
Genotyping of Porcine Circovirus 2 (PCV-2) in Vaccinated Pigs Suffering from PCV-2-Systemic Disease between 2009 and 2020 in Spain

 , , and
, , and
Abstract
:1. Introduction
2. Results
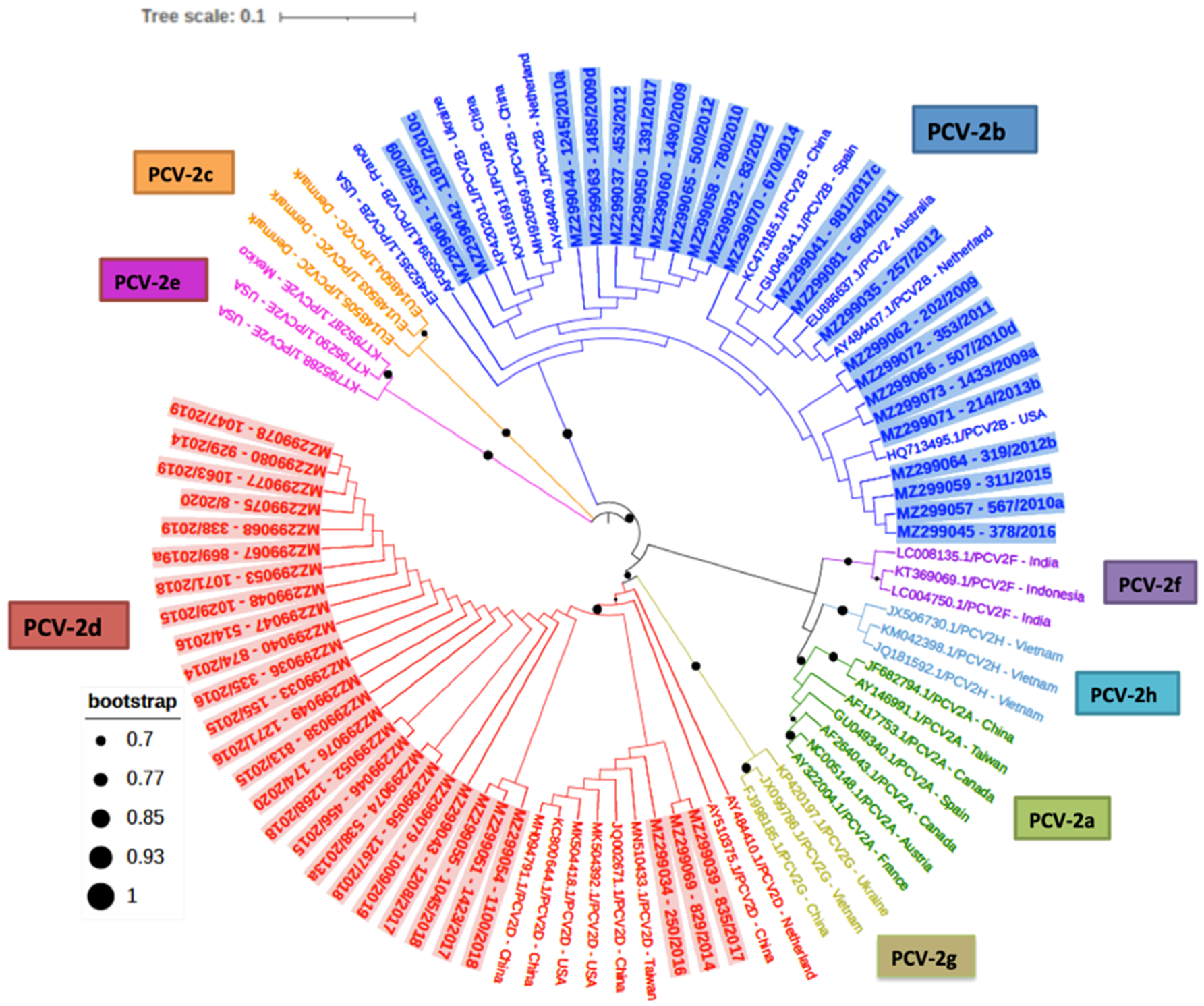
2.1. PCV-2 Phylogenetic Analyses and Genotyping
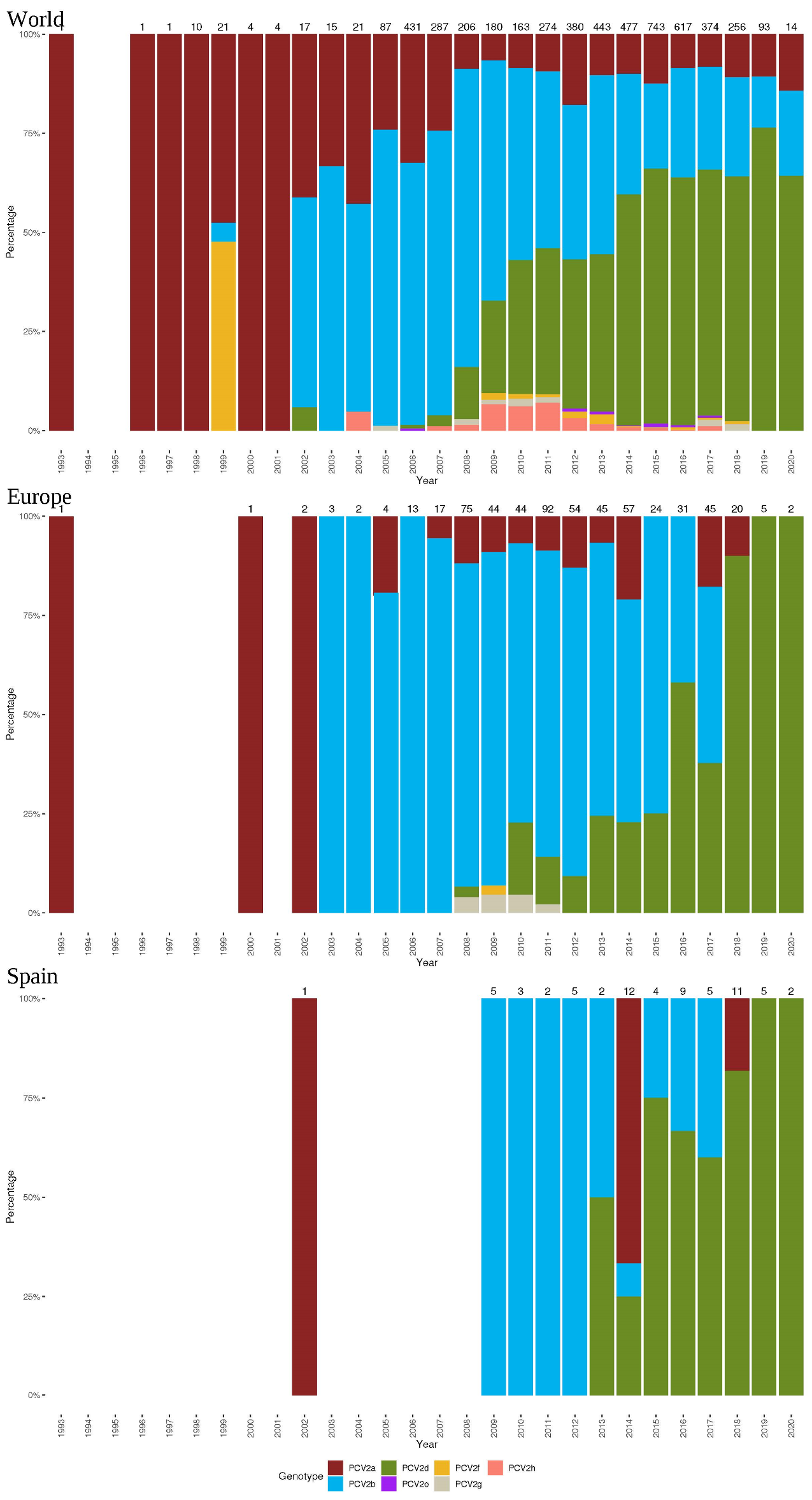
2.2. PCV-2 Genotype Frequencies at the World, European, and Spanish Levels
3. Discussion
4. Materials and Methods
4.1. Sample Selection
4.2. Deparaffination and DNA Extraction Protocol
4.3. PCV-2 CAP Gene Amplification and Sequencing
4.4. PCV-2 ORF2 Phylogenetic Analyses
4.5. Phylogenetic Analyses of PCV-2 Sequences at World, European, and Spanish Levels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Segales, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Alarcon, P.; Rushton, J.; Wieland, B. Cost of post-weaning multi-systemic wasting syndrome and porcine circovirus type-2 subclinical infection in England—an economic disease model. Prev. Vet. Med. 2013, 110, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segales, J.; Allan, G.M.; Domingo, M. Porcine circovirus diseases. Anim. Health Res. Rev. 2005, 6, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Segales, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar]
- Wang, S.; Xin, C.; Wu, X.; Shi, J.; Peng, Z.; Sun, P.; Wang, Y.; Xu, S.; Yang, Y.; Zhang, F. Genetic characterization of Porcine circovirus type 2 from 2013 to 2018 in Shandong Province, China. Vet. Med. Sci. 2020, 6, 76–81. [Google Scholar] [CrossRef]
- Wang, Y.; Noll, L.; Lu, N.; Porter, E.; Stoy, C.; Zheng, W.; Liu, X.; Peddireddi, L.; Niederwerder, M.; Bai, J. Genetic diversity and prevalence of porcine circovirus type 3 (PCV3) and type 2 (PCV2) in the Midwest of the USA during 2016–2018. Transbound. Emerg. Dis. 2020, 67, 1284–1294. [Google Scholar] [CrossRef]
