
Regulation of hBD-2, hBD-3, hCAP18/LL37, and Proinflammatory Cytokine Secretion by Human Milk Oligosaccharides in an Organotypic Oral Mucosal Model

 , , ,
, , ,
Abstract
:1. Introduction
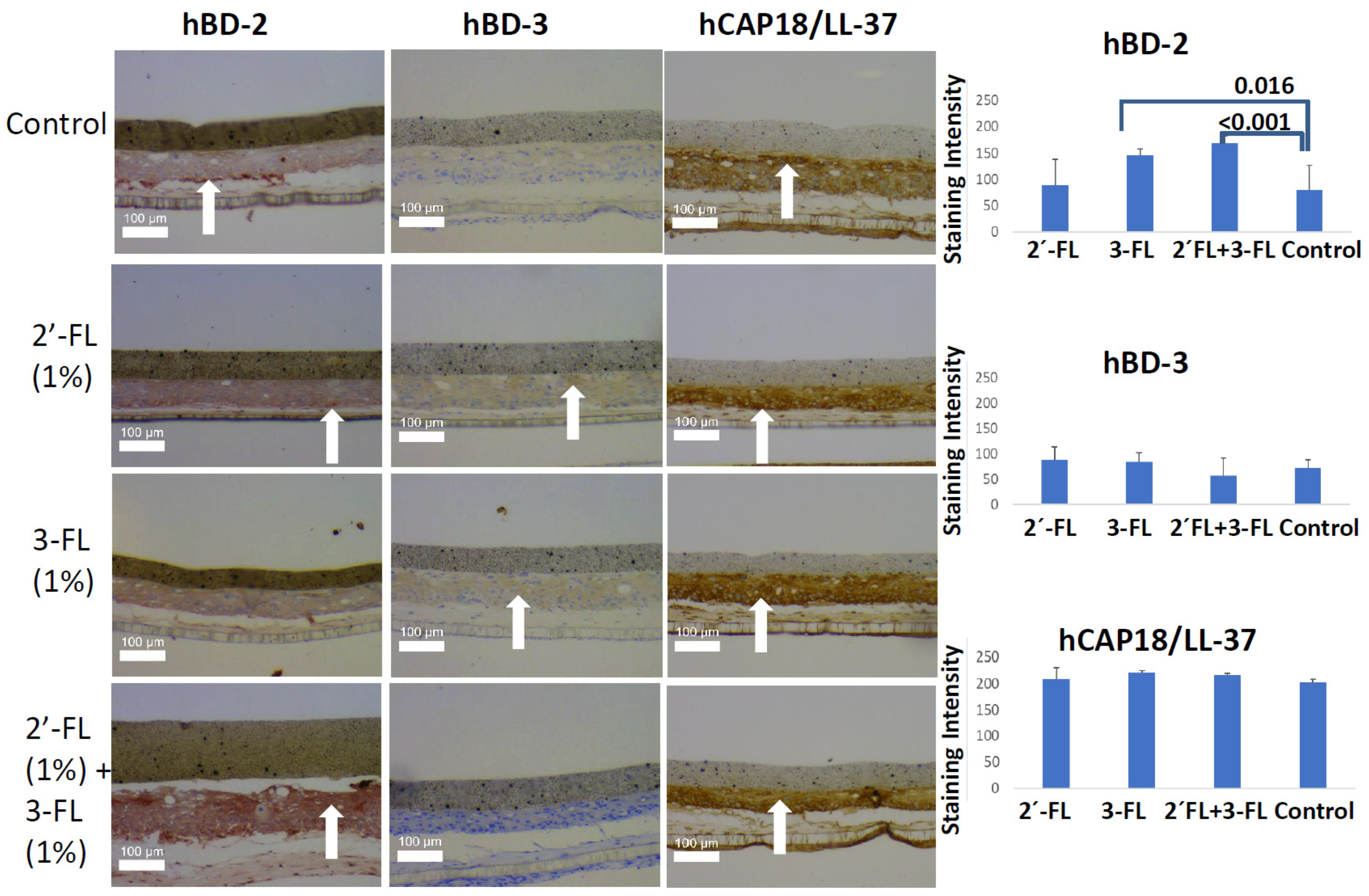
2. Results
2.1. Immunohistochemical Analysis of an Organotypic Oral Mucosa Model
2.2. Proinflammatory Cytokine Analysis in Culture Medium
3. Discussion
4. Materials and Methods
4.1. 2′-Fucosyllactose and 3-Fucosyllactose Production and Purity
4.2. Construction of Organotypic Oral Mucosal Model
4.3. Stimulation of Organotypic Oral Mucosal Model and Sample Collection
4.4. Immunohistochemical Examination of Organotypic Oral Mucosal Model
4.5. Proinflammatory Cytokine Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Urashima, T.; Hirabayashi, J.; Sato, S.; Kobata, A. Human Milk Oligosaccharides as Essential Tools for Basic and Application Studies on Galectins. Trends Glycosci. Glycotechnol. 2018, 30, SE51–SE65. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, O.; Zampini, L.; Galeazzi, T.; Padella, L.; Santoro, L.; Peila, C.; Giuliani, F.; Bertino, E.; Fabris, C.; Coppa, G.V. Preterm milk oligosaccharides during the first month of lactation. Pediatrics 2011, 128, e1520–e1531. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S.; Baier, W.; Klein, N.; Strobel, S. Oligosaccharides in human milk: Structural, functional, and metabolic aspects. Annu. Rev. Nutr. 2000, 20, 699–722. [Google Scholar] [CrossRef] [PubMed]
- Thurl, S.; Munzert, M.; Boehm, G.; Matthews, C.; Stahl, B. Systematic review of the concentrations of oligosaccharides in human milk. Nutr. Rev. 2017, 75, 920–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Könönen, E. Understanding the roles of gingival beta-defensins. J. Oral. Microbiol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Sechet, E.; Telford, E.; Bonamy, C.; Sansonetti, P.J.; Sperandio, B. Natural molecules induce and synergize to boost expression of the human antimicrobial peptide β-defensin-3. Proc. Natl. Acad. Sci. USA 2018, 115, E9869–E9878. [Google Scholar] [CrossRef] [Green Version]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Lawlor, N.T.; Newburg, D.S. Human Milk Components Modulate Toll-Like Receptor-Mediated Inflammation. Adv. Nutr. 2016, 7, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Vora, P.; Youdim, A.; Thomas, L.S.; Fukata, M.; Tesfay, S.Y.; Lukasek, K.; Michelsen, K.S.; Wada, A.; Hirayama, T.; Arditi, M.; et al. Beta-defensin-2 expression is regulated by TLR signaling in intestinal epithelial cells. J. Immunol. 2004, 173, 5398–5405. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.; Cheng, L.; Krenning, G.; Fledderus, J.; de Haan, B.J.; Walvoort, M.T.C.; de Vos, P. Human Milk Oligosaccharides Mediate the Crosstalk Between Intestinal Epithelial Caco-2 Cells and Lactobacillus Plantarum WCFS1in an In Vitro Model with Intestinal Peristaltic Shear Force. J. Nutr. 2020, 150, 2077–2088. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.R.; Rho, H.K.; Kessler, S.P.; Amin, R.; Homer, C.R.; McDonald, C.; Cowman, M.K.; de la Motte, C.A. Human milk hyaluronan enhances innate defense of the intestinal epithelium. J. Biol. Chem. 2013, 288, 29090–29104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, L.E.; Virmani, M.D.; Rosa, F.; Munblit, D.; Matazel, K.S.; Elolimy, A.A.; Yeruva, L. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front. Immunol. 2021, 12, 604080. [Google Scholar] [CrossRef]
- Thai, J.D.; Gregory, K.E. Bioactive Factors in Human Breast Milk Attenuate Intestinal Inflammation during Early Life. Nutrients 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cederlund, A.; Kai-Larsen, Y.; Printz, G.; Yoshio, H.; Alvelius, G.; Lagercrantz, H.; Strömberg, R.; Jörnvall, H.; Gudmundsson, G.H.; Agerberth, B. Lactose in human breast milk an inducer of innate immunity with implications for a role in intestinal homeostasis. PLoS ONE 2013, 8, e53876. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, S.; Kling, D.E.; Leone, S.; Lawlor, N.T.; Huang, Y.; Feinberg, S.B.; Hill, D.R.; Newburg, D.S. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016, 65, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koning, N.; Kessen, S.F.M.; Van Der Voorn, J.P.; Appelmelk, B.J.; Jeurink, P.V.; Knippels, L.M.J.; Garssen, J.; Van Kooyk, Y. Human Milk Blocks DC-SIGN-Pathogen Interaction via MUC1. Front. Immunol. 2015, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; van’t Land, B. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front. Pediatr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Ruben, M.P.; Frankl, S.N.; Wallace, S. The Histopathology of Periodontal Disease in Children. J. Periodontol. 1971, 42, 473–484. [Google Scholar] [CrossRef]
- Özdemir, M.; Caglayan, F.; Bikker, F.J.; Pussinen, P.; Könönen, E.; Yamalik, N.; Gürsoy, M.; Fteita, D.; Nazmi, K.; Güncü, G.N.; et al. Gingival tissue human beta-defensin levels in relation to infection and inflammation. J. Clin. Periodontol. 2020, 47, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hickey, R.M. The role of oligosaccharides from human milk and other sources in prevention of pathogen adhesion. Int. Dairy J. 2012, 22, 141–146. [Google Scholar] [CrossRef]
- Salli, K.; Söderling, E.; Hirvonen, J.; Gürsoy, U.K.; Ouwehand, A.C. Influence of 2′-fucosyllactose and galacto-oligosaccharides on the growth and adhesion of Streptococcus mutans. Br. J. Nutr. 2020, 124, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Weichert, S.; Jennewein, S.; Hüfner, E.; Weiss, C.; Borkowski, J.; Putze, J.; Schroten, H. Bioengineered 2′-fucosyllactose and 3-fucosyllactose inhibit the adhesion of Pseudomonas aeruginosa and enteric pathogens to human intestinal and respiratory cell lines. Nutr. Res. 2013, 33, 831–838. [Google Scholar] [CrossRef]
