
Establishment of a Rhesus Macaque Model for Scrub Typhus Transmission: Pilot Study to Evaluate the Minimal Orientia tsutsugamushi Transmission Time by Leptotrombidium chiangraiensis Chiggers


 ,
, 
Abstract
:1. Introduction
2. Results
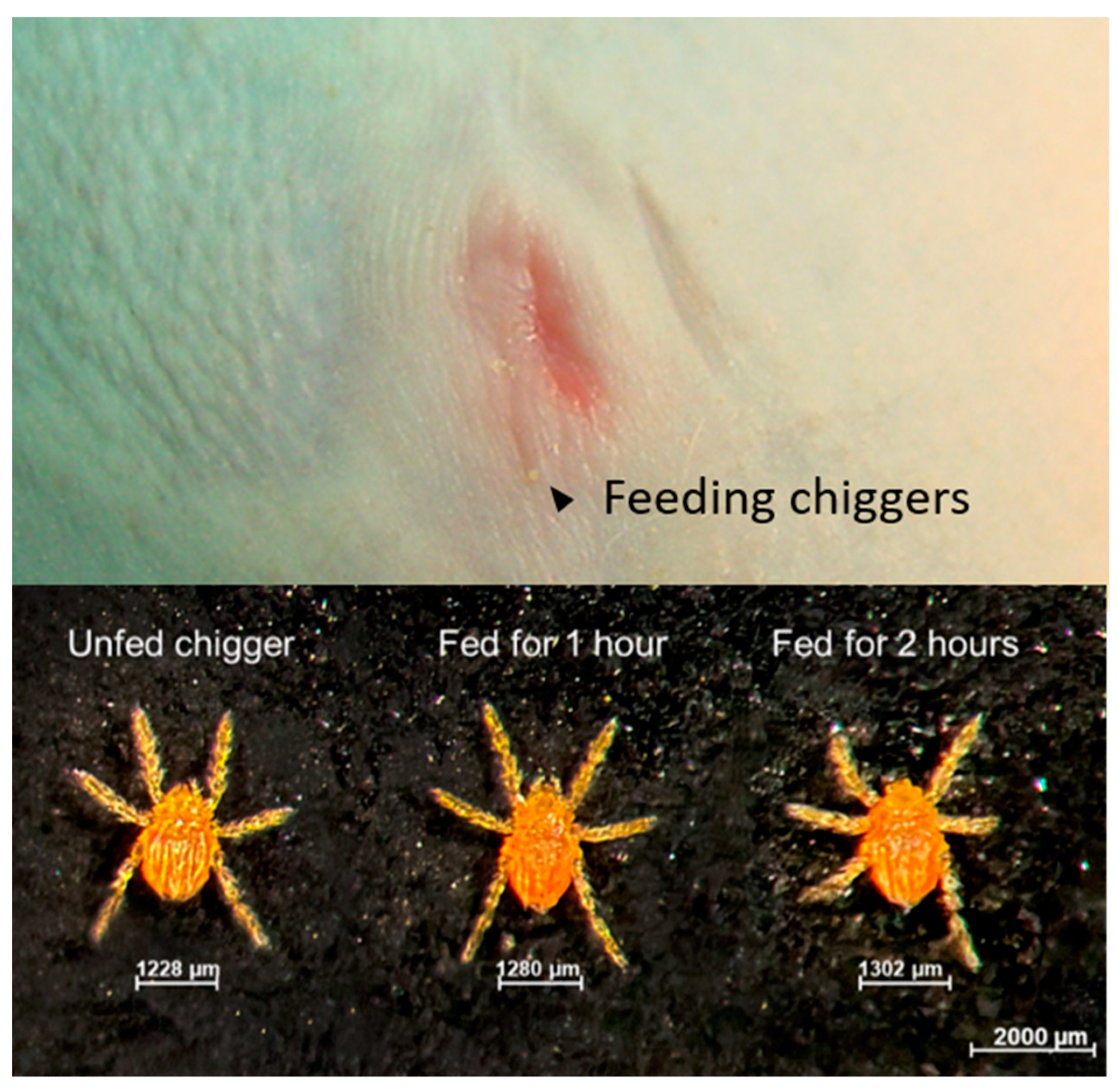
2.1. Bacterial Transmission to Rhesus Macaques Occurs Rapidly within 1 h of Chigger Feeding
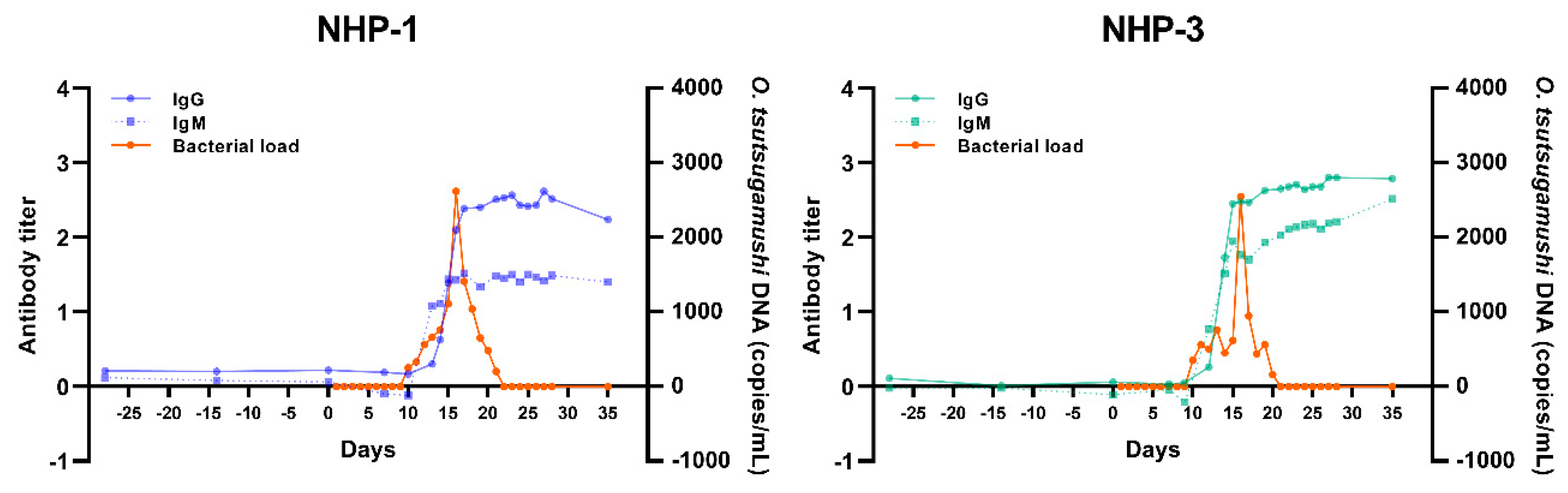
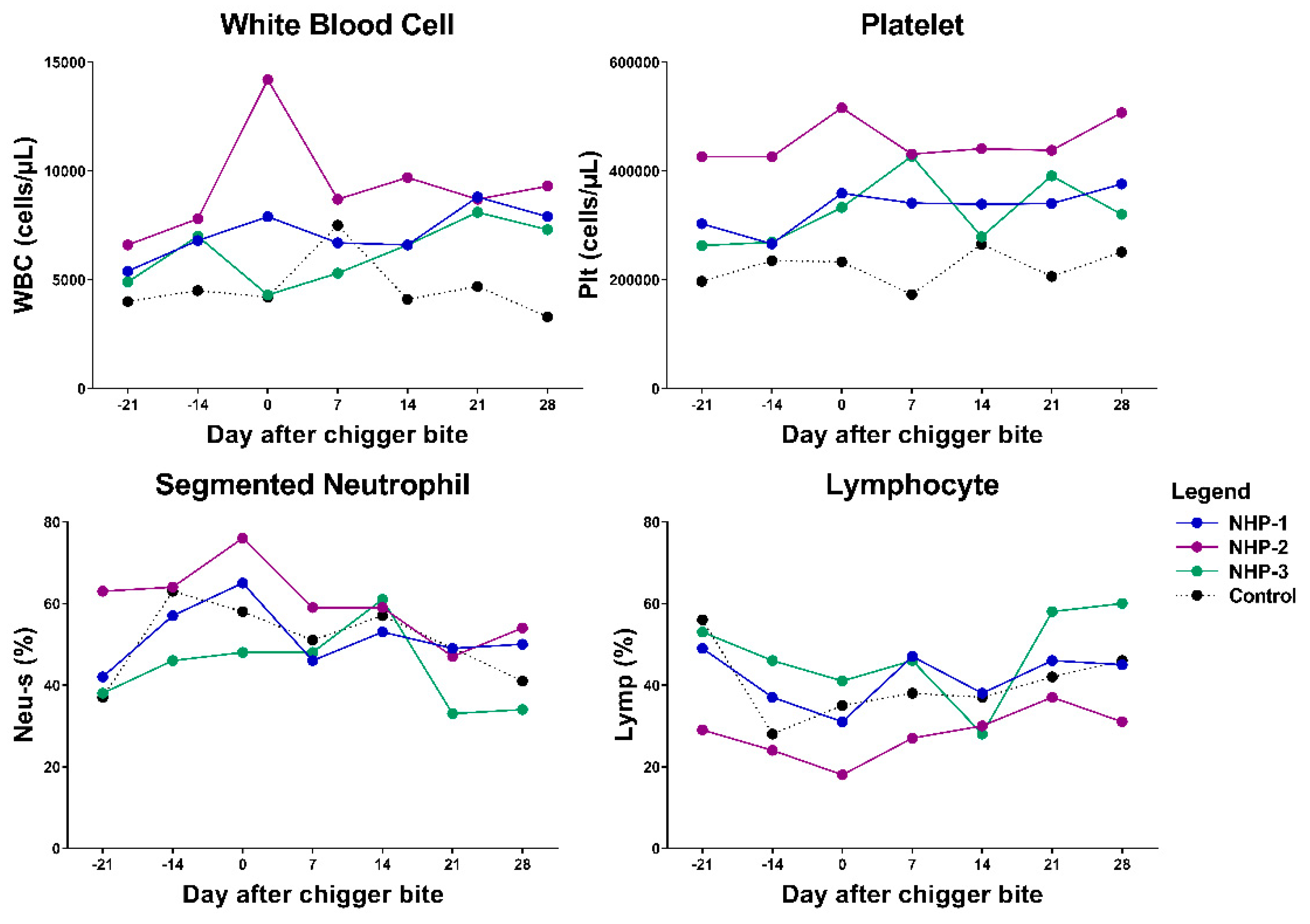
2.2. Bacteremia in Infected Rhesus Macaques Showed Comparable Trend to Those Observed in Scrub Typhus Patients
2.3. Specific Cellular Immune Responses to O. tsutsugamushi Infection
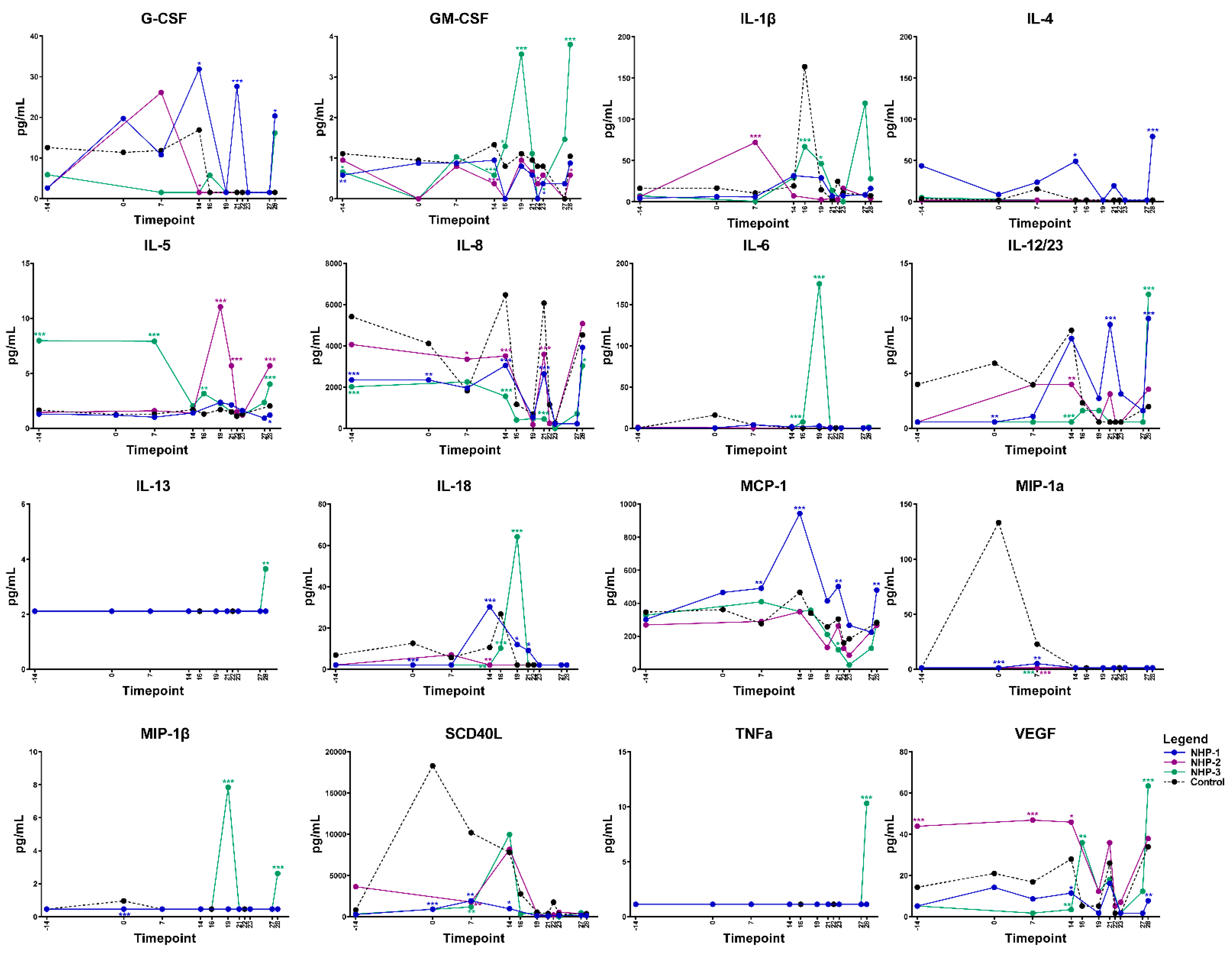
2.4. Elevated Levels of Serum Cytokines and Chemokines Are Associated with Progressive O. tsutsugamushi Infection
3. Discussion
4. Materials and Methods
4.1. Nonhuman Primates
4.2. Leptotrombidium sp. Chigger Mite Colonies
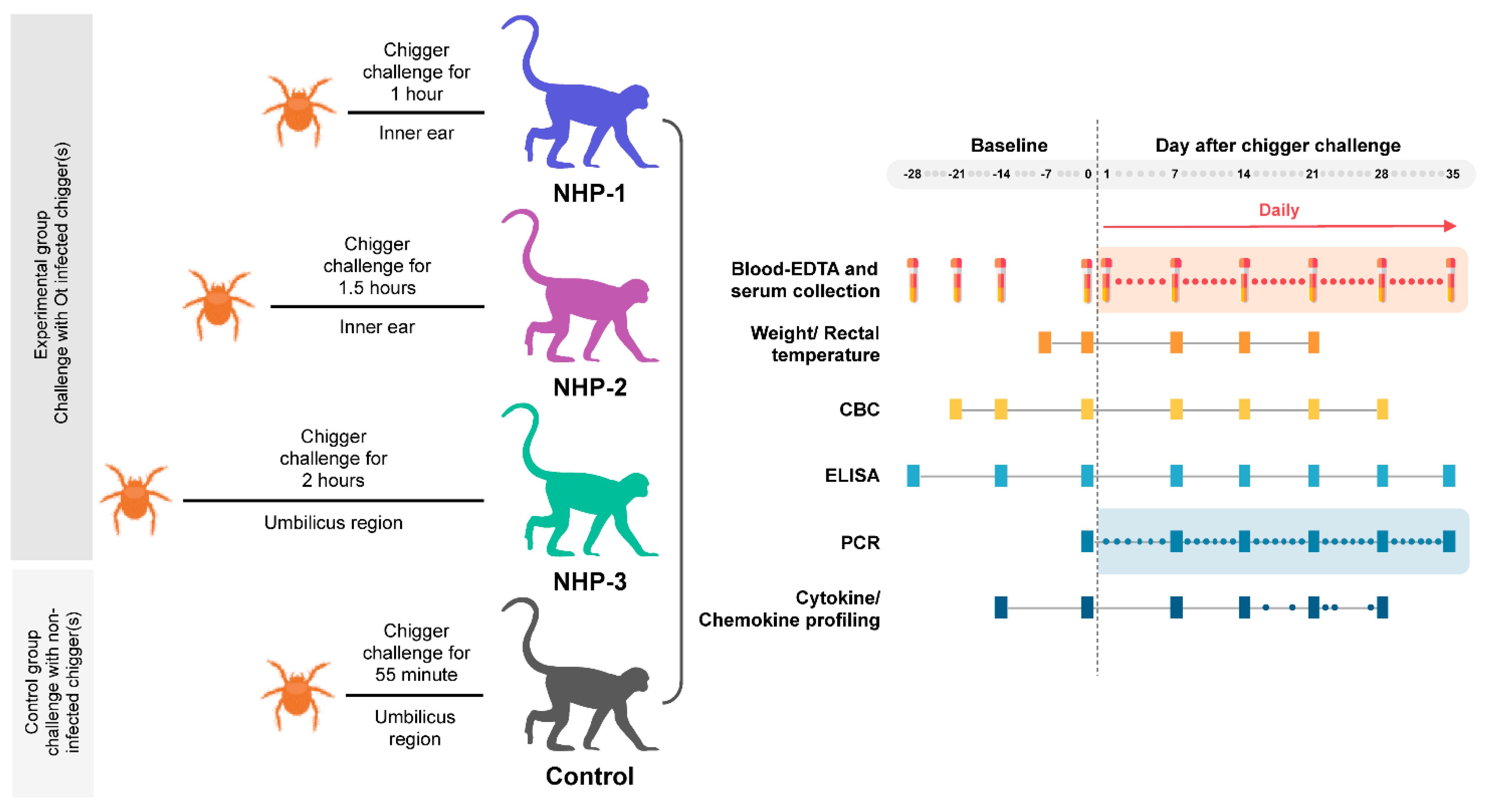
4.3. Chigger Challenge of Rhesus Macaques
4.3.1. Chigger Placement Experiment
4.3.2. Clinical Observation and Blood Sample Collection
4.4. Quantitation of O. tsutsugamushi Concentrations in Blood and Chigger Samples
4.5. Assessment of Host Immune Responses to O. tsutsugamushi Infection
4.5.1. Hematological Analysis
4.5.2. Serological Analysis
4.5.3. Multiplex Serum Cytokine and Chemokine Assay
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Disclaimer
Conflicts of Interest
References
- Abarca, K.; Martinez-Valdebenito, C.; Angulo, J.; Jiang, J.; Farris, C.M.; Richards, A.L.; Acosta-Jamett, G.; Weitzel, T. Molecular Description of a Novel Orientia Species Causing Scrub Typhus in Chile. Emerg. Infect. Dis. 2020, 26, 2148–2156. [Google Scholar] [CrossRef]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.; et al. Isolation of a novel Orientia species (O. chuto sp. nov.) from a patient infected in Dubai. J. Clin. Microbiol. 2010, 48, 4404–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masakhwe, C.; Linsuwanon, P.; Kimita, G.; Mutai, B.; Leepitakrat, S.; Yalwala, S.; Abuom, D.; Auysawasi, N.; Gilbreath, T.; Wanja, E.; et al. Identification and Characterization of Orientia chuto in Trombiculid Chigger Mites Collected from Wild Rodents in Kenya. J. Clin. Microbiol. 2018, 56, e01124-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitzel, T.; Martinez-Valdebenito, C.; Acosta-Jamett, G.; Jiang, J.; Richards, A.L.; Abarca, K. Scrub Typhus in Continental Chile, 2016–2018. Emerg. Infect. Dis. 2019, 25, 1214–1217. [Google Scholar] [CrossRef]