- Dupont, K.; Nielsen, E.O.; Baekbo, P.; Larsen, L.E. Genomic analysis of PCV2 isolates from Danish archives and a current PMWS case-control study supports a shift in genotypes with time. Vet. Microbiol. 2008, 128, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, S.; Cai, H.Y.; Delay, J.; Youssef, S.A.; McEwen, B.J.; Gagnon, C.A.; Tremblay, D.; Hazlett, M.; Lusis, J.; Fairles, J.; et al. The emergence of a new strain of porcine circovirus-2 in Ontario and Quebec swine and its association with severe porcine circovirus associated disease 2004–2006. Can. J. Vet. Res. 2008, 72, 259–268. [Google Scholar]
- Timmusk, S.; Wallgren, P.; Brunborg, I.M.; Wikstrom, F.H.; Allan, G.; Meehan, B.; McMenamy, M.; McNeilly, F.; Fuxler, L.; Belak, K.; et al. Phylogenetic analysis of porcine circovirus type 2 (PCV2) pre- and post-epizootic postweaning multisystemic wasting syndrome (PMWS). Virus Genes 2008, 36, 509–520. [Google Scholar] [CrossRef]
- Cortey, M.; Pileri, E.; Sibila, M.; Pujols, J.; Balasch, M.; Plana, J.; Segales, J. Genotypic shift of porcine circovirus type 2 from PCV-2a to PCV-2b in Spain from 1985 to 2008. Vet. J. 2011, 187, 363–368. [Google Scholar] [CrossRef]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef]
- Franzo, G.; Cortey, M.; Segales, J.; Hughes, J.; Drigo, M. Phylodynamic analysis of porcine circovirus type 2 reveals global waves of emerging genotypes and the circulation of recombinants forms. Mol. Phylogenet. Evol. 2016, 100, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, G.T.; Lin, Y.C.; Lin, W.H.; Lin, J.H.; Chiou, M.T.; Liu, H.F.; Lin, C.N. Phylogeographic and genetic characterization of porcine circovirus type 2 in Taiwan from 2001–2017. Sci. Rep. 2019, 9, 10782. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Langohr, I. Current state of knowledge on porcine circovirus type 2-associated lesions. Vet. Pathol. 