
Four Tick-Borne Microorganisms and Their Prevalence in Hyalomma Ticks Collected from Livestock in United Arab Emirates




Abstract
:1. Introduction
2. Results
2.1. Tick Identification
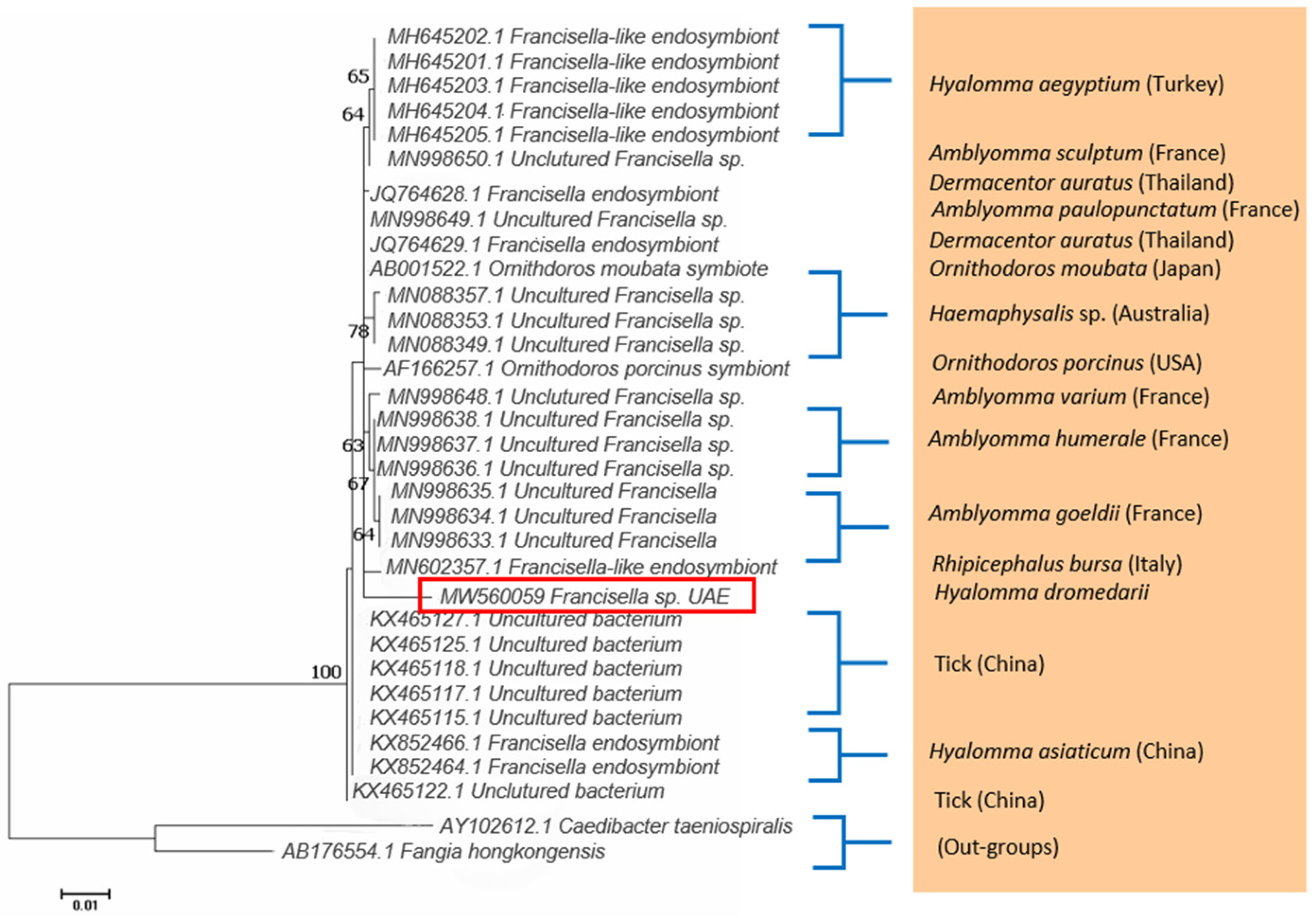
2.2. Detection of Francisella
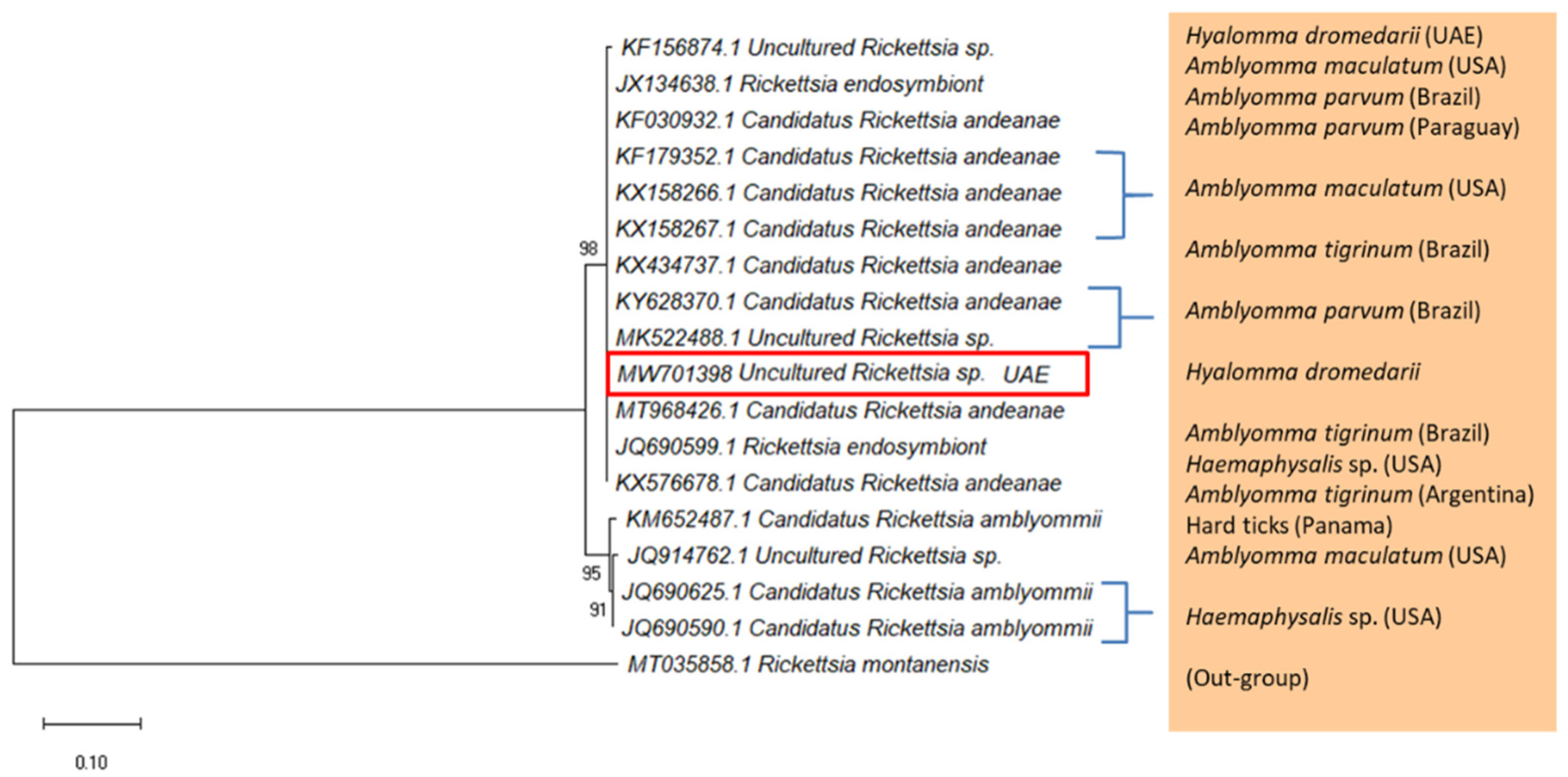
2.3. Detection of Rickettsia
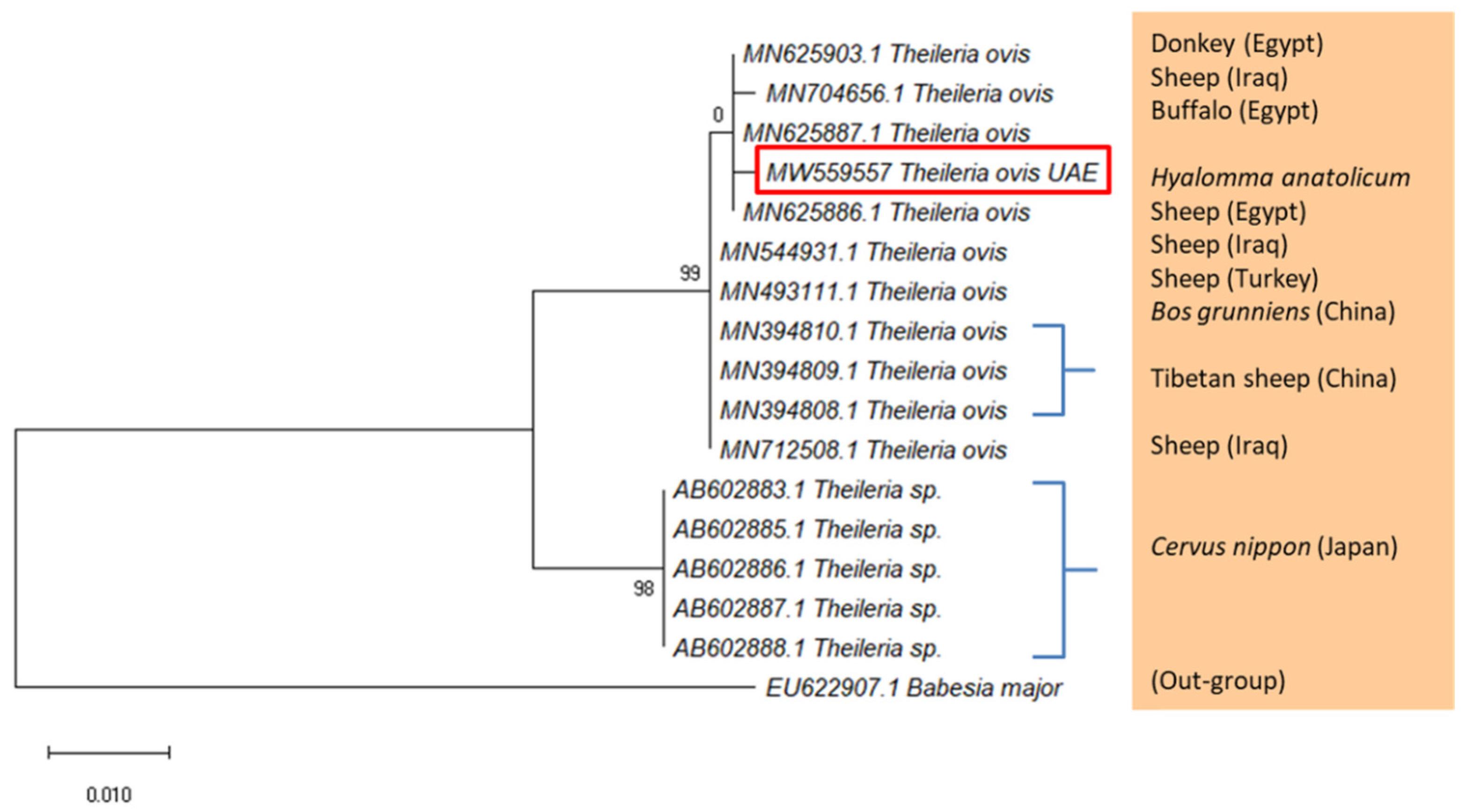
2.4. Detection of Piroplasmids
2.5. PCR-Based Infection Rates of Tick-Borne Microbes
3. Discussion
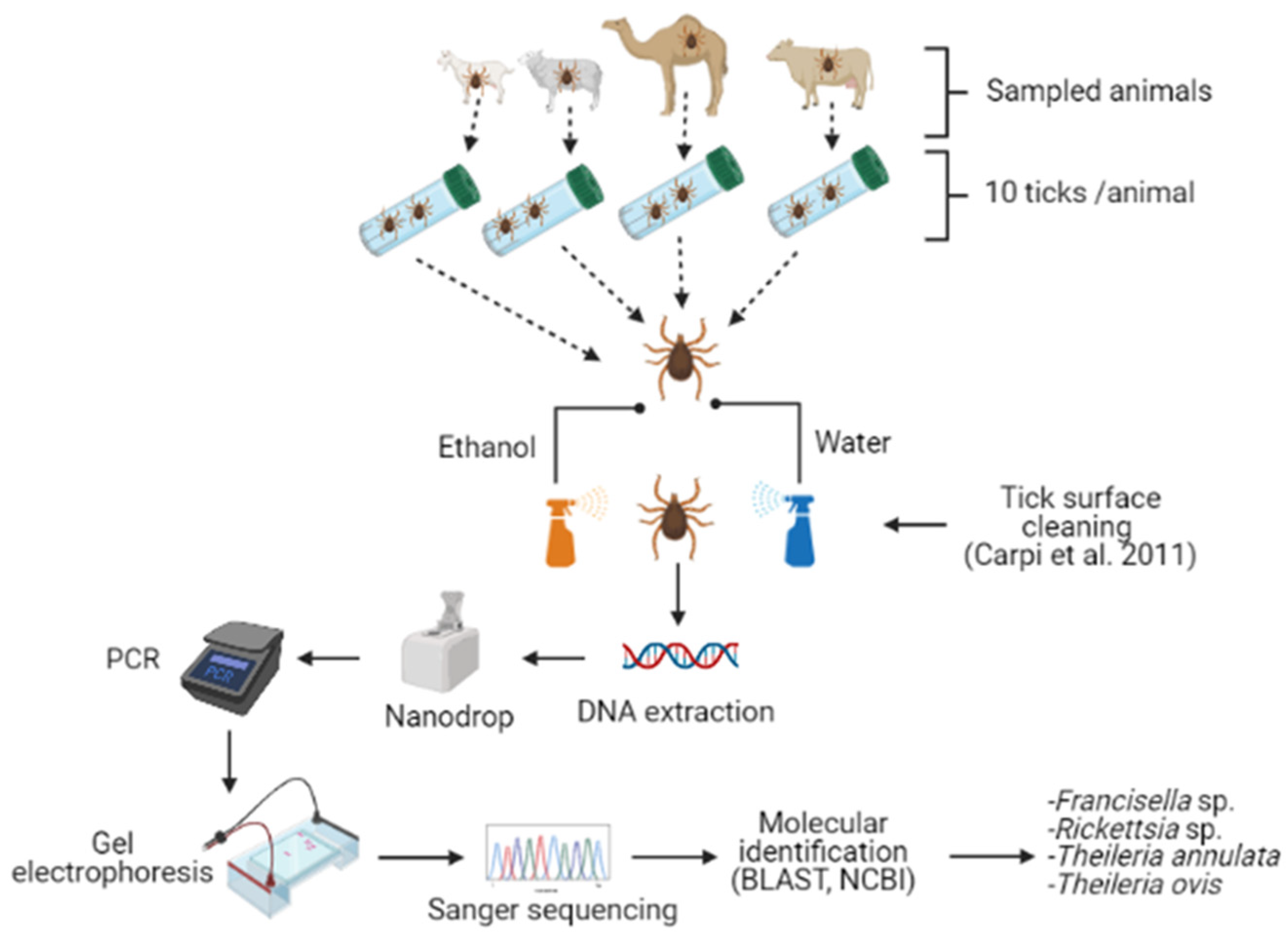
4. Materials and Methods
4.1. Ethics Statement
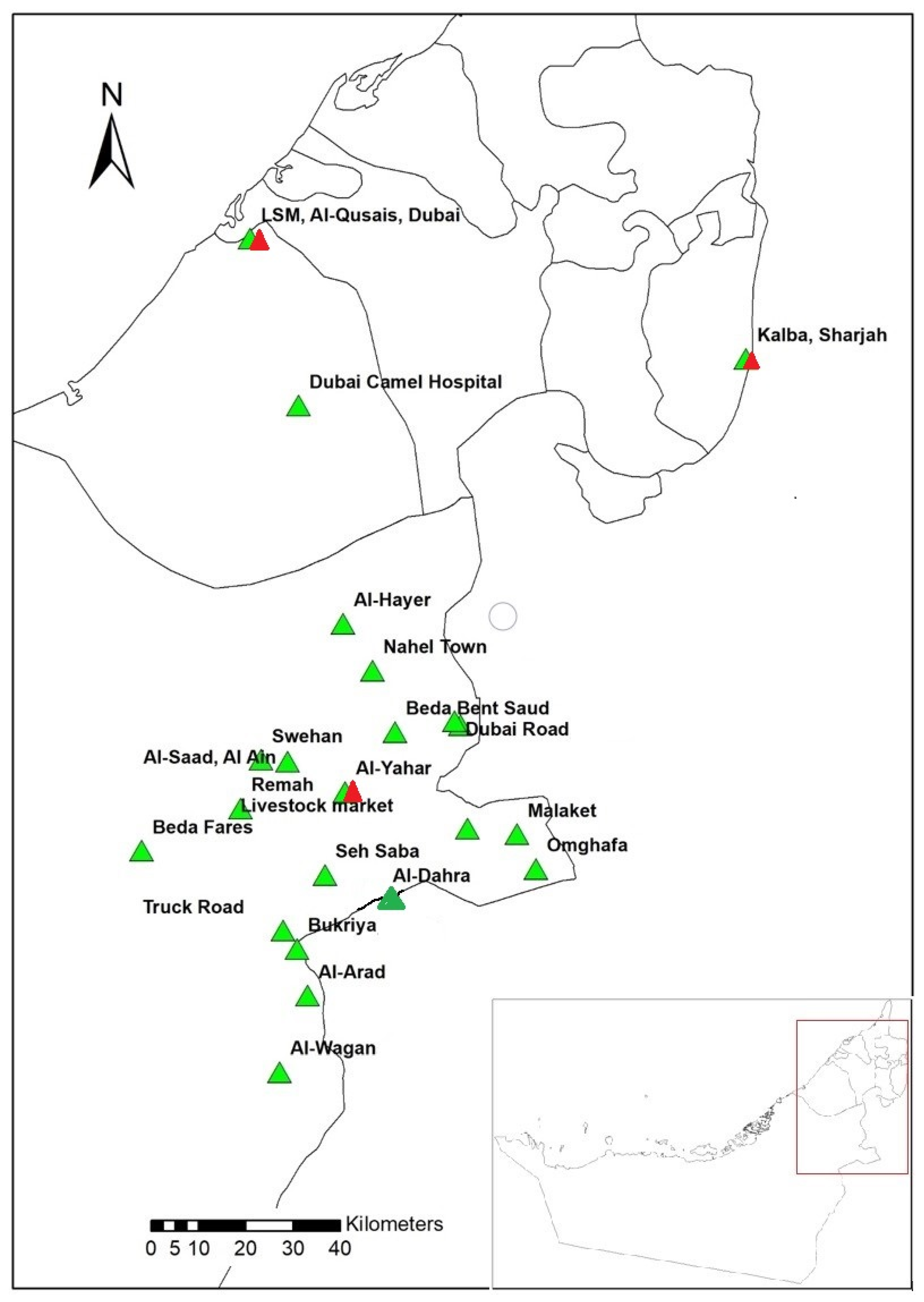
4.2. Study Area, Tick Collection and Identification
4.3. Genomic DNA Extraction
4.4. Polymerase Chain Reaction
4.5. Agarose Gel Electrophoresis and Amplicon Purification
4.6. DNA Sequencing, Phylogenetic Analysis, and Microorganism Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfäffle, M.; Littwin, N.; Muders, S.V.; Petney, T.N. The ecology of tick-borne diseases. Int. J. Parasitol. 2013, 43, 1059–1077. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.; Gadalla, A.A.H.; Postigo, M.; Al-Hamidhi, S.; Tageldin, M.H.; Skariah, S.; Sultan, A.A.; Johnson, E.H.; Shiels, B.; Pain, A.; et al. Dynamics and within-host interaction of Theileria lestoquardi and T. ovis among naive sheep in Oman. Sci. Rep. 2020, 10, 19802. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; de la Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Rybak, M.; Solano-Gallego, L.; Millán, J. A review of piroplasmid infections in wild carnivores worldwide: Importance for domestic animal health and wildlife conservation. Parasites Vectors 2016, 9, 538. [Google Scholar] [CrossRef] [Green Version]
