
Isolation and Characterization of Nocardiae Associated with Foaming Coastal Marine Waters




Abstract
:1. Introduction
2. Results
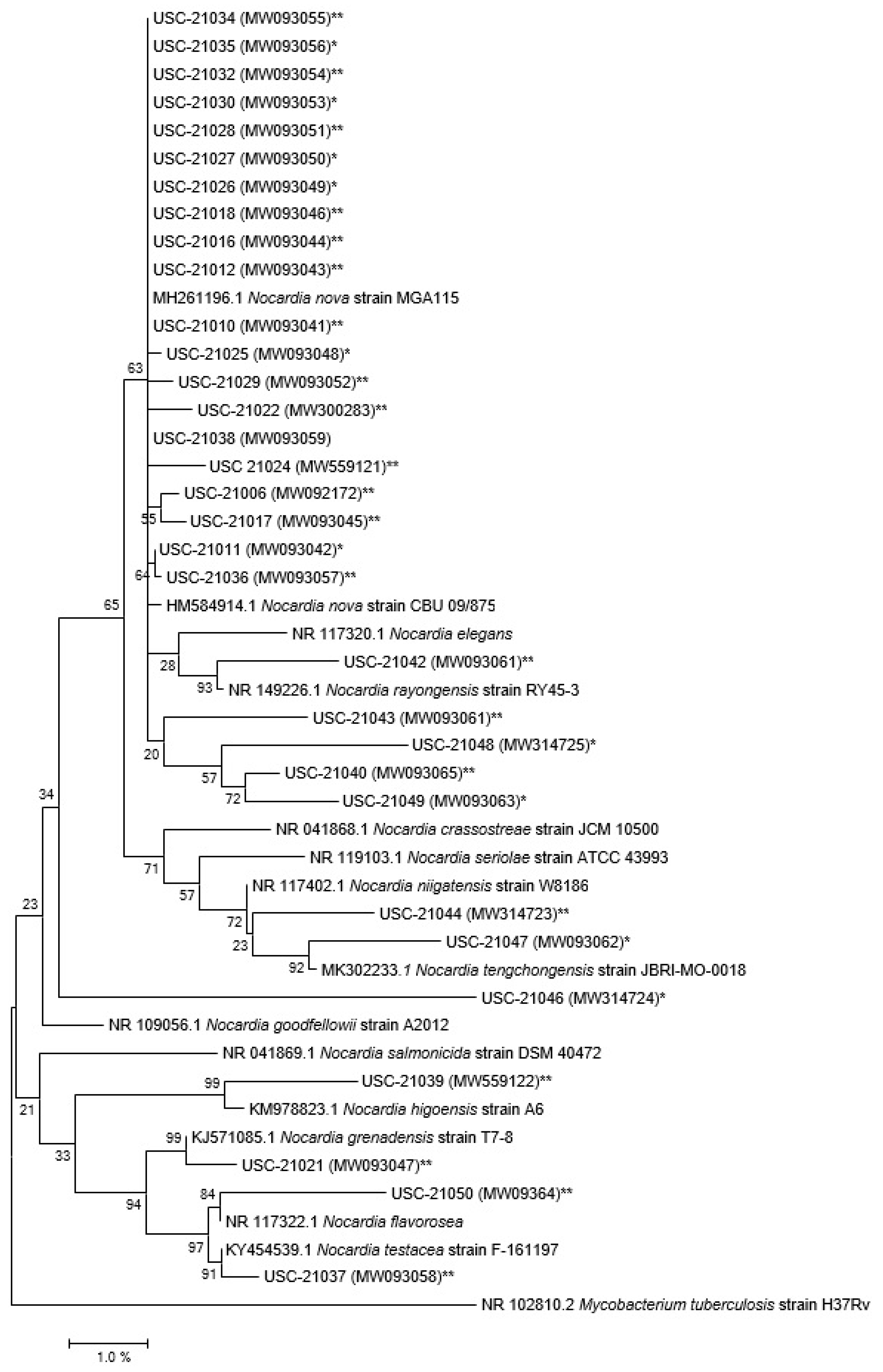
2.1. Selective Isolation and Molecular Level Identification of Nocardiae
2.2. Antibiotic Susceptibility Profiles of the Isolates
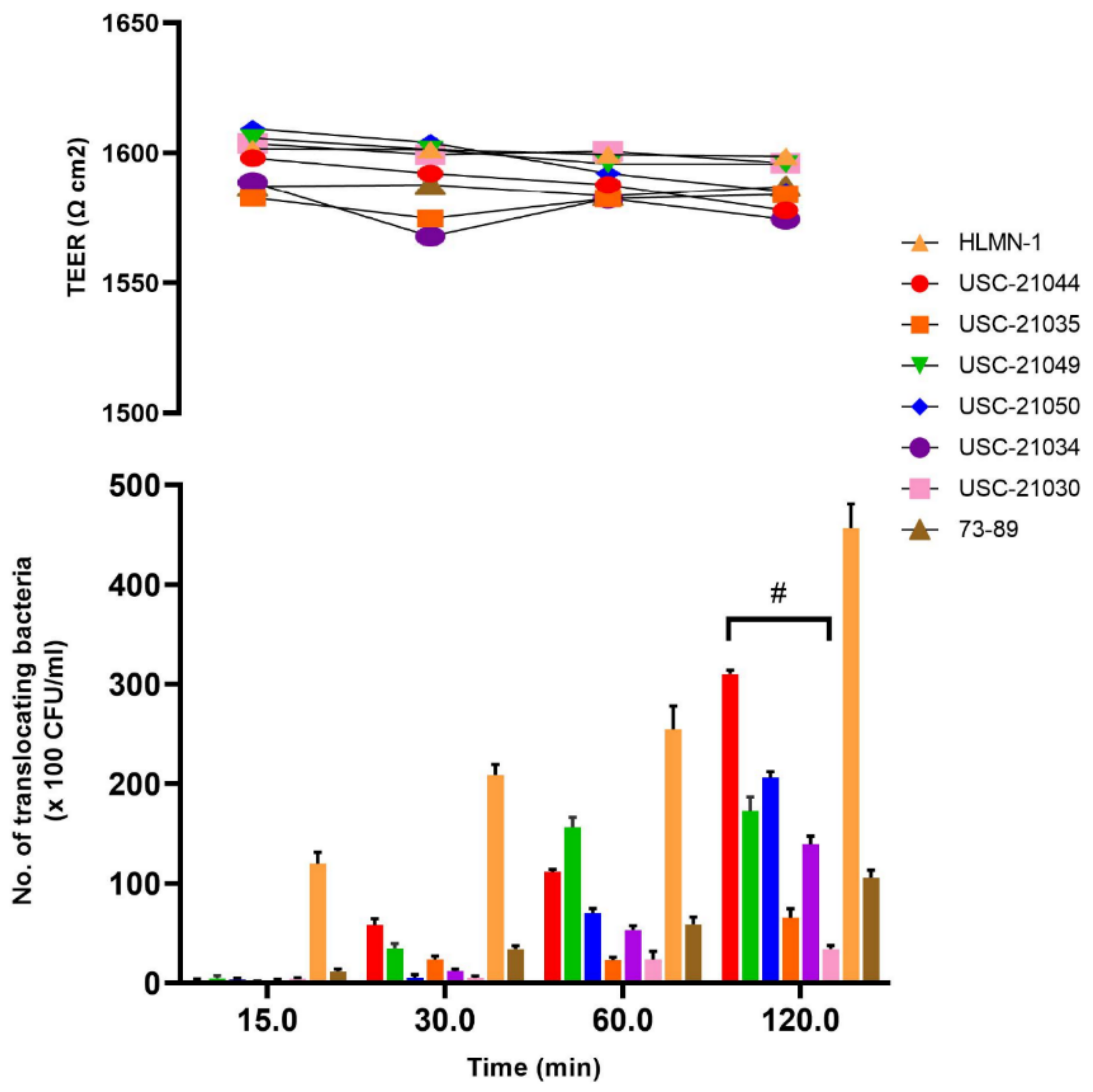
2.3. Adhesion of Nocardia Isolates to Calu-3 Cells
2.4. Translocation of Nocardia Isolates
2.5. Detection of Nocardia-Specific Phage Susceptibility of the Isolates
3. Discussion
4. Materials and Methods
4.1. Sampling Sites and Sample Types
4.2. Isolation of Nocardiae Using a Novel Isolation Technique
4.3. Molecular Characterization of the Isolates
4.4. Detection of Antibiotic Susceptibility Patterns of the Isolates
4.5. Calu-3 Cell Line
4.6. Adhesion and Translocation Assays
4.7. Trans-Epithelial Electrical Resistance Measurements and Translocation Assay
4.8. Detection of Nocardiae Specific Phage Susceptibility of the Isolates
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.-P.; Goodfellow, M.; Göker, M. Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [Green Version]
- Camozzota, C.; Goldman, A.; Tchernev, G.; Lotti, T.; Wollina, U. A Primary Cutaneous Nocardiosis of the Hand. Open Access Maced. J. Med Sci. 2017, 5, 470–472. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M. Nocardia and Related Genera. In Topley & Wilson’s Microbiology & Microbiological Infections; Balows, A., Duerden, B., Eds.; Edward Arnold: London, UK, 1998; pp. 463–489. [Google Scholar]