2013, 50, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Ssemadaali, M.A.; Ilha, M.; Ramamoorthy, S. Genetic diversity of porcine circovirus type 2 and implications for detection and control. Res. Vet. Sci. 2015, 103, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Oh, T.; Yang, S.; Cho, H.; Kang, I.; Chae, C. Evaluation of a porcine circovirus type 2a (PCV2a) vaccine efficacy against experimental PCV2a, PCV2b, and PCV2d challenge. Vet. Microbiol. 2019, 231, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kang, I.; Oh, T.; Yang, S.; Park, K.H.; Min, K.D.; Ham, H.J.; Chae, C. Comparative study of the virulence of 3 major Korean porcine circovirus type 2 genotypes (a, b, and d). Can. J. Vet. Res. 2020, 84, 235–240. [Google Scholar]
- Opriessnig, T.; Karuppannan, A.K.; Halbur, P.G.; Calvert, J.G.; Nitzel, G.P.; Matzinger, S.R.; Meng, X.J. Porcine circovirus type 2a or 2b based experimental vaccines provide protection against PCV2d/porcine parvovirus 2 co-challenge. Vaccine 2020, 38, 1975–1981. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Wang, H.; Feng, Y.; Song, M.; Li, Q.; Li, J. Genetic variation and phylogenetic analysis of Porcine circovirus type 2 in China from 2016 to 2018. Acta. Virol. 2019, 63, 459–468. [Google Scholar] [CrossRef]
- Weissenbacher-Lang, C.; Kristen, T.; Mendel, V.; Brunthaler, R.; Schawarz, L.; Weissenbock, H. Porcine circovirus type 2 (PCV2) genotyping in Austrian pigs in the years 2002 to 2017. BMC Vet. Res. 2020, 16, 198. [Google Scholar] [CrossRef]
- Mone, N.K.; Clark, N.J.; Kyaw-Tanner, M.; Turni, C.; Barnes, T.S.; Parke, C.R.; Alawneh, J.A.; Blackwall, P.J.; Meers, J. Genetic analysis of porcine circovirus type 2 (PCV2) in Queensland, Australia. Aust. Vet. J. 2020, 98, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Segales, J. Best practice and future challenges for vaccination against porcine circovirus type 2. Expert Rev. Vaccines 2015, 14, 473–487. [Google Scholar] [CrossRef]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G.; Matzinger, S.R.; Meng, X.J. Mutant USA strain of porcine circovirus type 2 (mPCV2) exhibits similar virulence to the classical PCV2a and PCV2b strains in caesarean-derived, colostrum-deprived pigs. J. Gen. Virol. 