- Park, S.W.; Ha, N.Y.; Ryu, B.; Bang, J.H.; Song, H.; Kim, Y.; Kim, G.; Oh, M.D.; Cho, N.H.; Lee, J.K. Urbanization of scrub typhus disease in South Korea. PLoS Negl. Trop. Dis. 2015, 9, e0003814. [Google Scholar] [CrossRef] [PubMed]
- Linsuwanon, P.; Krairojananan, P.; Rodkvamtook, W.; Leepitakrat, S.; Davidson, S.; Wanja, E. Surveillance for Scrub Typhus, Rickettsial Diseases, and Leptospirosis in US and Multinational Military Training Exercise Cobra Gold Sites in Thailand. US Army Med. Dep. J. 2018, 1, 29–39. [Google Scholar]
- Li, Z.; Xin, H.; Sun, J.; Lai, S.; Zeng, L.; Zheng, C.; Ray, S.E.; Weaver, N.D.; Wang, L.; Yu, J.; et al. Epidemiologic Changes of Scrub Typhus in China, 1952–2016. Emerg. Infect. Dis. 2020, 26, 1091–1101. [Google Scholar] [CrossRef]
- Elliott, I.; Pearson, I.; Dahal, P.; Thomas, N.V.; Roberts, T.; Newton, P.N. Scrub typhus ecology: A systematic review of Orientia in vectors and hosts. Parasites Vectors 2019, 12, 513. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jamett, G.; Martinez-Valdebenito, C.; Beltrami, E.; Silva-de La Fuente, M.C.; Jiang, J.; Richards, A.L.; Weitzel, T.; Abarca, K. Identification of trombiculid mites (Acari: Trombiculidae) on rodents from Chiloe Island and molecular evidence of infection with Orientia species. PLoS Negl. Trop. Dis. 2020, 14, e0007619. [Google Scholar] [CrossRef]
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A review of the global epidemiology of scrub typhus. PLoS Negl. Trop. Dis. 2017, 11, e0006062. [Google Scholar] [CrossRef] [Green Version]
- Tsay, R.W.; Chang, F.Y. Serious complications in scrub typhus. J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 1998, 31, 240–244. [Google Scholar]
- Paris, D.H.; Shelite, T.R.; Day, N.P.; Walker, D.H. Unresolved problems related to scrub typhus: A seriously neglected life-threatening disease. Am. J. Trop. Med. Hyg. 2013, 89, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.M.; Won, K.J.; Park, C.Y.; Yu, K.D.; Kim, H.S.; Yang, T.Y.; Lee, J.H.; Kim, H.K.; Song, H.J.; Lee, S.H.; et al. Distribution of eschars on the body of scrub typhus patients: A prospective study. Am. J. Trop. Med. Hyg. 2007, 76, 806–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murhekar, M.V.; Mittal, M.; Prakash, J.A.; Pillai, V.M.; Mittal, M.; Girish Kumar, C.P.; Shinde, S.; Ranjan, P.; Oak, C.; Gupta, N.; et al. Acute encephalitis syndrome in Gorakhpur, Uttar Pradesh, India—Role of scrub typhus. J. Infect. 