- Torres, O.H.; Domingo, P.; Pericas, R.; Boiron, P.; Montiel, J.A.; Vázquez, G. Infection caused by Nocardia farcinica: Case report and review. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 205–212. [Google Scholar] [CrossRef]
- Hashemi-Shahraki, A.; Heidarieh, P.; Bostanabad, S.Z.; Hashemzadeh, M.; Feizabadi, M.M.; Schraufnagel, D.; Mirsaeidi, M. Genetic diversity and antimicrobial susceptibility of Nocardia species among patients with nocardiosis. Sci. Rep. 2015, 5, 17862. [Google Scholar] [CrossRef]
- Desai, H.; Wickstrom, K.; Low, S.; Mahmoud, N. Nocardia: An unusual bacteria in an unexpected patient. Am. J. Respir. Crit. Care Med. 2016, 193, 1–18. [Google Scholar]
- Pitt, P.; Jenkins, D. Causes and control of Nocardia in activated sludge. J. Water Pollut. Control Fed. 1990, 62, 143–150. [Google Scholar]
- Thomas, J.A.; Soddell, J.A.; Kurtböke, D.I. Fighting foam with phages? Water Sci. Technol. 2002, 46, 511–518. [Google Scholar]
- Ovez, S.; Orhon, D. Microbial Ecology of Bulking and Foaming Activated Sludge Treating Tannery Wastewater. J. Environ. Sci. Health Part A 2005, 40, 409–422. [Google Scholar] [CrossRef]
- Pajdak-Stós, A.; Kocerba-Soroka, W.; Fyda, J.; Sobczyk, M.; Fiałkowska, E. Foam-forming bacteria in activated sludge effectively reduced by rotifers in laboratory- and real-scale wastewater treatment plant experiments. Environ. Sci. Pollut. Res. 2017, 24, 13004–13011. [Google Scholar] [CrossRef] [Green Version]
- Wandl, G.; Müller-Rechberger, H.; Matsché, N.; Svardal, K.; Winkler, S. Two stage activated sludge plants—Influence of different operational modes on sludge bulking and nitrification. Water Sci. Technol. 2002, 46, 479–486. [Google Scholar] [CrossRef]