2014, 95, 2495–2503. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Xie, J.; Theuns, S.; Nauwynck, H.J. Changes on the viral capsid surface during the evolution of porcine circovirus type 2 (PCV2) from 2009 till 2018 may lead to a better receptor binding. Virus Evol. 2019, 5, vez026. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Tucciarone, C.M.; Cecchinato, M.; Drigo, M. Porcine circovirus type 2 (PCV2) evolution before and after the vaccination introduction: A large scale epidemiological study. Sci. Rep. 2016, 6, 39458. [Google Scholar] [CrossRef]
- Franzo, G.; Segalés, J. Porcine circovirus 2 genotypes, immunity and vaccines: Multiple genotypes but one single serotype. Pathogens 2020, 9, 1049. [Google Scholar] [CrossRef] [PubMed]
- Grierson, S.S.; Werling, D.; Bidewell, C.; Williamson, S. Characterisation of porcine circovirus type 2 in porcine circovirus disease cases in England and Wales. Vet. Rec. 2018, 182, 22. [Google Scholar] [CrossRef] [PubMed]
- Grau-Roma, L.; Baekbo, P.; Rose, N.; Wallgren, P.; Fraile, L.; Larsen, L.E.; Segalés, J. Clinical and laboratory studies on herds affected with postweaning multisystemic wasting syndrome in Denmark, Frane, Spain and Sweden: Disease progression and a proposal for herd case definition. J. Swine Health Prod. 2012, 20, 129–136. [Google Scholar]
- Rosell, C.; Segales, J.; Plana-Duran, J.; Balasch, M.; Rodriguez-Arrioja, G.M.; Kennedy, S.; Allan, G.M.; McNeilly, F.; Latimer, K.S.; Domingo, M. Pathological, immunohistochemical, and in-situ hybridization studies of natural cases of postweaning multisystemic wasting syndrome (PMWS) in pigs. J. Comp. Pathol. 1999, 120, 59–78. [Google Scholar] [CrossRef]
