
Morphological and Phylogenetic Analysis of Eustrongylides sp. and Gnathostoma spinigerum Parasitizing the Asian Swamp Eel Monopterusalbus in China


 ,
, 
Abstract
:1. Introduction
2. Results
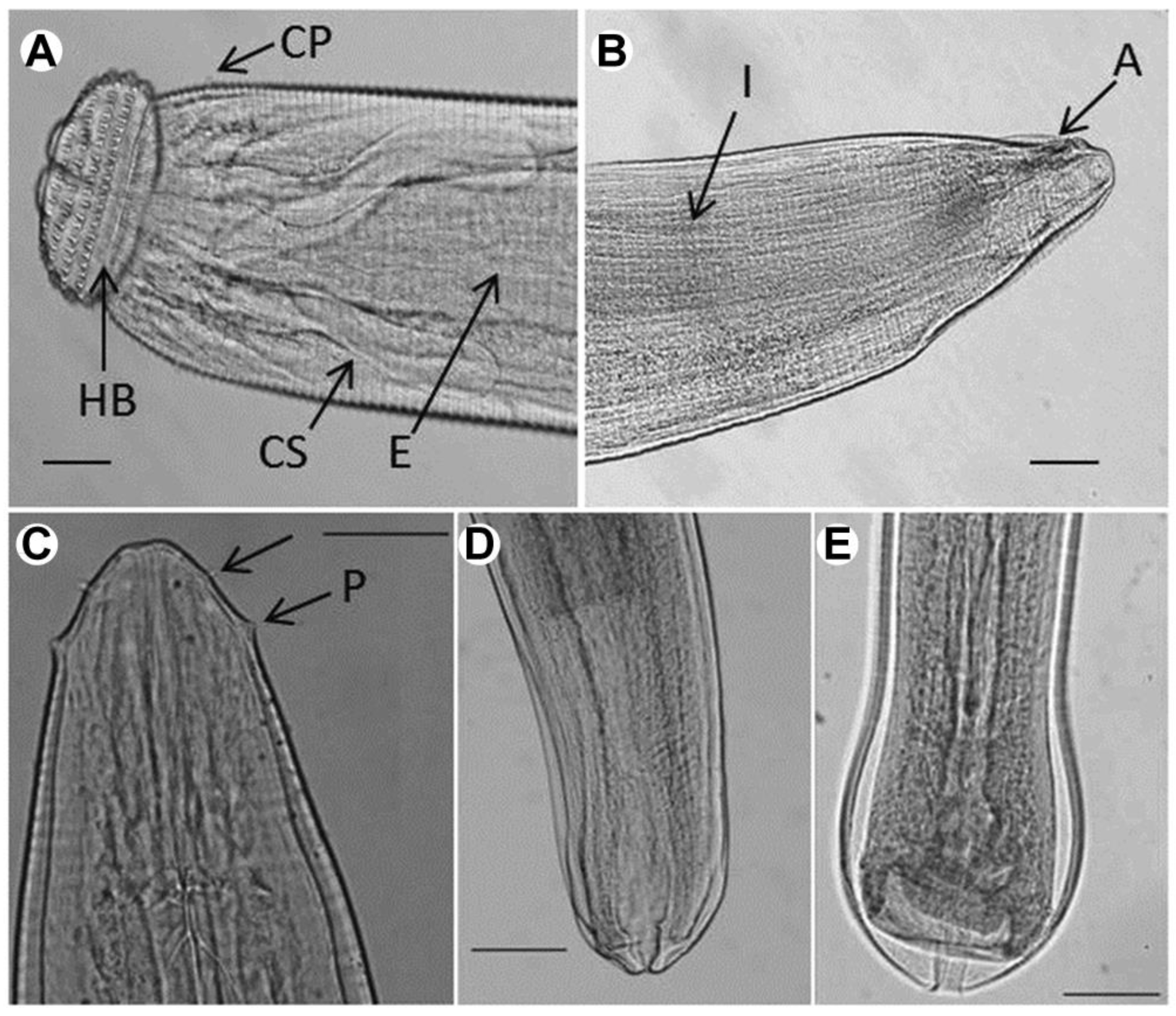