2016, 73, 623–626. [Google Scholar] [CrossRef]
- Yum, K.S.; Na, S.J.; Lee, K.O.; Ko, J.H. Scrub typhus meningo-encephalitis with focal neurologic signs and associated brain MRI abnormal findings: Literature review. Clin. Neurol. Neurosurg. 2011, 113, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Han, D.K.; Baek, H.J.; Shin, M.G.; Kim, J.W.; Kook, H.; Hwang, T.J. Scrub typhus-associated severe hemophagocytic lymphohistiocytosis with encephalomyelitis leading to permanent sequelae: A case report and review of the literature. J. Pediatric Hematol./Oncol. 2012, 34, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Hwang, J.H.; Lee, C.S. Dissemination of Orientia tsutsugamushi in a Case of Scrub Typhus. Am. J. Trop. Med. Hyg. 2019, 100, 235–236. [Google Scholar] [CrossRef]
- El Sayed, I.; Liu, Q.; Wee, I.; Hine, P. Antibiotics for treating scrub typhus. Cochrane Database Syst. Rev. 2018, 9, CD002150. [Google Scholar] [CrossRef]
- Taylor, A.J.; Paris, D.H.; Newton, P.N. A Systematic Review of Mortality from Untreated Scrub Typhus (Orientia tsutsugamushi). PLoS Negl. Trop. Dis. 2015, 9, e0003971. [Google Scholar] [CrossRef] [PubMed]
- Bonell, A.; Lubell, Y.; Newton, P.N.; Crump, J.A.; Paris, D.H. Estimating the burden of scrub typhus: A systematic review. PLoS Negl. Trop. Dis. 2017, 11, e0005838. [Google Scholar] [CrossRef] [Green Version]
- McGready, R.; Prakash, J.A.; Benjamin, S.J.; Watthanaworawit, W.; Anantatat, T.; Tanganuchitcharnchai, A.; Ling, C.L.; Tan, S.O.; Ashley, E.A.; Pimanpanarak, M.; et al. Pregnancy outcome in relation to treatment of murine typhus and scrub typhus infection: A fever cohort and a case series analysis. PLoS Negl. Trop. Dis. 2014, 8, e3327. [Google Scholar] [CrossRef] [Green Version]