- Perveen, N.; Muzaffar, S.B.; Al-deeb, M.A. Ticks and Tick-Borne Diseases of Livestock in the Middle East and North Africa: A Review. Insects 2021, 12, 83. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Horak, I.G. The genus Hyalomma koch, 1844: V. re-evaluation of the taxonomic rank of taxa comprising the H. (Euhyalomma) marginatum koch complex of species (acari: Ixodidae) with redescription of all parasitic stages and notes on biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Capek, M.; Literak, I.; Kocianova, E.; Sychra, O.; Najer, T.; Trnka, A.; Kverek, P. Ticks of the Hyalomma marginatum complex transported by migratory birds into Central Europe. Ticks Tick Borne Dis. 2014, 5, 489–493. [Google Scholar] [CrossRef]
- Dennis, D.T.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Friedlander, A.M.; Hauer, J.; Layton, M.; et al. Tularemia as a biological weapon: Medical and public health management. J. Am. Med. Assoc. 2001, 285, 2763–2773. [Google Scholar] [CrossRef]
- Ellis, J.; Oyston, P.C.F.; Green, M.; Titball, R.W. Tularemia. Clin. Microbiol. Rev. 2002, 15, 631–646. [Google Scholar] [CrossRef] [Green Version]
- Gürcan, Ş. Epidemiology of tularemia. Balk. Med. J. 2014, 31, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Shahsavari, S.; Bannazadeh Baghi, H.; Samadi Kafil, H.; Ebrahimzadeh Leylabadlo, H. Re-emerging tularemia in some Middle East Countries: What are the reasons? Iran. J. Public Health 2018, 47, 305–306. [Google Scholar]
- Kılıç, S. Francisella tularensis ve Türkiye’de Tularemi Epidemiyolojisine Genel Bir Bakış. Flora İnfeksiyon Hast. Klin. Mikrobiyoloji Derg. 2010, 15, 37–58. [Google Scholar]
- Petersen, J.M.; Mead, P.S.; Schriefer, M.E. Francisella tularensis: An arthropod-borne pathogen. Vet. Res. 2009, 40, 7. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, E.L.; Alford, H.I., Jr. Incidence of tularemia and Rocky Mountain spotted fever among common ticks of Arkansas. Am. J. Trop. Med. Hyg. 1955, 4, 310–317. [Google Scholar] [CrossRef]