- Oksanen, J.; Hormia, M. An organotypic in vitro model that mimics the dento-epithelial junction. J. Periodontol. 2002, 73, 86–93. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Pöllänen, M.; Könönen, E.; Uitto, V.J. Biofilm formation enhances the oxygen tolerance and invasiveness of Fusobacterium nucleatum in an oral mucosa culture model. J. Periodontol. 2010, 81, 1084–1091. [Google Scholar] [CrossRef]
- Mäkelä, M.; Larjava, H.; Pirilä, E.; Maisi, P.; Salo, T.; Sorsa, T.; Uitto, V.J. Matrix metalloproteinase 2 (gelatinase A) is related to migration of keratinocytes. Exp. Cell Res. 1999, 251, 67–78. [Google Scholar] [CrossRef]


{kind=link}
| IL-1β (pg/mL) | IL-1RA (pg/mL) | IL-8 (pg/mL) | MCP-1 (pg/mL) | |
|---|---|---|---|---|
| 5 h | ||||
| Control | 0.9 ± 0.5 | 1688 ± 847 | 2,70,293 ± 120527 | 2218 ± 303 |
| 1% 2′-FL | 0.8 ± 0.3 | 619 ± 24 | 1,76,987 ± 83948 | 2253 ± 359 |
| 1% 3-FL | 1.2 ± 0.3 | 1830 ± 161 | 2,99,054 ± 234387 | 2410 ± 618 |
| 1% 2′-FL+3-FL | 0.8 ± 0.2 | 848 ± 361 | 2,05,258 ± 94,475 | 1911 ± 260 |
| 24 h | ||||
| Control | 0.7 ± 0.2 | 1959 ± 167 | 1,14,840 ± 58,017 | 1832 ± 232 |
| 1% 2′-FL | 0.6 ± 0.4 | 1736 ± 435 | 1,11,945 ± 94,296 | 1763 ± 371 |
| 1% 3-FL | 0.9 ± 0.5 | 1877 ± 246 | 1,11,233 ± 37,557 | 1693 ± 74 |
| 1% 2′-FL+3-FL | 0.5 ± 0.3 | 1698 ± 113 | 1,37,496 ± 78,958 | 1758 ± 229 |
| 24 h | ||||
| Control | 0.4 ± 0.1 | 572 ± 94 | 70,376 ± 15,278 | 1570 ± 88 |
| 0.1% 2′-FL | 1.2 ± 0.1 | 1542 ± 183 | 1,11,861 ± 36,202 | 1940 ± 504 |
| 0.1% 3-FL | 1.1 ± 0.7 | 1206 ± 1035 | 15,9765 ± 1,23,521 | 1759 ± 244 |
| 0.1% 2′-FL+3-FL | 1.0 ± 0.5 | 837 ± 618 | 1,46,936 ± 81,918 | 1782 ± 227 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gürsoy, U.K.; Salli, K.; Söderling, E.; Gürsoy, M.; Hirvonen, J.; Ouwehand, A.C. Regulation of hBD-2, hBD-3, hCAP18/LL37, and Proinflammatory Cytokine Secretion by Human Milk Oligosaccharides in an Organotypic Oral Mucosal Model. Pathogens 2021, 10, 739. https://doi.org/10.3390/pathogens10060739
Gürsoy UK, Salli K, Söderling E, Gürsoy M, Hirvonen J, Ouwehand AC. Regulation of hBD-2, hBD-3, hCAP18/LL37, and Proinflammatory Cytokine Secretion by Human Milk Oligosaccharides in an Organotypic Oral Mucosal Model. Pathogens. 2021; 10(6):739. https://doi.org/10.3390/pathogens10060739
Chicago/Turabian StyleGürsoy, Ulvi K., Krista Salli, Eva Söderling, Mervi Gürsoy, Johanna Hirvonen, and Arthur C. Ouwehand. 2021. "Regulation of hBD-2, hBD-3, hCAP18/LL37, and Proinflammatory Cytokine Secretion by Human Milk Oligosaccharides in an Organotypic Oral Mucosal Model" Pathogens 10, no. 6: 739. https://doi.org/10.3390/pathogens10060739
APA StyleGürsoy, U. K., Salli, K., Söderling, E., Gürsoy, M., Hirvonen, J., & Ouwehand, A. C. (2021). Regulation of hBD-2, hBD-3, hCAP18/LL37, and Proinflammatory Cytokine Secretion by Human Milk Oligosaccharides in an Organotypic Oral Mucosal Model. Pathogens, 10(6), 739. https://doi.org/10.3390/pathogens10060739