- DeWitt, D.; Wagoner, D.L.; Jarrell, J. Use of On-Line Analyzers—Identification of New Approaches for WWTP Process Control and Optimization. Proc. Water Environ. Fed. 2013, 2013, 6766–6776. [Google Scholar] [CrossRef]
- Jenkins, D.; Richard, M.G.; Daigger, G.T. Manual on the Causes and Control of Activated Sludge Bulking, Foaming, and Other Solids Separation Problems; Lewis Publishers: Boca Raton, FL, USA, 2004. [Google Scholar]
- Schilling, K.; Zessner, M. Foam in the aquatic environment. Water Res. 2011, 45, 4355–4366. [Google Scholar] [CrossRef]
- Fryer, M.; Gray, N. Foaming Scum Index (FSI)—A new tool for the assessment and characterisation of biological mediated activated sludge foams. J. Environ. Manag. 2012, 110, 8–19. [Google Scholar] [CrossRef]
- Kurtböke, D.I. (Lab Report) ’Chocolate mousse’ on Sunshine Coast beaches. Microbiol. Aust. 2008, 29, 104–105. [Google Scholar]
- Kurtböke, D.I. Actinomycetes in Biodiscovery: Genomic Advances and New Horizons. In The Handbook of Microbial Resources; Gupta, V.K., Sharma, G.D., Tuohy, M.G., Gaur, R., Eds.; CAB International Publications: Oxfordshire, UK, 2016; pp. 567–590. [Google Scholar]
- Kurtbӧke, D.I. Ecology and Habitat Distribution of Actinobacteria. In Biology and Biotechnology of Actinobacteria; Wink, J., Hamedi, J., Eds.; Springer: Berlin, Germany, 2017; pp. 123–149. [Google Scholar]
- Genetic and Rare Diseases Information Center (GARD). Available online: https://rarediseases.info.nih.gov/diseases/7210/nocardiosis (accessed on 15 March 2020).