- Keim, P.; Johansson, A.; Wagner, D.M. Molecular epidemiology, evolution, and ecology of Francisella. Ann. N. Y. Acad. Sci. 2007, 1105, 30–66. [Google Scholar] [CrossRef] [PubMed]
- Scoles, G.A. Phylogenetic analysis of the Francisella-like endosymbionts of Dermacentor ticks. J. Med. Entomol. 2004, 41, 277–286. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, I.L.; Toledo, A.; Carvalho, C.L.; Barandika, J.F.; Respicio-Kingry, L.B.; Garcia-Amil, C.; García-Pérez, A.L.; Olmeda, A.S.; Zé-Zé, L. Francisella species in ticks and animals, Iberian Peninsula. Ticks Tick Borne Dis. 2016, 7, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Azagi, T.; Klement, E.; Perlman, G.; Lustig, Y.; Mumcuoglu, K.Y. Francisella-Like Endosymbionts and Rickettsia Species in Local and Imported Hyalomma Ticks. Appl. Environ. Microbiol. 2017, 83, e01302-17. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.N.; Mitkova, N.; Reye, A.L.; Hübschen, J.M.; Vatcheva-Dobrevska, R.S.; Dobreva, E.G.; Kantardjiev, T.V.; Muller, C.P. Detection of new Francisella-like tick endosymbionts in Hyalomma spp. and Rhipicephalus spp. (Acari: Ixodidae) from Bulgaria. Appl. Environ. Microbiol. 2011, 77, 5562–5565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.V.; Scoles, G.A.; Fish, D.; O’Neill, S.L. Francisella-like endosymbionts of ticks. J. Invertebr. Pathol. 2000, 76, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Machado-Ferreira, E.; Piesman, J.; Zeidner, N.S.; Soares, C.A.G. Francisella-like endosymbiont DNA and Francisella tularensis virulence-related genes in Brazilian ticks (Acari: Ixodidae). J. Med. Entomol. 2009, 46, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, N.H.; Abdel-Moein, K.A.; Zaher, H.M. Molecular detection of Francisella spp. among ticks attached to camels in Egypt. Vector Borne Zoonotic Dis. 2017, 17, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Perveen, N.; Muzaffar, S.B.; Vijayan, R.; Al-Deeb, M.A. Microbial communities associated with the camel tick, Hyalomma dromedarii: 16S rRNA gene-based analysis. Sci. Rep. 2020, 10, 17305. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Raoult, D. Tick-borne rickettsioses around the world: Emerging diseases challenging old concepts. Clin. Microbiol. Rev. 2005, 18, 719–756. [Google Scholar] [CrossRef] [Green Version]
- Raoult, D.; Roux, V. Rickettsioses as paradigms of new or emerging infectious diseases. Clin. Microbiol. Rev. 1997, 10, 694–719. [Google Scholar] [CrossRef]
- Al-Deeb, M.A.; Muzaffar, S.B.; Abu-Zeid, Y.A.; Enan, M.R.; Karim, S. First record of a spotted fever group Rickettsia sp. and Theileria annulata in Hyalomma dromedarii (Acari: Ixodidae) ticks in the United Arab Emirates. Fla. Entomol. 2015, 98, 135–139. [Google Scholar] [CrossRef]
- Demoncheaux, J.P.; Socolovschi, C.; Davoust, B.; Haddad, S.; Raoult, D.; Parola, P. First detection of Rickettsia aeschlimannii in Hyalomma dromedarii ticks from Tunisia. Ticks Tick Borne Dis. 2012, 3, 398–402. [Google Scholar] [CrossRef]
- Loftis, A.D.; Reeves, W.K.; Szumlas, D.E.; Abbassy, M.M.; Helmy, I.M.; Moriarity, J.R.; Dasch, G.A. Rickettsial agents in Egyptian ticks collected from domestic animals. Exp. Appl. Acarol. 2006, 40, 67–81. [Google Scholar] [CrossRef]
- Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Genome sequence of the endosymbiont Rickettsia peacockii and comparison with virulent Rickettsia rickettsii: Identification of virulence factors. PLoS ONE 2009, 4, e8361. [Google Scholar] [CrossRef]
- Aktas, M.; Dumanli, N.; Angin, M. Cattle infestation by Hyalomma ticks and prevalence of Theileria in Hyalomma species in the east of Turkey. Vet. Parasitol. 2004, 119, 1–8. [Google Scholar] [CrossRef]
- Youssef, S.Y.; Yasien, S. Vector identification and clinical, hematological, biochemical, and parasitological characteristics of camel (Camelus dromedarius) theileriosis in Egypt. Trop. Anim. Health Prod. 2015, 47, 649–656. [Google Scholar] [CrossRef]