2.1. Morphological Characterization
2.1.1. Gnathostoma spinigerum
2.1.2. Eustrongylides sp.
2.2. ITS and COI Amplification and Sequencing
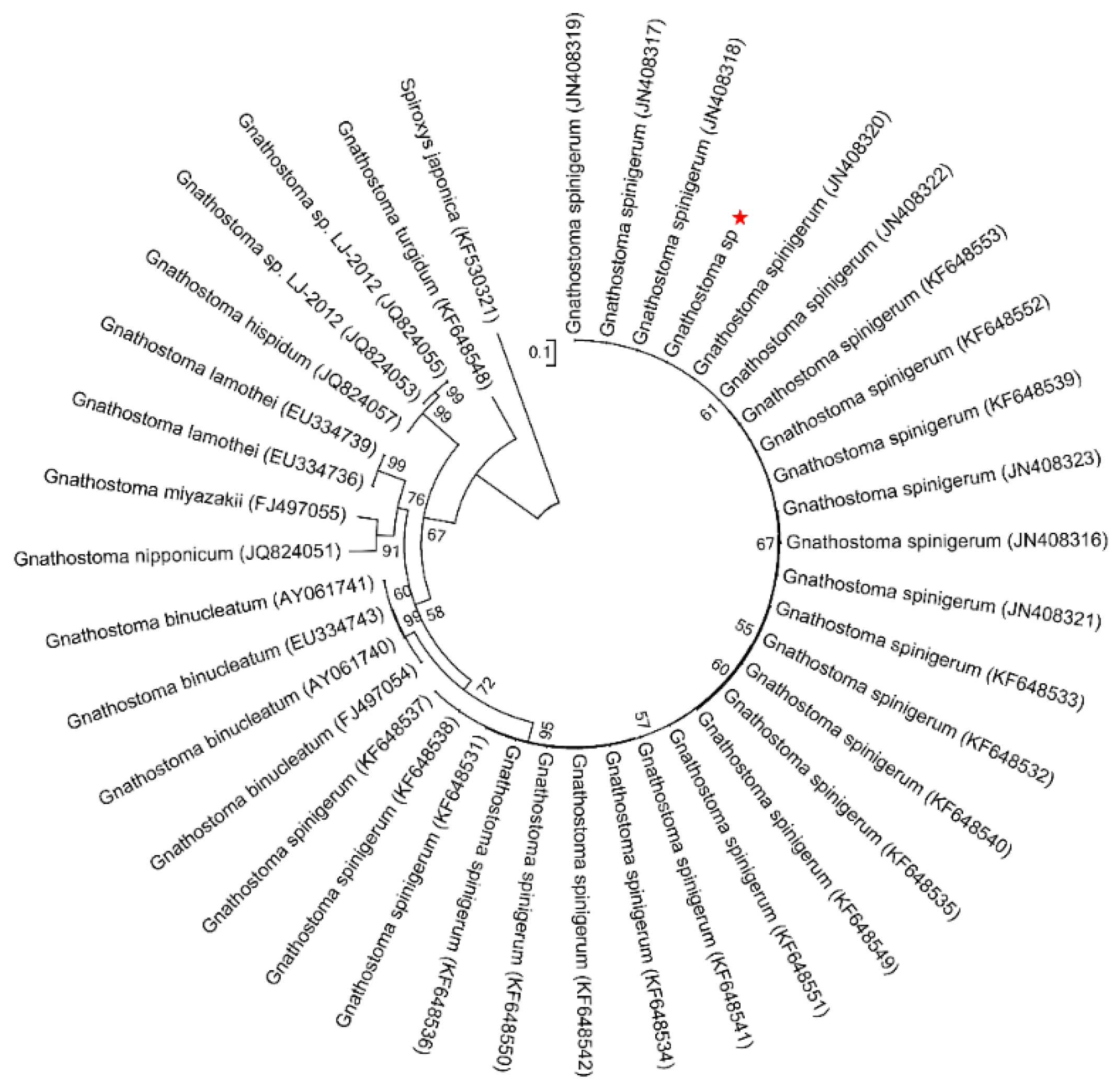
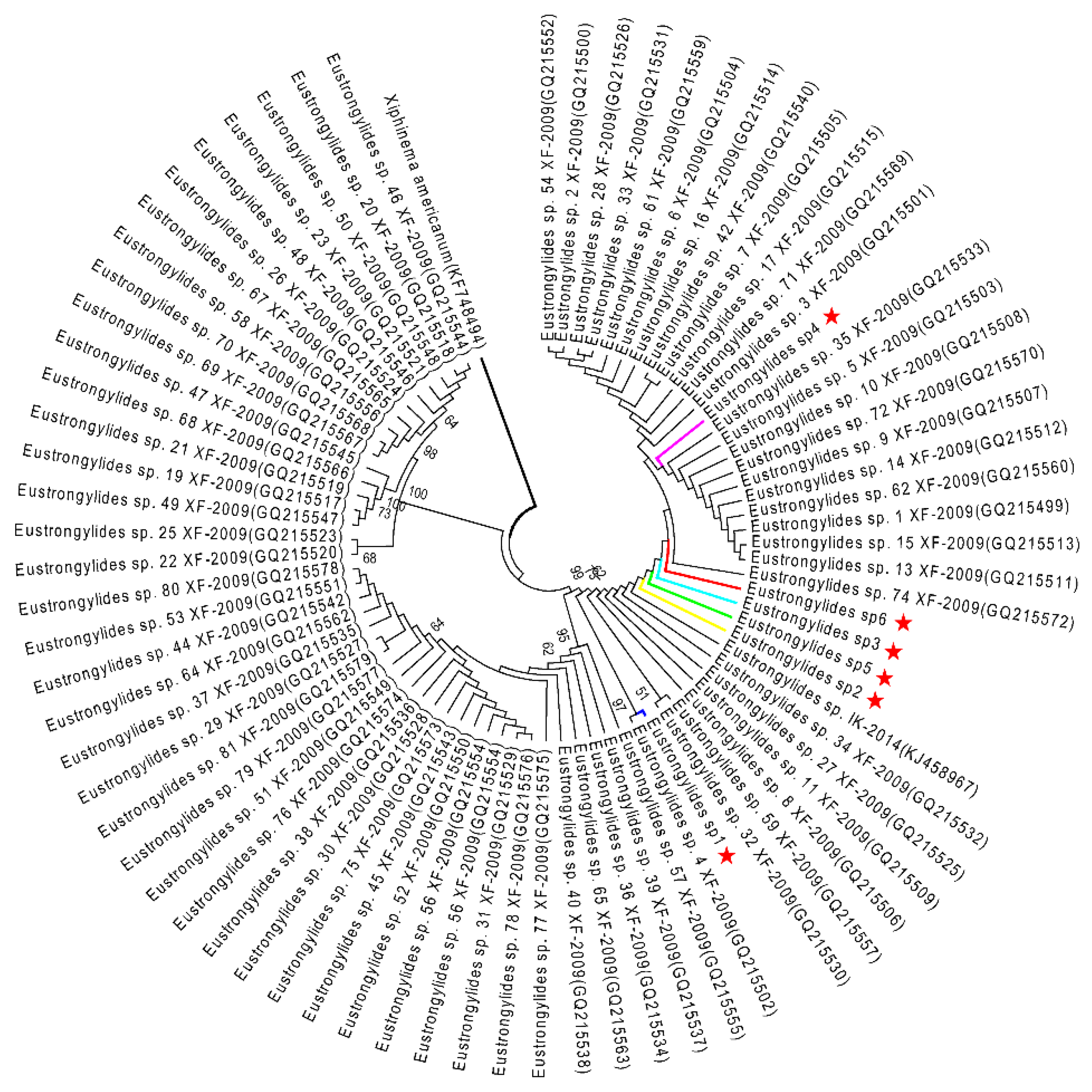
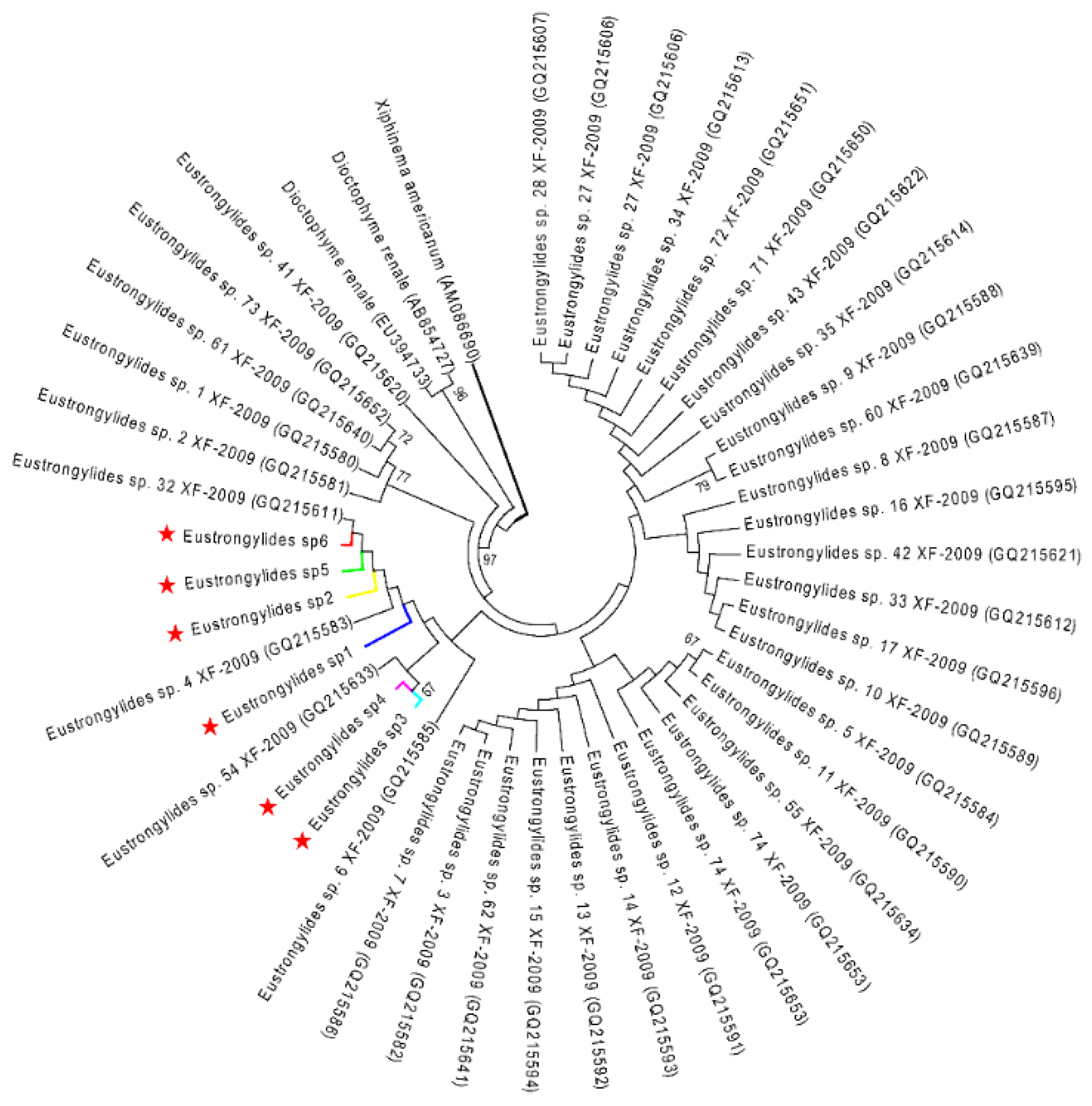
2.3. Phylogenetic Analysis
2.4. Ecology of Eustrongylides sp. and Gnathostoma Spinigerum in the Asian Swamp Eel
3. Discussion
4. Materials and Methods
4.1. Parasites Collection
4.2. Morphological Observation
4.3. DNA Extraction and Molecular Identification
4.4. Phylogenetic Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Froese, R.; Pauly, D. (Eds.) FishBase. Available online: https://www.fishbase.se/summary/Monopterus-albus.html (accessed on 7 January 2021).