- Watt, G.; Jongsakul, K.; Chouriyagune, C.; Paris, R. Differentiating dengue virus infection from scrub typhus in Thai adults with fever. Am. J. Trop. Med. Hyg. 2003, 68, 536–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valbuena, G.; Walker, D.H. Approaches to vaccines against Orientia tsutsugamushi. Front. Cell. Infect. Microbiol. 2012, 2, 170. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, S.; Richards, A.L. Scrub typhus vaccines: Past history and recent developments. Hum. Vaccines 2007, 3, 73–80. [Google Scholar] [CrossRef]
- Ridgway, R.L.; Oaks, S.C., Jr.; LaBarre, D.D. Laboratory animal models for human scrub typhus. Lab. Anim. Sci. 1986, 36, 481–485. [Google Scholar]
- Robinson, D.M.; Gan, E.; Chan, T.C.; Huxsoll, D.L. Clinical and immunologic responses of silvered leaf monkeys (Presbytis cristatus) to experimental reinfection with Rickettsia tsutsugamushi. J. Infect. Dis. 1981, 143, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.M.; Chan, T.C.; Huxsoll, D.L. Clinical response of silvered leaf monkeys (Presbytis cristatus) to infection with strains of Rickettsia tsutsugamushi virulent and avirulent for mice. J. Infect. Dis. 1976, 134, 193–197. [Google Scholar] [CrossRef]
- Shirai, A.; Montrey, R.D.; Werner, R.M.; Arimbalam, S.; Huxsoll, D.L. Clinical responses of silvered leaf monkeys to infection with selected strains of Rickettsia tsutsugamushi. J. Infect. Dis. 1979, 140, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Shirai, A.; Montrey, R.D.; Werner, R.M.; Arimbalam, S.; Huxsoll, D.L. Comparison of experimental Rickettsia tsutsugamushi infections in silvered leaf (Presbytis cristatus) and cynomolgus (Macaca fascicularis) monkeys. Jpn. J. Med. Sci. Biol. 1979, 32, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.M.; Gan, E.; Chan, T.C.; Huxsoll, D.L. Patterns of rickettsemia and antibody response in silvered leaf monkeys (Presbytis cristatus) after inoculation with virulent and avirulent strains of Rickettsia tsutsugamushi. J. Infect. Dis. 1977, 135, 664–668. [Google Scholar] [CrossRef]
- Walker, J.S.; Cadigan, F.C.; Vosdingh, R.A.; Chye, C.T. The silvered leaf-monkey of Malaysia, Presbytis cristatus: Disease model for human scrub typhus. J. Infect. Dis. 1973, 128, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.S.; Delacruz, E.C.; Abalos, R.M.; Tan, E.V.; Jiang, J.; Richards, A.L.; Eamsila, C.; Rodkvantook, W.; Myint, K.S. Clinical and histological features of inoculation site skin lesions in cynomolgus monkeys experimentally infected with Orientia tsutsugamushi. Vector Borne Zoonotic Dis. 2007, 7, 547–554. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, J.G.; Rice, R.M.; Jerrells, T.R. Development of antigen-specific cell-mediated immune responses after infection of cynomolgus monkeys (Macaca fascicularis) with Rickettsia tsutsugamushi. J. Infect. Dis. 1985, 152, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Heisey, G.B.; Gan, E.; Shirai, A.; Groves, M.G. Scrub Typhus antibody in cynomolgus monkeys (Macaca fascicularis) in Malaysia. Lab. Anim. Sci. 1981, 31, 289–291. [Google Scholar] [PubMed]
- Chattopadhyay, S.; Jiang, J.; Chan, T.C.; Manetz, T.S.; Chao, C.C.; Ching, W.M.; Richards, A.L. Scrub typhus vaccine candidate Kp r56 induces humoral and cellular immune responses in cynomolgus monkeys. Infect. Immun. 2005, 73, 5039–5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunyakumthorn, P.; Somponpun, S.J.; Im-Erbsin, R.; Anantatat, T.; Jenjaroen, K.; Dunachie, S.J.; Lombardini, E.D.; Burke, R.L.; Blacksell, S.D.; Jones, J.W.; et al. Characterization of the rhesus macaque (Macaca mulatta) scrub typhus model: Susceptibility to intradermal challenge with the human pathogen Orientia tsutsugamushi Karp. PLoS Negl. Trop. Dis. 2018, 12, e0006305. [Google Scholar] [CrossRef] [Green Version]
- Kotal, J.; Langhansova, H.; Lieskovska, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecky, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelar, J. Modulation of host immunity by tick saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, D.; Cantaert, T.; Misse, D. Aedes Mosquito Salivary Components and Their Effect on the Immune Response to Arboviruses. Front. Cell. Infect. Microbiol. 2020, 10, 407. [Google Scholar] [CrossRef]
- McCracken, M.K.; Gromowski, G.D.; Garver, L.S.; Goupil, B.A.; Walker, K.D.; Friberg, H.; Currier, J.R.; Rutvisuttinunt, W.; Hinton, K.L.; Christofferson, R.C.; et al. Route of inoculation and mosquito vector exposure modulate dengue virus replication kinetics and immune responses in rhesus macaques. PLoS Negl. Trop. Dis. 2020, 14, e0008191. [Google Scholar] [CrossRef] [Green Version]
- Banajee, K.H.; Embers, M.E.; Langohr, I.M.; Doyle, L.A.; Hasenkampf, N.R.; Macaluso, K.R. Amblyomma maculatum Feeding Augments Rickettsia parkeri Infection in a Rhesus Macaque Model: A Pilot Study. PLoS ONE 2015, 10, e0135175. [Google Scholar] [CrossRef]