- Schlaberg, R.; Fisher, M.A.; Hanson, K.E. Susceptibility Profiles of Nocardia Isolates Based on Current Taxonomy. Antimicrob. Agents Chemother. 2013, 58, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Kandi, V. Human Nocardia Infections: A Review of Pulmonary Nocardiosis. Cureus 2015, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mehta, H.H.; Shamoo, Y. Pathogenic Nocardia: A diverse genus of emerging pathogens or just poorly recognized? PLoS Pathog. 2020, 16, e1008280. [Google Scholar] [CrossRef] [Green Version]
- Beaman, B.L.; Beaman, L. Nocardia Asteroides as an Invasive, Intracellular Pathogen of the Brain and Lungs. In Bacterial Invasion into Eukaryotic Cells; Oelschlaeger, T.A., Hacker, J., Eds.; Springer: New York, NY, USA, 2000; pp. 167–197. [Google Scholar]
- Trevino-Villarreal, J.H.; Vera-Cabrera, L.; Valero-Guillén, P.L.; Salinas-Carmona, M.C. Nocardia brasiliensis Cell Wall Lipids Modulate Macrophage and Dendritic Responses That Favor Development of Experimental Actinomycetoma in BALB/c Mice. Infect. Immun. 2012, 80, 3587–3601. [Google Scholar] [CrossRef] [Green Version]
- Christova, N.; Lang, S.; Wray, V.; Kaloyanov, K.; Konstantinov, S.; Stoineva, I. Production, Structural Elucidation, and In Vitro Antitumor Activity of Trehalose Lipid Biosurfactant from Nocardia farcinica Strain. J. Microbiol. Biotechnol. 2015, 25, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Tournu, H.; Fiori, A.; Van Dijck, P. Relevance of Trehalose in Pathogenicity: Some General Rules, Yet Many Exceptions. PLoS Pathog. 2013, 9, e1003447. [Google Scholar] [CrossRef] [Green Version]
- Beaman, B.L. Differential binding of Nocardia asteroides in the murine lung and brain suggests multiple ligands on the nocardial surface. Infect. Immun. 1996, 64, 4859–4862. [Google Scholar] [CrossRef] [Green Version]
- Tam, S.; Barry, D.P.; Beaman, L.; Beaman, B.L. Neuroinvasive Nocardia asteroides GUH-2 Induces Apoptosis in the Substantia Nigra in Vivo and Dopaminergic Cells in Vitro. Exp. Neurol. 2002, 177, 453–460. [Google Scholar] [CrossRef]
- Barry, D.P.; Beaman, B.L. Nocardia asteroides strain GUH-2 induces proteasome inhibition and apoptotic death of cultured cells. Res. Microbiol. 2007, 158, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Brown-Elliott, B.A.; Conville, P.; Wallace, R.J. Current Status of Nocardia Taxonomy and Recommended Identification Methods. Clin. Microbiol. Newsl. 2015, 37, 25–32. [Google Scholar] [CrossRef]
- Macrogen. 2020. Available online: http://www.macrogen.com (accessed on 24 March 2020).