- Cole, R.A.; Choudhury, A.; Nico, L.G.; Griffin, K.M. Gnathostoma spinigerum in live Asian swamp eels (Monopterus spp.) from food markets and wild populations, United States. Emerg. Infect. Dis. 2014, 20, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Measures, L.N. Epizootiology, Pathology, and Description of Eustrongylides-Tubifex (Nematoda, Dioctophymatoidea) in Fish. Can. J. Zool. 1988, 66, 2212–2222. [Google Scholar] [CrossRef]
- Chen, Q.Q.; Lin, X.M. A survey of epidemiology of Gnathostoma hispidum and experimental studies of its larvae in animals. Southeast Asian J. Trop. Med. Public Health 1991, 22, 611–617. [Google Scholar] [PubMed]
- Coyner, D.F.; Spalding, M.G.; Forrester, D.J. Epizootiology of Eustrongylides ignotus in Florida: Transmission and development of larvae in intermediate hosts. J. Parasitol. 2003, 89, 290–298. [Google Scholar] [CrossRef]
- Alvarez-Guerrero, C.; Muñoz-Guzmán, M.A.; Buendía-Jiménez, J.A.; Alba-Hurtado, F. Gnathostoma binucleatum: Pathological and parasitological aspects in experimentally infected dogs. Exp. Parasitol. 2011, 127, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Bertoni-Ruiz, F.; Argumedo, M.R.L.Y.; Garcia-Prieto, L.; Osorio-Sarabia, D.; Leon-Regagnon, V. Systematics of the genus Gnathostoma (Nematoda: Gnathostomatidae) in the Americas. Rev. Mex. Biodivers. 2011, 82, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Defendi, G.L. Gnathostomiasis. Available online: http://emedicine.medscape.com/article/998278-overview (accessed on 17 January 2021).
- Wai, A.P.; Sripan, P.; Boonmars, T.; Sriraj, P.; Aukkanimart, R.; Songsri, J.; Boueroy, P.; Boonjaraspinyo, S.; Khueangchaingkhwang, S.; Laummaunwai, P.; et al. Seasonal variation of parasitic infections in Asian swamp eels (Monopterus albus) from local markets in Yangon, Myanmar. Vet. Integr. Sci. 2019, 17, 181–193. [Google Scholar]
- Sohn, W.M.; Kho, W.G.; Lee, S.H. Larval Gnathostoma nipponicum found in the imported Chinese loaches. Korean J. Parasitol. 1993, 31, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Haugen, P.; Hemmingsen, W.; MacKenzie, K. The distribution of Eustrongylides sp (Nematoda: Dioctophymatoidea) in brown trout Salmo trutta L. in the River Otra in southern Norway. Bull. Eur. Assoc. Fish Pathol. 2008, 28, 138–143. [Google Scholar]
- Hernández-Gómez, R.E.; Martínez-Salazar, E.A.; López-Jiménez, S.; León-Règagnon, V. Molecular identification of the advanced third-stage larvae (ADV L(3)) of Gnathostoma lamothei in Tabasco, Mexico. Parasitol. Int. 2010, 59, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Wittner, M.; Turner, J.W.; Jacquette, G.; Ash, L.R.; Salgo, M.P.; Tanowitz, H.B. Eustrongylidiasis—A parasitic infection acquired by eating sushi. N. Engl. J. Med. 1989, 320, 1124–1126. [Google Scholar] [CrossRef]
- Narr, L.L.; O’Donnell, J.G.; Libster, B.; Alessi, P.; Abraham, D. Eustrongylidiasis—A parasitic infection acquired by eating live minnows. J. Am. Osteopath. Assoc. 1996, 96, 400–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirikulchayanonta, V.; Viriyavejakul, P. Various morphologic features of Gnathostoma spinigerum in histologic sections: Report of 3 cases with reference to topographic study of the reference worm. Southeast Asian J. Trop. Med. Public Health 2001, 32, 302–307. [Google Scholar] [PubMed]
