Keefe, G.P.; Tiwari, A.; Tremblay, R. Associations between reproductive performance and seropositivity for bovine leukemia virus, bovine viral-diarrhea virus, Mycobacterium avium subspecies paratuberculosis, and Neospora caninum in Canadian dairy cows. Prev. Vet. Med. 2010, 94, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kuczewski, A.; Hogeveen, H.; Orsel, K.; Wolf, R.; Thompson, J.; Spackman, E.; van der Meer, F. Economic evaluation of 4 bovine leukemia virus control strategies for Alberta dairy farms. J. Dairy Sci. 2019, 102, 2578–2592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohara, J.; Takeuchi, M.; Hirano, Y.; Sakurai, Y.; Takahashi, T. Vector control efficacy of fly nets preventing bovine leukemia virus transmission. J. Vet. Med. Sci. 2018, 80, 1524–1527. [Google Scholar] [CrossRef] [Green Version]
- Kohara, J.; Konnai, S.; Onuma, M. Experimental transmission of Bovine leukemia virus in cattle via rectal palpation. Jpn. J. Vet. Res. 2006, 54, 25–30. [Google Scholar]
- Lassauzet, M.L.; Thurmond, M.C.; Johnson, W.O.; Stevens, F.; Picanso, J.P. Effect of brucellosis vaccination and dehorning on transmission of bovine leukemia virus in heifers on a California dairy. Can. J. Vet. Res. 1990, 54, 184. [Google Scholar]
- Mekata, H.; Sekiguchi, S.; Konnai, S.; Kirino, Y.; Honkawa, K.; Nonaka, N.; Horii, Y.; Norimine, J. Evaluation of the natural perinatal transmission of bovine leukaemia virus. Vet. Rec. 2015, 176, 254. [Google Scholar] [CrossRef]
- Ruiz, V.; Porta, N.G.; Lomónaco, M.; Trono, K.; Alvarez, I. Bovine Leukemia virus infection in neonatal calves. Risk factors and control measures. Front. Vet. Sci. 2018, 5, 267. [Google Scholar] [CrossRef] [Green Version]
- Watanuki, S.; Takeshima, S.N.; Borjigin, L.; Sato, H.; Bai, L.; Murakami, H.; Sato, R.; Ishizaki, H.; Matsumoto, Y.; Aida, Y. Visualizing bovine leukemia virus (BLV)-infected cells and measuring BLV proviral loads in the milk of BLV seropositive dams. Vet. Res. 2019, 50, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jimba, M.; Takeshima, S.N.; Matoba, K.; Endoh, D.; Aida, Y. BLV-CoCoMo-qPCR: Quantitation of bovine leukemia virus proviral load using the CoCoMo algorithm. Retrovirology 2010, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Watanuki, S.; Murakami, H.; Sato, R.; Ishizaki, H.; Aida, Y. Development of a luminescence syncytium induction assay (LuSIA) for easily detecting and quantitatively measuring bovine leukemia virus infection. Arch. Virol. 2018, 163, 1519–1530. [Google Scholar] [CrossRef]