- Kurtböke, D.I. Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003. [Google Scholar]
- Kurtböke, D.I. Biodiscovery from rare actinomycetes: An eco-taxonomical perspective. Appl. Microbiol. Biotechnol. 2012, 93, 1843–1852. [Google Scholar] [CrossRef]
- Azadi, D.; Shojaei, H. Biodegradation of polycyclic aromatic hydrocarbons, phenol and sodium sulfate by Nocardia species isolated and characterized from Iranian ecosystems. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Vautrin, F.; Pujic, P.; Paquet, C.; Bergeron, E.; Mouniée, D.; Marchal, T.; Salord, H.; Bonnet, J.-M.; Cournoyer, B.; Winiarski, T.; et al. Microbial risk assessment of Nocardia cyriacigeorgica in polluted environments, case of urban rainfall water. Comput. Struct. Biotechnol. J. 2021, 19, 384–400. [Google Scholar] [CrossRef]
- Stratton, H.; Seviour, R.; Soddell, J.; Blackall, L.; Muir, D. The opportunistic pathogen Nocardia farcinica is a foam-producing bacterium in activated sludge plants. Lett. Appl. Microbiol. 1996, 22, 342–346. [Google Scholar] [CrossRef]
- Alavidze, Z.; Aminov, R.; Betts, A.; Bardiau, M.; Bretaudeau, L.; Caplin, J.; Nino, C.; Coffey, A.; Cooper, I.; De Vos, D.; et al. Silk route to the acceptance and re-implementation of bacteriophage therapy-expert round table on acceptance and re-implementation of bacteriophage therapy. Biotechnol. J. 2016, 11, 595–600. [Google Scholar]
- Sybesma, W.; Rohde, C.; Bardy, P.; Pirnay, J.-P.; Cooper, I.; Caplin, J.; Chanishvili, N.; Coffey, A.; De Vos, D.; Scholz, A.H.; et al. Silk Route to the Acceptance and Re-Implementation of Bacteriophage Therapy—Part II. Antibiotics 2018, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Lebeaux, D.; Bergeron, E.; Berthet, J.; Djadi-Prat, J.; Mouniée, D.; Boiron, P.; Lortholary, O.; Rodriguez-Nava, V. Antibiotic susceptibility testing and species identification of Nocardia isolates: A retrospective analysis of data from a French expert laboratory, 2010–2015. Clin. Microbiol. Infect. 2019, 25, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, R.; Caron, F.; Cavallo, J.; Chardon, H.; Chidiac, C.; Courvalin, P.; Drugeon, H.; Dubreuil, L.; Jarlier, V.; Jehl, F. Comité de l’Antibiogramme de la Société Française de Microbiologie. Recommandations. 2013. Available online: https://resapath.anses.fr/resapath_uploadfiles/files/Documents/2013_CASFM.pdf (accessed on 13 April 2021).
- Tan, Y.E.; Chen, S.C.-A.; Halliday, C.L. Antimicrobial susceptibility profiles and species distribution of medically relevant Nocardia species: Results from a large tertiary laboratory in Australia. J. Glob. Antimicrob. Resist. 2020, 20, 110–117. [Google Scholar] [CrossRef]
- Navarrete-Navarrete, N.; Sevilla, J.E.; García, M.T.; Urbano, F.; Sabio, J.M.; Jiménez-Alonso, J. A Man with Unilateral Endophthalmitis: A Case of Disseminated Nocardiosis. Case Rep. Infect. Dis. 2015, 2015, 1–2. [Google Scholar] [CrossRef]
- Muricy, E.C.M.; Lemes, R.A.; Bombarda, S.; Ferrazoli, L.; Chimara, E. Differentiation Between Nocardia spp. and Mycobacterium spp.: Critical Aspects for Bacteriological Diagnosis. Revista Instituto Medicina Tropical São Paulo 2014, 56, 397–401. [Google Scholar] [CrossRef]
- Tan, C.-K.; Lai, C.-C.; Lin, S.-H.; Liao, C.-H.; Chou, C.-H.; Hsu, H.-L.; Huang, Y.-T.; Hsueh, P.-R. Clinical and microbiological characteristics of Nocardiosis including those caused by emerging Nocardia species in Taiwan, 1998–2008. Clin. Microbiol. Infect. 2010, 16, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Public Health Agency of Canada. Nocardia. Available online: http://www.phac-aspc.gc.ca/lab-bio/res/psds-ftss/nocardia-eng.php (accessed on 16 April 2020).