- Tiwari, S.; Chayani, N.; Rautaraya, B. Intraocular Gnathostoma spinigerum: A case report. Cases J. 2009, 2, 9370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussaratid, V.; Dekumyoy, P.; Desakorn, V.; Jaroensuk, N.; Liebtawee, B.; Pakdee, W.; Wattanagoon, Y. Predictive factors for Gnathostoma seropositivity in patients visiting the Gnathostomiasis Clinic at the Hospital for Tropical Diseases, Thailand during 2000–2005. Southeast Asian J. Trop. Med. Public Health 2010, 41, 1316–1321. [Google Scholar] [PubMed]
- Kim, J.H.; Lim, H.; Hwang, Y.S.; Kim, T.Y.; Han, E.M.; Shin, E.H.; Chai, J.Y. Gnathostoma spinigerum infection in the upper lip of a Korean woman: An autochthonous case in Korea. Korean J. Parasitol. 2013, 51, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wang, Y.; Wang, Z.Q. Cutaneous gnathostomiasis with recurrent migratory nodule and persistent eosinophilia: A case report from China. Korean J. Parasitol. 2013, 51, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Ruiz-Tiben, E. Cutaneous emergence of Eustrongylides in two persons from South Sudan. Am. J. Trop. Med. Hyg. 2014, 90, 315–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chotmongkol, V.; Kitkhuandee, A.; Sawanyawisuth, K. Spinal epidural hematoma and gnathostomiasis. Am. J. Trop. Med. Hyg. 2015, 92, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, J.H. Gnathostomiasis: An Emerging Infection of Raw Fish Consumers in Gnathostoma Nematode-Endemic and Nonendemic Countries. J. Travel Med. 2015, 22, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.H.; Sun, M.M.; Elsheikha, H.M.; Fu, Y.T.; Sugiyama, H.; Ando, K.; Sohn, W.M.; Zhu, X.Q.; Yao, C. Human gnathostomiasis: A neglected food-borne zoonosis. Parasites Vectors 2020, 13, 616. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, A.; Caffara, M.; Gustinelli, A.; Agnetti, F.; Sgariglia, E.; Lo Vaglio, G.; Quaglio, F.; Fioravanti, M.L. Morphological and Molecular Characterization of Larval and Adult Stages of Eustrongylides excisus (Nematoda: Dioctophymatoidea) with Histopathological Observations. J. Parasitol. 2019, 105, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Menconi, V.; Riina, M.V.; Pastorino, P.; Mugetti, D.; Canola, S.; Pizzul, E.; Bona, M.C.; Dondo, A.; Acutis, P.L.; Prearo, M. First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range. Int. J. Environ. Res. Public Health 2020, 17, 4171. [Google Scholar] [CrossRef] [PubMed]
- Novakov, N.; Bielic-Cabrilo, O.; Cirkovic, M.; Jubojevic, D.; Lujic, J.; Davidov, I.; Jovanovic, M. Eustrongylidosis of European catfish (Siluris glanis). Bulg. J. Agric. Sci. 2013, 19, 72–76. [Google Scholar]
- Gasser, R.B.; Hoste, H. Genetic markers for closely-related parasitic nematodes. Mol. Cell. Probes 1995, 9, 315–320. [Google Scholar] [CrossRef]
- Almeyda-Artigas, R.J.; Bargues, M.D.; Mas-Coma, S. rDNA of Gnathostoma species (Nematoda): ITS-2 microsatellites and 5.8S gene secondary structure. Res. Rev. Parasitol. 2000, 60, 51–56. [Google Scholar]
- Ngarmamonpirat, C.; Waikagul, J.; Petmitr, S.; Dekumyoy, P.; Rojekittikhun, W.; Anantapruti, M.T. Analysis of sequence variation in Gnathostoma spinigerum mitochondrial DNA by single-strand conformation polymorphism analysis and DNA sequence. Parasitol. Int. 2005, 54, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Wang, G.T.; Wu, S.G.; Nie, P. Development of Eustrongylides ignotus (Nematoda: Dioctophmida) in domestic ducks (Anas platyrhynchos domestica (L.)). J. Parasitol. 2009, 95, 1035–1039. [Google Scholar] [CrossRef]