- Al-Fahdi, A.; Alqamashoui, B.; Al-Hamidhi, S.; Kose, O.; Tageldin, M.H.; Bobade, P.; Johnson, E.H.; Hussain, A.R.; Karagenc, T.; Tait, A.; et al. Molecular surveillance of Theileria parasites of livestock in Oman. Ticks Tick Borne Dis. 2017, 8, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Perveen, N.; Muzaffar, S.B.; Al-Deeb, M.A. Population dynamics of Hyalomma dromedarii on camels in the United Arab Emirates. Insects 2020, 11, 320. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Daszak, P.; Wood, J.L.N. One health, emerging infectious diseases and wildlife: Two decades of progress? Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binetruy, F.; Buysse, M.; Lejarre, Q.; Barosi, R.; Villa, M.; Rahola, N.; Paupy, C.; Ayala, D.; Duron, O. Microbial community structure reveals instability of nutritional symbiosis during the evolutionary radiation of Amblyomma ticks. Mol. Ecol. 2020, 29, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, M.; Rymaszewska, A. Expansion of Tick-Borne Rickettsioses in the World. Microorganisms 2020, 8, 1906. [Google Scholar] [CrossRef] [PubMed]
- Paddock, C.D.; Sumner, J.W.; Comer, J.A.; Zaki, S.R.; Goldsmith, C.S.; Goddard, J.; McLellan, S.L.F.; Tamminga, C.L.; Ohl, C.A. Rickettsia parkeri: A Newly Recognized Cause of Spotted Fever Rickettsiosis in the United States. Clin. Infect. Dis. 2004, 38, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Delgado-de la Mora, J.; Sánchez-Montes, S.; Licona-Enríquez, J.D.; de la Mora, D.D.; Paddock, C.D.; Beati, L.; Colunga-Salas, P.; Guzmán-Cornejo, C.; Zambrano, M.L.; Karpathy, S.E.; et al. Rickettsia parkeri and Candidatus Rickettsia andeanae in Ticks of the Amblyomma maculatum Group, Mexico. Emerg. Infect. Dis. 2019, 25, 836–838. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Moraru, G.M.; Stokes, J.V.; Wills, R.W.; Mitchell, E.; Unz, E.; Moore-Henderson, B.; Harper, A.B.; Varela-Stokes, A.S. Rickettsia parkeri and “Candidatus Rickettsia andeanae” in Questing Amblyomma maculatum (Acari: Ixodidae) From Mississippi. J. Med. Entomol. 2018, 54, 476–480. [Google Scholar] [CrossRef]
- Noden, B.H.; Roselli, M.A.; Loss, S.R. Rickettsia parkeri and Candidatus Rickettsia andeanae in Amblyomma maculatum Group Ticks. Emerg. Infect. Dis. 2020, 26, 2018–2021. [Google Scholar] [CrossRef] [Green Version]
- Paddock, C.D.; Denison, A.M.; Dryden, M.W.; Noden, B.H.; Lash, R.R.; Abdelghani, S.S.; Evans, A.E.; Kelly, A.R.; Hecht, J.A.; Karpathy, S.E.; et al. High prevalence of “Candidatus Rickettsia andeanae” and apparent exclusion of Rickettsia parkeri in adult Amblyomma maculatum (Acari: Ixodidae) from Kansas and Oklahoma. Ticks Tick Borne Dis. 2015, 6, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, O.; Binetruy, F.; Noël, V.; Cremaschi, J.; McCoy, K.D.; Arnathau, C.; Plantard, O.; Goolsby, J.; Pérez de León, A.A.; Heylen, D.J.A.; et al. Evolutionary changes in symbiont community structure in ticks. Mol. Ecol. 2017, 26, 2905–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef]
- Bilgic, H.B.; Aksulu, A.; Bakırcı, S.; Unlu, A.H.; Kose, O.; Hacılarlıoglu, S.; Weir, W.; Karagenc, T. Infection dynamics of Theileria annulata over a disease season following cell line vaccination. Vet. Parasitol. 2019, 265, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanazi, A.D.; Alouffi, A.S.; Alshahrani, M.Y.; Alyousif, M.S.; Abdullah, H.H.A.M.; Allam, A.M.; Elsawy, B.S.M.; Abdel-Shafy, S.; Alsulami, M.N.; Khan, A.; et al. A report on tick burden and molecular detection of tick-borne pathogens in cattle blood samples collected from four regions in Saudi Arabia. Ticks Tick Borne Dis. 2021, 12, 101652. [Google Scholar] [CrossRef]