- Kurtböke, D.; Okazaki, T.; Vobis, G. Actinobacteria in Marine Environments: From Terrigenous Origin to Adapted Functional Diversity. In Encyclopedia of Marine Biotechnology; Kim, S.K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 1951–1978. [Google Scholar]
- Han, H.; Kwak, M.; Ha, S.; Yang, S.; Kim, J.D.; Cho, K.; Kim, T.; Cho, M.Y.; Kim, B.; Jung, S.; et al. Genomic characterization of Nocardia seriolae strains isolated from diseased fish. Microbiology 2018, 8, e00656. [Google Scholar] [CrossRef] [Green Version]
- Das, A.K.; Nandy, S.; Dudeja, M.; Tiwari, R.; Alam, S. The Incidence of Nocardiosis at Pulmonary and Extra-Pulmonary Sites. J. Clin. Diagn. Res. 2013, 7, 1427–1429. [Google Scholar] [CrossRef]
- Currie, B.J.; Carapetis, J.R. Skin infections and infestations in Aboriginal communities in northern Australia. Australas. J. Dermatol. 2000, 41, 139–143. [Google Scholar] [CrossRef]
- Leibniz Institute DSMZ—German Collection of Microorganisms and Cell Cultures. 2007. Available online: https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium65.pdf (accessed on 1 January 2020).
- Global Biodiversity Information Facility (GBIF). 2014. Available online: http://www.gbif-prokarya.de/microorganisms/wink.html (accessed on 14 January 2020).
- Gathogo, E.W.N.; Waugh, A.C.W.; Perić, N.; Redpath, M.B.; Long, P.F. Colony PCR amplification of actinomycete DNA. J. Antibiot. 2003, 56, 423–424. [Google Scholar] [CrossRef] [Green Version]
- Madueño, L.; Coppotelli, B.; Alvarez, H.; Morelli, I. Isolation and characterization of indigenous soil bacteria for bioaugmentation of PAH contaminated soil of semiarid Patagonia, Argentina. Int. Biodeterior. Biodegrad. 2011, 65, 345–351. [Google Scholar] [CrossRef]
- DNA Baser. Sequence Assembly Software, 2020. Available online: http://www.dnabaser.com (accessed on 8 March 2020).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information (NCBI). BLAST: Basic Local Alignment Tool. 2020. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 4 November 2020).
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Nettelbladt, C.G.; Katouli, M.; Bark, T.; Svenberg, T.; Möllby, R.; Ljungqvist, O. Evidence of Bacterial Translocation in Fatal Hemorrhagic Pancreatitis. J. Trauma Inj. Infect. Crit. Care 2000, 48, 314–315. [Google Scholar] [CrossRef]
- Katouli, M.; Nettebladt, C.G.; Muratov, V.; Ljungqvist, O.; Bark, T.; Svenberg, T.; Möllby, R. Selective translocation of coliform bacteria adhering to caecal epithelium of rats during catabolic stress. J. Med. Microbiol. 1997, 46, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Vollmerhausen, T.L.; Woods, J.L.; Faoagali, J.; Katouli, M. Interactions of uroseptic Escherichia coli with renal (A-498) and gastrointestinal (HT-29) cell lines. J. Med. Microbiol. 2014, 63, 1575–1583. [Google Scholar] [CrossRef]
- Dekali, S.; Gamez, C.; Kortulewski, T.; Blazy, K.; Rat, P.; Lacroix, G. Assessment of an in vitro model of pulmonary barrier to study the translocation of nanoparticles. Toxicol. Rep. 2014, 1, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Maher, S.; McClean, S. Investigation of the cytotoxicity of eukaryotic and prokaryotic antimicrobial peptides in intestinal epithelial cells in vitro. Biochem. Pharmacol. 2006, 71, 1289–1298. [Google Scholar] [CrossRef]
- Ramos, N.L.; Lamprokostopoulou, A.; Chapman, T.A.; Chin, J.C.; Römling, U.; Brauner, A.; Katouli, M. Virulence characteristics of translocating Escherichia coli and the interleukin-8 response to infection. Microb. Pathog. 2011, 50, 81–86. [Google Scholar] [CrossRef]