- Nabavi, R.; Conneely, B.; McCarthy, E.; Good, B.; Shayan, P.; De-Waal, T. Comparison of internal transcribed spacers and intergenic spacer regions of five common Iranian sheep bursate nematodes. Iran. J. Parasitol. 2014, 9, 350–357. [Google Scholar] [PubMed]
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Diaz Camacho, S.P.; Willms, K.; Ramos, M.Z.; del Carmen de la Cruz Otero, M.; Nawa, Y.; Akahane, H. Morphology of Gnathostoma spp. isolated from natural hosts in Sinaloa, Mexico. Parasitol. Res. 2002, 88, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Li, W.X.; Wu, S.G.; Zou, H.; Wang, G.T. Molecular phylogeny and host specificity of the larval Eustrongylides (Nematoda: Dioctophmidae) from freshwater fish in China. J. Parasitol. 2013, 99, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, C.S.; Müller, G. Third-stage larvae of the enoplid nematode Dioctophyme renale (Goeze, 1782) in the freshwater turtle Trachemys dorbigni from southern Brazil. J. Helminthol. 2015, 89, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Q.; Gasser, R.B.; Podolska, M.; Chilton, N.B. Characterisation of anisakid nematodes with zoonotic potential by nuclear ribosomal DNA sequences. Int. J. Parasitol. 1998, 28, 1911–1921. [Google Scholar] [CrossRef]
- Vogler, A.P.; Monaghan, M.T. Recent advances in DNA taxonomy. J. Zool. Syst. Evol. Res. 2007, 45, 1–10. [Google Scholar] [CrossRef]
- Marucci, G.; Interisano, M.; La Rosa, G.; Pozio, E. Molecular identification of nematode larvae different from those of the Trichinella genus detected by muscle digestion. Vet. Parasitol. 2013, 194, 117–120. [Google Scholar] [CrossRef]
- Sieu, T.P.; Dung, T.T.; Nga, N.T.; Hien, T.V.; Dalsgaard, A.; Waikagul, J.; Murrell, K.D. Prevalence of Gnathostoma spinigerum infection in wild and cultured swamp eels in Vietnam. J. Parasitol. 2009, 95, 246–248. [Google Scholar] [CrossRef]
- Sujata, D.N.; Renu, B.S. Intraocular gnathostomiasis from coastal part of Maharashtra. Trop. Parasitol. 2013, 3, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Rojekittikhun, W.; Chaiyasith, T.; Nuamtanong, S.; Pubampen, S.; Maipanich, W.; Tungtrongchitr, R. Gnathostoma infection in Nakhon Nayok and Prachin Buri, Central Thailand. Southeast Asian J. Trop Med. Public Health 2002, 33, 474–484. [Google Scholar] [PubMed]
- Saksirisampant, W.; Thanomsub, B.W. Positivity and intensity of Gnathostoma spinigerum infective larvae in farmed and wild-caught swamp eels in Thailand. Korean J. Parasitol. 2012, 50, 113–118. [Google Scholar] [CrossRef]
- Jackson, G.J.; Bier, J.W.; Payne, W.L.; McClure, F.D. Recovery of parasitic nematodes from fish by digestion or elution. Appl. Environ. Microbiol. 1981, 41, 912–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.M.; Chen, X.R.; Zhou, J.S.; Xu, Z.B.; Nawa, Y.; Dekumyoy, P. Short report: Case of gnathostomiasis in Beijing, China. Am. J. Trop. Med. Hyg. 2009, 80, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Watanobe, T.; Liu, C.X.; Init, I.; Blair, D.; Ohnishi, S.; Agatsuma, T. Mitochondrial DNA and nuclear DNA indicate that the Japanese Fasciola species is F. gigantica. Parasitol. Res. 1997, 83, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonroumkaew, P.; Sanpool, O.; Rodpai, R.; Sadaow, L.; Somboonpatarakun, C.; Laymanivong, S.; Aung, W.P.P.; Un, M.; Laummaunwai, P.; Intapan, P.M.; et al. Molecular identification and genetic diversity of Gnathostoma spinigerum larvae in freshwater fishes in southern Lao PDR, Cambodia, and Myanmar. Parasitol. Res. 2019, 118, 1465–1472. [Google Scholar] [CrossRef]