- Walker, A.R.; Bouattour, A.; Camicas, J.-L.; Estrada-Peña, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; ISBN 095451730X. [Google Scholar]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The Genus Hyalomma: VII. Redescription of all Parasitic Stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef]
- Carpi, G.; Cagnacci, F.; Wittekindt, N.E.; Zhao, F.; Qi, J.; Lynn, P.; Drautz, D.I.; Rizzoli, A.; Schuster, S.C. Metagenomic Profile of the Bacterial Communities Associated with Ixodes ricinus Ticks. PLoS ONE 2011, 6, e25604. [Google Scholar] [CrossRef]
- Duzlu, O.; Yildirim, A.; Inci, A.; Gumussoy, K.S.; Ciloglu, A.; Onder, Z. Molecular Investigation of Francisella-Like Endosymbiont in Ticks and Francisella tularensis in Ixodid Ticks and Mosquitoes in Turkey. Vector Borne Zoonotic Dis. 2016, 16, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Blair, P.J.; Jiang, J.; Schoeler, G.B.; Anaya, E.; Cespedes, M.; Felices, V.; Guevara, C.; Mendoza, L.; Villaseca, P.; Sumner, J.W.; et al. Characterization of Spotted Fever Group Rickettsiae in Flea and Tick Specimens from Northern Peru. J. Clin. Microbiol. 2004, 42, 4961–4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, R.; Vojta, L.; Mrljak, V.; Marinculić, A.; Beck, A.; Živičnjak, T.; Cacciò, S.M. Diversity of Babesia and Theileria species in symptomatic and asymptomatic dogs in Croatia. Int. J. Parasitol. 2009, 39, 843–848. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Tick Species | Number of Animals | Number of Samples | Number of Positive Samples (Total Infection Rate) | |||
|---|---|---|---|---|---|---|---|
| Francisella sp. | Rickettsia sp. | T. annulata | T. ovis | ||||
| Camel | H. dromedarii | 518 | 516 | 30 (5.81) | 7 (1.36) | 0 (0) | 0 (0) |
| Sheep * | H. anatolicum | 70 | 14 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Goat * | H. anatolicum | 34 | 10 | 0 (0) | 0 (0) | 0 (0) | 1 (10) |