- Katouli, M.; Ramos, N.L.; Nettelbladt, C.G.; Ljungdahl, M.; Robinson, W.; Ison, H.M.; Brauner, A.; Möllby, R. Host species-specific translocation of Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1095–1103. [Google Scholar] [CrossRef]
- Kurtböke, D.I. Bioactive Actinomycetes: Reaching Rarity Through Sound Understanding of Selective Culture and Molecular Diversity. In Microbial Resources; Kurtböke, D.I., Ed.; Elsevier: London, UK, 2017; pp. 45–76. [Google Scholar]
- Bradley, S.; Anderson, D.; Jones, L. Phylogeny of actinomycetes as revealed by susceptibility to actinophage. J. Ind. Microbiol. 1961, 2, 223–237. [Google Scholar]
- Jonns, J.A.; Brooks, P.R.; Exley, P.; Poole, S.; Kurtböke, D.I. Streptophage-mediated control of off-flavour taint producing streptomycetes isolated from barramundi ponds. Synth. Syst. Biotechnol. 2017, 2, 105–112. [Google Scholar] [CrossRef]
- Porri, F.; Puccinelli, E.; Weidberg, N.; Pattrick, P. Lack of match between nutrient-enriched marine seafoam and intertidal abundance of long-lived invertebrate larvae. J. Sea Res. 2021, 170, 102009. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | % Resistance |
|---|---|
| Ampicillin (10 μg) | 15.7 (n = 5) |
| Ceftriaxone (30 μg) | 75 (n = 24) |
| Cefotaxime (30 μg) | 84.4 (n = 27) |
| Imipenem (10 μg) | 6.3 (n = 2) |
| Amikacin (30 μg) | − |
| Minocycline (30 μg) | 6.3 (n = 2) |
| Sulfamethoxazole/ trimethoprim (1.25/23.75 µg) | 50 (n = 16) * |
| Erythromycin (15 μg) | 6.3 (n = 2) |
| Tobramycin (10 μg) | 63 (n = 20) |
| Strain Code | Phage Susceptibility |
|---|---|
| USC-21042, USC-21037 | +++ |
| USC-21044, USC-21021, USC-21046 | ++ |
| USC-21018 | ± |
| USC-21034, USC-21050, USC-21049, USC-21035, USC-21030, USC-21025, USC-21006, USC-21038, USC-21039, USC-21036, USC-21048, USC-21017, USC-21028, USC-21027, USC-21010, USC-21029, USC-21047, USC-21026, USC-21022, USC-21040, USC-21043, USC-21043, USC-21016, USC-21012, USC-21011, USC-21024 | − |
| Phage Codes | Propagation Hosts Used to Isolate the Phages |
|---|---|
| Ø1 | Nocardia soli (DSMZ-44490) |
| Ø2 | Nocardia soli (DSMZ-44490) |
| Ø3 | Nocardia soli (DSMZ-44490) |
| Ø4 | Nocardia asteroides (ACM-2963) * |
| Ø5 | Nocardia soli (DSMZ-44490) |
| Ø6 | Nocardia asteroids (ACM-2963) * |
| Type Strain IDs | Phage Susceptibility |
|---|---|
| N. shimofusensis (DSMZ 44733) | − |
| N. takedensis (DSMZ 44802) | ± |
| N. cumidelens/soli (DSMZ 44488) | ++ |
| N. soli (DSMZ 44490) | +++ |
| N. uniformis (DSMZ 43136) | ++ |
| N. salmonicida (DSMZ 40472) | ++ |
| N. pseudovaccinii (DSMZ 43406) | ++ |
| N. veterana (DSMZ 44445) | ++ |
| N. fluminea (DSMZ 44489) | − |
| N. flavorosea (DSMZ 44480) | +++ |
| N. asteroides (ACM 131) | ++ |
| N. asteroides (ACM 2963) | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wright, L.; Katouli, M.; Kurtböke, D.İ. Isolation and Characterization of Nocardiae Associated with Foaming Coastal Marine Waters. Pathogens 2021, 10, 579. https://doi.org/10.3390/pathogens10050579
Wright L, Katouli M, Kurtböke Dİ. Isolation and Characterization of Nocardiae Associated with Foaming Coastal Marine Waters. Pathogens. 2021; 10(5):579. https://doi.org/10.3390/pathogens10050579
Chicago/Turabian StyleWright, Luke, Mohammad Katouli, and D. İpek Kurtböke. 2021. "Isolation and Characterization of Nocardiae Associated with Foaming Coastal Marine Waters" Pathogens 10, no. 5: 579. https://doi.org/10.3390/pathogens10050579
APA StyleWright, L., Katouli, M., & Kurtböke, D. İ. (2021). Isolation and Characterization of Nocardiae Associated with Foaming Coastal Marine Waters. Pathogens, 10(5), 579. https://doi.org/10.3390/pathogens10050579