- Juliarena, M.A.; Barrios, C.N.; Ceriani, M.C.; Esteban, E.N. Hot topic: Bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J. Dairy Sci. 2016, 99, 4586–4589. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.N.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-CoCoMo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.W.; Borjigin, L.; Saito, S.; Fukunaga, K.; Saitou, E.; Okazaki, K.; Mizutani, T.; Satoshi Wada, S.T.; Aida, Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia. Viruses 2020, 12, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshima, S.; Aida, Y. Structure, function and disease susceptibility of the bovine major histocompatibility complex. Anim. Sci. J. 2006, 77, 138–150. [Google Scholar] [CrossRef]
- Takeshima, S.N.; Ohno, A.; Aida, Y. Bovine leukemia virus proviral load is more strongly associated with bovine major histocompatibility complex class II DRB3 polymorphism than with DQA1 polymorphism in Holstein cow in Japan. Retrovirology 2019, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, T.; Takeshima, S.N.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, G.; Alvarez, I.; Merlini, R.; Rondelli, F.; Trono, K. Dynamics of perinatal bovine leukemia virus infection. BMC Vet. Res. 2014, 10, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, G.E.; Frankena, K. Survival analysis on aggregate data to assess time to sero-conversion after experimental infection with Bovine Leukemia virus. Prev. Vet. Med. 2005, 68, 241–262. [Google Scholar] [CrossRef] [PubMed]
- More, S.; Bøtner, A.; Butterworth, A.; Calistri, P.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Gortázar Schmidt, C.; Michel, V.; et al. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Enzootic bovine leukosis (EBL). EFSA J. 2017, 15, 4956. [Google Scholar]
- Ruggiero, V.J.; Norby, B.; Benitez, O.J.; Hutchinson, H.; Sporer, K.R.B.; Droscha, C.; Swenson, C.L.; Bartlett, P.C. Controlling bovine leukemia virus in dairy herds by identifying and removing cows with the highest proviral load and lymphocyte counts. J. Dairy Sci. 2019, 102, 9165–9175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshima, S.N.; Kitamura-Muramatsu, Y.; Yuan, Y.; Polat, M.; Saito, S.; Aida, Y. BLV-CoCoMo-qPCR-2: Improvements to the BLV-CoCoMo-qPCR assay for bovine leukemia virus by reducing primer degeneracy and constructing an optimal standard curve. Arch. Virol. 2015, 160, 1325–1332. [Google Scholar] [CrossRef]
- Takeshima, S.; Matsumoto, Y.; Miyasaka, T.; Saito, H.; Onuma, M. A new method for typing bovine major histocompatibility complex class II DRB3 alleles by combining two established PCR sequence-based techniques. Tissue Antigens 2011, 78, 208–213. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Year | |||||
|---|---|---|---|---|---|
| Farm | 2017 | 2018 | 2019 | 2020 | Total |
| A | 2 | 5 | 2 | 0 | 9 |
| B | 2 | 2 | 0 | 0 | 4 |
| C | 2 | 3 | 0 | 0 | 5 |
| D | 5 | 3 | 11 | 0 | 19 |
| E | 16 | 18 | 32 | 17 | 83 |
| Total | 27 | 31 | 45 | 17 | 120 |