| Cow * | H. anatolicum | 26 | 22 | 0 (0) | 0 (0) | 1 (4.55) | 0 (0) |
| Total | 648 | 562 | 30 | 7 | 1 | 1 | |
| Pathogen | Target Gene | Primer | Sequence (5′–3′) | Cycle Conditions | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Francisella sp. | 16S rRNA | Fr153F0.1 Fr1281R0.1 | GCCCATTTGAGGGGGATACC GGACTAAGAGTACCTTTTTGAGT | 95 °C 4 min 40 cycles: 94 °C 30 s 60 °C 45 s 72 °C 60 s 72 °C 20 min | 1151 | [51] |
| Rickettsia sp. | ompA | RR 190-70 (1st PCR) RR 190-701(1st PCR) | ATGGCGAATATTTCTCCAAAA GTTCCGTTAATGGCAGCATCT | 94 °C 1 min 35 cycles: 94 °C 30 s 50 °C 1 min 68 °C 4 min 72 °C 20 min | 590 | [52] |
| 190-FN1 (nested) 190-RN1 (nested) | AAGCAATACAACAAGGTC TGACAGTTATTATACCTC | 540 | ||||
| Theileria sp. | ssrRNA | Pirop-F Pirop-F | GTCTTGTAATTGGAATGATGG CCAAAGACTTTGATTTCTCTC | 94 °C 2 min 35 cycles: 94 °C 30 s 50 °C 30 s 72 °C 60 s 72 °C 7 min | 560 | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perveen, N.; Muzaffar, S.B.; Al-Deeb, M.A. Four Tick-Borne Microorganisms and Their Prevalence in Hyalomma Ticks Collected from Livestock in United Arab Emirates. Pathogens 2021, 10, 1005. https://doi.org/10.3390/pathogens10081005
Perveen N, Muzaffar SB, Al-Deeb MA. Four Tick-Borne Microorganisms and Their Prevalence in Hyalomma Ticks Collected from Livestock in United Arab Emirates. Pathogens. 2021; 10(8):1005. https://doi.org/10.3390/pathogens10081005
Chicago/Turabian StylePerveen, Nighat, Sabir Bin Muzaffar, and Mohammad Ali Al-Deeb. 2021. "Four Tick-Borne Microorganisms and Their Prevalence in Hyalomma Ticks Collected from Livestock in United Arab Emirates" Pathogens 10, no. 8: 1005. https://doi.org/10.3390/pathogens10081005
APA StylePerveen, N., Muzaffar, S. B., & Al-Deeb, M. A. (2021). Four Tick-Borne Microorganisms and Their Prevalence in Hyalomma Ticks Collected from Livestock in United Arab Emirates. Pathogens, 10(8), 1005. https://doi.org/10.3390/pathogens10081005