- Dudley, D.M.; Newman, C.M.; Lalli, J.; Stewart, L.M.; Koenig, M.R.; Weiler, A.M.; Semler, M.R.; Barry, G.L.; Zarbock, K.R.; Mohns, M.S.; et al. Infection via mosquito bite alters Zika virus tissue tropism and replication kinetics in rhesus macaques. Nat Commun 2017, 8, 2096. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Chaisiri, K.; Xia, D.; Armstrong, S.D.; Fang, Y.; Donnelly, M.J.; Kadowaki, T.; McGarry, J.W.; Darby, A.C.; Makepeace, B.L. Genomes of trombidid mites reveal novel predicted allergens and laterally-transferred genes associated with secondary metabolism. GigaScience 2018, 7, giy127. [Google Scholar] [CrossRef] [Green Version]
- Shatrov, A.B.; Takahashi, M.; Noda, S.; Misumi, H. Stylostome organization in feeding Leptotrombidium larvae (Acariformes: Trombiculidae). Exp. Appl. Acarol. 2014, 64, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Lurchachaiwong, W.; Monkanna, T.; Leepitakrat, S.; Ponlawat, A.; Sattabongkot, J.; Schuster, A.L.; McCardle, P.W.; Richards, A.L. Variable clinical responses of a scrub typhus outbred mouse model to feeding by Orientia tsutsugamushi infected mites. Exp. Appl. Acarol. 2012, 58, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.J. Lyme borreliosis: A review of data on transmission time after tick attachment. Int. J. Gen. Med. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Des Vignes, F.; Piesman, J.; Heffernan, R.; Schulze, T.L.; Stafford, K.C., 3rd; Fish, D. Effect of tick removal on transmission of Borrelia burgdorferi and Ehrlichia phagocytophila by Ixodes scapularis nymphs. J. Infect. Dis. 2001, 183, 773–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadosaka, T.; Kimura, E. Electron microscopic observations of Orientia tsutsugamushi in salivary gland cells of naturally Infected Leptotrombidium pallidum larvae during feeding. Microbiol. Immunol. 2003, 47, 727–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, L.W.; Robinson, D.M.; Rapmund, G.; Walker, J.S.; Gan, E.; Ram, S. Distribution of Rickettsia tsutsugamushi in organs of Leptotrombidium (Leptotrombidium) fletcheri (Prostigmata: Trombiculidae). J. Med. Entomol. 1975, 12, 345–348. [Google Scholar] [CrossRef]