- Eamsobhana, P.; Wanachiwanawin, D.; Roongruangchai, K.; Song, S.L.; Yong, H.S. Genetic diversity of infective larvae of Gnathostoma spinigerum (Nematoda: Gnathostomatidae) in freshwater swamp eels from Thailand. J. Helminthol. 2017, 91, 767–771. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic * | AL3 (n = 6) |
|---|---|
| Body length | 3.632 ± 0.594 |
| Maximum width | 0.395 ± 0.019 |
| Length of cephalic bulb | 0.108 ± 0.018 |
| Width of cephalic bulb | 0.209 ± 0.019 |
| Esophagus | 1.120 ± 0.028 |
| Cervical papillae to anterior end | 0.350 ± 0.055 |
| Length of cervical sacs | 0.451 ± 0.028 |
| Anus to posterior end | 0.062 ± 0.014 |
| Tail | 0.037 |
| Characteristic * | Males (n = 6) | Females (n = 6) |
|---|---|---|
| Body length | 55.450 ± 3.136 | 57.675 ± 3.172 |
| Max body width | 0.419 ± 0.032 | 0.502 ± 0.048 |
| Internal circle of papillae to anterior end | 0.022 ± 0.015 | 0.022 ± 0.018 |
| External circle of papillae to anterior end | 0.062 ± 0.017 | 0.066 ± 0.012 |
| Nerve ring to anterior end | 0.313 ± 0.035 | 0.325 ± 0.059 |
| Esophagus | 14.091 ± 1.523 | 14.311 ± 1.152 |
| Esophagus/body length (%) | 25.408 ± 1.445 | 25.816 ± 1.488 |
| Tail | 0.236 ± 0.074 | 0.250 ± 0.062 |
| Vulvar primordium to posterior end | – | 6.150 ± 0.680 |
| Species | Gene | Primers (5–3′) | Annealing Temperature | Target Size (bp) | |
|---|---|---|---|---|---|
| Eustrongylides spp.* | ITS | F R F R | TGGATGATTCGGTGAGGT | 55 ℃ | 900 |
| AACCGCTTAGTAATATGCT | |||||
| COI | ACNACRTARTANGTRTCRTG | 55 ℃ | 419 | ||
| TGRTTYTTYGGNCAYCC | |||||
| Gnathostoma spp.†‡ | ITS | F R F R | TGTGTCGATGAAGAACGCAG | 55 ℃ | 568 |
| TTCTATGCTTAAATTCAGGGG | |||||
| COI | TTTTGGGCATCCTGAGGTTTAT | 55 ℃ | 726 | ||
| AAAGAAAGAACATAATGAAAA | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Huang, G.; Li, L.; Liu, X.; Tang, X.; Suo, X. Morphological and Phylogenetic Analysis of Eustrongylides sp. and Gnathostoma spinigerum Parasitizing the Asian Swamp Eel Monopterusalbus in China. Pathogens 2021, 10, 711. https://doi.org/10.3390/pathogens10060711
Zhang S, Huang G, Li L, Liu X, Tang X, Suo X. Morphological and Phylogenetic Analysis of Eustrongylides sp. and Gnathostoma spinigerum Parasitizing the Asian Swamp Eel Monopterusalbus in China. Pathogens. 2021; 10(6):711. https://doi.org/10.3390/pathogens10060711
Chicago/Turabian StyleZhang, Sixin, Guangping Huang, Liang Li, Xianyong Liu, Xiaoli Tang, and Xun Suo. 2021. "Morphological and Phylogenetic Analysis of Eustrongylides sp. and Gnathostoma spinigerum Parasitizing the Asian Swamp Eel Monopterusalbus in China" Pathogens 10, no. 6: 711. https://doi.org/10.3390/pathogens10060711
APA StyleZhang, S., Huang, G., Li, L., Liu, X., Tang, X., & Suo, X. (2021). Morphological and Phylogenetic Analysis of Eustrongylides sp. and Gnathostoma spinigerum Parasitizing the Asian Swamp Eel Monopterusalbus in China. Pathogens, 10(6), 711. https://doi.org/10.3390/pathogens10060711