| Dams Alleles | gp51 a | PVL b | Dams Alleles | gp51 | PVL | Dams Alleles | gp51 | PVL |
|---|---|---|---|---|---|---|---|---|
| Resistant/Neutral Genotypes | Neutral/Neutral Genotypes | |||||||
| 009:02/001:01 | +d | 189 | 012:01/011:01 | + | 17,625 | 011:01/007:01 | + | 78,079 |
| 009:02/001:01 | + | 47 | 012:01/011:01 | + | 11,459 | 011:01/011:01 | + | 70,859 |
| 009:02/010:01 | −e | 0 | 012:01/010:01 | + | 862 | 011:01/011:01 | + | 57,379 |
| 009:02/010:01 | − | 0 | 012:01/016:01 | + | 205 | 002:01/027:03 | + | 47,903 |
| 014:01:01/001:01 | + | 12,544 | 012:01/011:01 | − | 0 | 011:01/001:01 | + | 43,822 |
| 014:01:01/027:03 | + | c 1491 | 015:01/010:01 | + | 83,036 | 011:01/010:01 | + | 42,597 |
| 014:01:01/001:01 | + | 1181 | 015:01/011:01 | + | 72,853 | 011:01/007:01 | + | 40,583 |
| 014:01:01/027:03 | + | 674 | 015:01/001:01 | + | 67,185 | 010:01/001:01 | + | 35,257 |
| 014:01:01/011:01 | + | 490 | 015:01/007:01 | + | 65,839 | 010:01/010:01 | + | 31,068 |
| 014:01:01/007:01 | + | 428 | 015:01/011:01 | + | 63,536 | 011:01/027:03 | + | 28,112 |
| 014:01:01/011:01 | + | 321 | 015:01/001:01 | + | 54,091 | 011:01/010:01 | + | 26,038 |
| 014:01:01/011:01 | + | 68 | 015:01/011:01 | + | 43,868 | 011:01/011:01 | + | 25,977 |
| 014:01:01/011:01 | + | 43 | 015:01/011:01 | + | 42,759 | 010:01/010:01 | + | 25,839 |
| 014:01:01/027:03 | − | 0 | 015:01/027:03 | + | 41,824 | 011:01/010:01 | + | 25,547 |
| 014:01:01/007:01 | − | 0 | 015:01/001:01 | + | 40,714 | 011:01/010:01 | + | 25,547 |
| 014:01:01/011:01 | − | 0 | 015:01/011:01 | + | 38,416 | 001:01/001:01 | + | 25,031 |
| 014:01:01/011:01 | − | 0 | 015:01/011:01 | + | 38,371 | 010:01/001:01 | + | 22,180 |
| Resistant/susceptible genotypes | 015:01/011:01 | + | 33,300 | 010:01/010:01 | + | 20,134 | ||
| 014:01:01/015:01 | + | 33,529 | 015:01/001:01 | + | 24,249 | 011:01/027:03 | + | 18,607 |
| 014:01:01/015:01 | + | 3122 | 015:01/011:01 | + | 22,690 | 010:01/010:01 | + | 13,613 |
| 014:01:01/015:01 | + | 360 | 015:01/010:01 | + | 20,238 | 010:01/027:03 | + | 11,914 |
| 014:01:01/012:01 | + | 570 | 015:01/011:01 | + | 16,800 | 011:01/027:03 | + | 10,928 |
| 014:01:01/012:01 | + | 153 | 015:01/001:01 | + | 13,247 | 001:01/010:01 | + | 6876 |
| Susceptible/susceptible genotypes | 015:01/011:01 | + | 9196 | 010:01/001:01 | + | 3955 | ||
| 012:01/015:01 | + | 52,072 | 015:01/011:01 | + | 7288 | 010:01/001:01 | + | 1893 |
| 012:01/015:01 | + | 28,311 | 015:01/010:01 | + | 7063 | 010:01/001:01 | + | 794 |
| 012:01/015:01 | − | 0 | 015:01/010:01 | + | 6252 | 010:01/010:01 | + | 684 |
| 015:01/015:01 | + | 45,321 | 015:01/010:01 | + | 6077 | 011:01/001:01 | + | 223 |
| 015:01/015:01 | + | 16,735 | 015:01/011:01 | + | 6000 | 011:01/011:01 | + | 144 |
| 015:01/015:01 | + | 12,157 | 015:01/001:01 | + | 1727 | 011:01/001:01 | + | 109 |