- Wright, J.D.; Hastriter, M.W.; Robinson, D.M. Observations on the ultrastructure and distribution of Rickettsia tsutsugamushi in naturally infected Leptotrombidium (Leptotrombidium) arenicola (Acari: Trombiculidae). J. Med. Entomol. 1984, 21, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Hase, T.; Roberts, L.W.; Hildebrandt, P.K.; Cavanaugh, D.C. Stylostome formation by Leptotrombidium mites (Acari: Trombiculidae). J. Parasitol. 1978, 64, 712–718. [Google Scholar] [CrossRef]
- Shatrov, A.B. Stylostome formation in trombiculid mites (Acariformes: Trombiculidae). Exp. Appl. Acarol. 2009, 49, 261–280. [Google Scholar] [CrossRef]
- Kramme, S.; An, V.L.; Khoa, N.D.; Trin, V.L.; Tannich, E.; Rybniker, J.; Fleischer, B.; Drosten, C.; Panning, M. Orientia tsutsugamushi bacteremia and cytokine levels in Vietnamese scrub typhus patients. J. Clin. Microbiol. 2009, 47, 586–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lurchachaiwong, W.; Chan, T.C.; Richards, A.L.; McCardle, W.; Schuster, A.L. Establishment of Orientia tsutsugamushi Lc-1 (Rickettsiales: Rickettsiaceae) infection in ICR outbred mice (Rodentia: Muridae) by needle challenge. J. Med. Entomol. 2014, 51, 658–660. [Google Scholar] [CrossRef] [PubMed]
- Lerdthusnee, K.; Khuntirat, B.; Leepitakrat, W.; Tanskul, P.; Monkanna, T.; Khlaimanee, N.; Inlao, I.; Kengluecha, A.; Mungviriya, S.; Chandranoi, K.; et al. Scrub typhus: Vector competence of Leptotrombidium chiangraiensis chiggers and transmission efficacy and isolation of Orientia tsutsugamushi. Ann. N. Y. Acad. Sci. 2003, 990, 25–35. [Google Scholar] [CrossRef]
- Lurchachaiwong, W.; McCardle, W.; Chan, T.C.; Schuster, A.L.; Richards, A.L. Development of an Orientia tsutsugamushi Lc-1 Murine Intraperitoneal Challenge Model for Scrub Typhus: Determination of Murine Lethal Dose (MuLD50), Tissue Bacterial Loads, and Clinical Outcomes. Vector Borne Zoonotic Dis. 2015, 15, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Takhampunya, R.; Tippayachai, B.; Promsathaporn, S.; Leepitakrat, S.; Monkanna, T.; Schuster, A.L.; Melendrez, M.C.; Paris, D.H.; Richards, A.L.; Richardson, J.H. Characterization based on the 56-Kda type-specific antigen gene of Orientia tsutsugamushi genotypes isolated from Leptotrombidium mites and the rodent host post-infection. Am. J. Trop. Med. Hyg. 2014, 90, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phasomkusolsil, S.; Tanskul, P.; Ratanatham, S.; Watcharapichat, P.; Phulsuksombati, D.; Frances, S.P.; Lerdthusnee, K.; Linthicum, K.J. Transstadial and transovarial transmission of Orientia tsutsugamushi in Leptotrombidium imphalum and Leptotrombidium chiangraiensis (Acari: Trombiculidae). J. Med. Entomol. 