- Oliver-Ferrando, S.; Segales, J.; Lopez-Soria, S.; Callen, A.; Merdy, O.; Joisel, F.; Sibila, M. Evaluation of natural porcine circovirus type 2 (PCV2) subclinical infection and seroconversion dynamics in piglets vaccinated at different ages. Vet. Res. 2016, 47, 121. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stehcer, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Year | N | Genotypes | |
|---|---|---|---|
| PCV-2b (%) | PCV-2d (%) | ||
| 2009 | 5 | 5 (100) | 0 (0) |
| 2010 | 5 | 5 (100) | 0 (0) |
| 2011 | 2 | 2 (100) | 0 (0) |
| 2012 | 5 | 5 (100) | 0 (0) |
| 2013 | 2 | 1 (50) | 1 (50) |
| 2014 | 4 | 1 (25) | 3 (75) |
| 2015 | 5 | 1 (20) | 4 (80) |
| 2016 | 5 | 1 (20) | 4 (80) |
| 2017 | 5 | 2 (40) | 3 (60) |
| 2018 | 5 | 0 (0) | 5 (100) |
| 2019 | 5 | 0 (0) | 5 (100) |
| 2020 | 2 | 0 (0) | 2 (100) |
| Total | 50 | 23 (46) | 27 (54) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sibila, M.; Rocco, C.; Franzo, G.; Huerta, E.; Domingo, M.; Núñez, J.I.; Segalés, J. Genotyping of Porcine Circovirus 2 (PCV-2) in Vaccinated Pigs Suffering from PCV-2-Systemic Disease between 2009 and 2020 in Spain. Pathogens 2021, 10, 1016. https://doi.org/10.3390/pathogens10081016
Sibila M, Rocco C, Franzo G, Huerta E, Domingo M, Núñez JI, Segalés J. Genotyping of Porcine Circovirus 2 (PCV-2) in Vaccinated Pigs Suffering from PCV-2-Systemic Disease between 2009 and 2020 in Spain. Pathogens. 2021; 10(8):1016. https://doi.org/10.3390/pathogens10081016
Chicago/Turabian StyleSibila, Marina, Caterina Rocco, Giovanni Franzo, Eva Huerta, Mariano Domingo, José Ignacio Núñez, and Joaquim Segalés. 2021. "Genotyping of Porcine Circovirus 2 (PCV-2) in Vaccinated Pigs Suffering from PCV-2-Systemic Disease between 2009 and 2020 in Spain" Pathogens 10, no. 8: 1016. https://doi.org/10.3390/pathogens10081016
APA StyleSibila, M., Rocco, C., Franzo, G., Huerta, E., Domingo, M., Núñez, J. I., & Segalés, J. (2021). Genotyping of Porcine Circovirus 2 (PCV-2) in Vaccinated Pigs Suffering from PCV-2-Systemic Disease between 2009 and 2020 in Spain. Pathogens, 10(8), 1016. https://doi.org/10.3390/pathogens10081016