| Susceptible/neutral genotypes | 015:01/001:01 | + | 1043 | 011:01/001:01 | + | 106 | ||
| 012:01/001:01 | + | 61,538 | 015:01/007:01 | + | 616 | 011:01/001:01 | + | 66 |
| 012:01/007:04 | + | 59,974 | 015:01/007:01 | + | 420 | 010:01/007:01 | − | 0 |
| 012:01/007:01 | + | 59,441 | 015:01/001:01 | + | 279 | 011:01/010:01 | − | 0 |
| 012:01/007:04 | + | 57,550 | 015:01/007:01 | + | 275 | 011:01/010:01 | − | 0 |
| 012:01/011:01 | + | 51,915 | 015:01/001:01 | − | 0 | 011:01/018:01 | − | 0 |
| 012:01/001:01 | + | 44,098 | 015:01/011:01 | − | 0 | 001:01/027:03 | − | 0 |
| 012:01/001:01 | + | 42,750 | 015:01/001:01 | − | 0 | 001:01/027:03 | − | 0 |
| 012:01/027:03 | + | 30,863 | 015:01/011:01 | − | 0 | 002:01/016:01 | − | 0 |
| 012:01/027:03 | + | 19,058 | 015:01/001:01 | − | 0 | 001:01/027:01 | − | 0 |
| 011:01/007:01 | − | 0 | ||||||
| 011:01/001:01 | − | 0 | ||||||
| 001:01/007:01 | − | 0 | ||||||
| BoLA-DRB3 Genotype | Dam no./Total no. (%) | Dam no. (%) with | ||
|---|---|---|---|---|
| All Dams a | BLV-Positive Dams b | BLV-Negative Calves c | BLV-Positive Calves d | |
| Resistant/neutral | 17/120 (14.2) | 11/96 (11.5) | 10/67 (14.9) | 1/29 (3.4) |
| Resistant/susceptible | 5/120 (4.2) | 5/96 (5.2) | 3/67 (4.5) | 2/29 (6.9) |
| Susceptible/susceptible | 6/120 (5.0) | 5/96 (5.2) | 2/67 (3.0) | 3/29 (10.3) |
| Susceptible/neutral | 49/120 (40.8) | 43/96 (44.8) | 28/67 (41.8) | 15/29 (51.7) |
| Neutral/neutral | 43/120 (35.8) | 32/96 (33.3) | 24/67 (35.8) | 8/29 (27.6) |
| Total | 120 | 96 | 67 | 29 |
| Calves Alleles | PVL | Calves Alleles | PVL | Calves Alleles | PVL |
|---|---|---|---|---|---|
| Resistant/neutral genotypes | |||||
| 009:02/010:01 | 0 | 015:01/011:01 | 0 | 001:01/011:01 | 0 |
| 009:02/010:01 | 0 | 015:01/011:01 | 0 | 001:01/011:01 | 0 |
| 009:02/002:01 | 0 | 015:01/011:01 | 0 | 001:01/011:01 | 0 |
| 014:01:01/011:01 | 0 | 015:01/011:01 | 0 | 001:01/010:01 | 3721 |
| 014:01:01/011:01 | 0 | 015:01/010:01 | 3654 | 001:01/010:01 | 3255 |
| 014:01:01/001:01 | 0 | 015:01/010:01 | 0 | 001:01/010:01 | 374 |
| Resistant/resistant genotypes | 015:01/010:01 | 0 | 001:01/010:01 | 0 | |
| 014:01:01/014:01:01 | 0 | 015:01/010:01 | 0 | 001:01/010:01 | 0 |
| 014:01:01/014:01:01 | 0 | 015:01/010:01 | 0 | 001:01/010:01 | 0 |
| Resistant/susceptible genotypes | 015:01/010:01 | 0 | 001:01/010:01 | 0 | |
| 014:01:01/015:01 | 0 | 015:01/001:01 | 174 | 001:01/010:01 | 0 |
| 014:01:01/015:01 | 0 | 015:01/001:01 | 0 | 001:01/010:01 | 0 |
| 014:01:01/015:01 | 0 | 015:01/001:01 | 0 | 001:01/007:01 | 0 |
| 014:01:01/015:01 | 0 | 015:01/001:01 | 0 | 001:01/007:01 | 0 |
| 014:01:01/015:01 | 0 | 015:01/001:01 | 0 | 001:01/002:01 | 0 |
| 014:01:01/012:01 | 0 | 015:01/001:01 | 0 | 001:01/001:01 | 59 |
| Susceptible/susceptible genotypes | 015:01/001:01 | 0 | 001:01/001:01 | 0 | |
| 012:01/015:01 | 4044 | 015:01/001:01 | 0 | 001:01/001:01 | 0 |
| 012:01/015:01 | 1496 | 015:01/007:01 | 0 | 001:01/001:01 | 0 |
| 012:01/015:01 | 0 | 015:01/002:01 | 0 | 001:01/001:01 | 0 |
| 012:01/015:01 | 0 | 012:01/011:01 | 0 | 001:01/001:01 | 0 |
| 012:01/015:01 | 0 | 012:01/011:01 | 0 | 001:01/001:01 | 0 |
| 012:01/015:01 | 0 | 012:01/010:01 | 1366 | 001:01/001:01 | 0 |
| 015:01/015:01 | 9988 | 012:01/010:01 | 246 | 001:01/001:01 | 0 |
| 015:01/015:01 | 662 | 012:01/007:01 | 4525 | 002:01/011:01 | 0 |
| 015:01/015:01 | 183 | 012:01/007:01 | 0 | 002:01/007:01 | 42 |
| 015:01/015:01 | 0 | 012:01/005:03 | 0 | 005:03/010:01 | 0 |
| 015:01/015:01 | 0 | 012:01/018:01 | 0 | 007:01/027:03 | 0 |
| 015:01/015:01 | 0 | 012:01/016:01 | 0 | 007:01/011:01 | 147 |
| Susceptible/neutral genotypes | Neutral/neutral genotypes | 007:01/007:01 | 0 | ||
| 015:01/027:03 | 5086 | 010:01/007:04 | 0 | 010:01/027:03 | 472 |
| 015:01/027:03 | 1096 | 011:01/027:03 | 8046 | 010:01/027:03 | 0 |
| 015:01/027:03 | 106 | 011:01/027:03 | 0 | 010:01/027:03 | 0 |
| 015:01/027:03 | 0 | 001:01/027:03 | 553 | 010:01/011:01 | 721 |
| 015:01/027:03 | 0 | 001:01/027:03 | 0 | 010:01/011:01 | 0 |
| 015:01/018:01 | 0 | 001:01/027:03 | 0 | 010:01/011:01 | 0 |
| 015:01/016:01 | 9355 | 001:01/027:03 | 0 | 010:01/010:01 | 0 |
| 015:01/016:01 | 0 | 001:01/027:03 | 0 | 011:01/010:01 | 0 |
| 015:01/011:01 | 1545 | 001:01/016:01 | 0 | 011:01/011:01 | 420 |
| 015:01/011:01 | 109 | 001:01/011:01 | 1562 | 011:01/011:01 | 0 |
| 015:01/011:01 | 0 | 001:01/011:01 | 545 | 011:01/011:01 | 0 |
| 015:01/011:01 | 0 | 001:01/011:01 | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borjigin, L.; Lo, C.-W.; Bai, L.; Hamada, R.; Sato, H.; Yoneyama, S.; Yasui, A.; Yasuda, S.; Yamanaka, R.; Mimura, M.; et al. Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus. Pathogens 2021, 10, 502. https://doi.org/10.3390/pathogens10050502
Borjigin L, Lo C-W, Bai L, Hamada R, Sato H, Yoneyama S, Yasui A, Yasuda S, Yamanaka R, Mimura M, et al. Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus. Pathogens. 2021; 10(5):502. https://doi.org/10.3390/pathogens10050502
Chicago/Turabian StyleBorjigin, Liushiqi, Chieh-Wen Lo, Lanlan Bai, Rania Hamada, Hirotaka Sato, Shuji Yoneyama, Anna Yasui, Sohei Yasuda, Risa Yamanaka, Munehito Mimura, and et al. 2021. "Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus" Pathogens 10, no. 5: 502. https://doi.org/10.3390/pathogens10050502
APA StyleBorjigin, L., Lo, C. -W., Bai, L., Hamada, R., Sato, H., Yoneyama, S., Yasui, A., Yasuda, S., Yamanaka, R., Mimura, M., Inokuma, M., Shinozaki, Y., Tanaka, N., Takeshima, S. -N., & Aida, Y. (2021). Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus. Pathogens, 10(5), 502. https://doi.org/10.3390/pathogens10050502