2009, 46, 1442–1445. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chan, T.C.; Temenak, J.J.; Dasch, G.A.; Ching, W.M.; Richards, A.L. Development of a quantitative real-time polymerase chain reaction assay specific for Orientia tsutsugamushi. Am. J. Trop. Med. Hyg. 2004, 70, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Paris, D.H.; Blacksell, S.D.; Aukkanit, N.; Newton, P.N.; Phetsouvanh, R.; Izzard, L.; Stenos, J.; Graves, S.R.; Day, N.P.; et al. Diversity of the 47-kD HtrA nucleic acid and translated amino acid sequences from 17 recent human isolates of Orientia. Vector Borne Zoon. Dis. 2013, 13, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Suwanabun, N.; Chouriyagune, C.; Eamsila, C.; Watcharapichat, P.; Dasch, G.A.; Howard, R.S.; Kelly, D.J. Evaluation of an enzyme-linked immunosorbent assay in Thai scrub typhus patients. Am. J. Trop. Med. Hyg. 1997, 56, 38–43. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Sex | Location for Chigger Placement | Age (Years) | Sign of Illness | ChiggerFeeding Time | Physical Condition | CBC | Serum Chemistry | Bacteremia |
|---|---|---|---|---|---|---|---|---|---|
| NHP-1 | Female | Inner ear | 16 | Moderate reduced appetite, transient anorexia, and loss weight by 10% on day 7 | 1 h | Normal | Normal | Normal | Yes |
| NHP-2 | Male | Inner ear | 16 | Normal | 1.5 h | Normal | Normal | Normal | No |
| NHP-3 | Male | Umbilicus | 15 | Moderate reduced appetite and transient mild diarrhea starting from day 8 until day 28 | 2 h | Normal | Normal | Normal | Yes |
| Control | Female | Inner ear | 15 | Normal | 1 h | Normal | Normal | Normal | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linsuwanon, P.; Wongwairot, S.; Auysawasdi, N.; Monkanna, T.; Richards, A.L.; Leepitakrat, S.; Sunyakumthorn, P.; Im-Erbsin, R.; Poole-Smith, K.; McCardle, P. Establishment of a Rhesus Macaque Model for Scrub Typhus Transmission: Pilot Study to Evaluate the Minimal Orientia tsutsugamushi Transmission Time by Leptotrombidium chiangraiensis Chiggers. Pathogens 2021, 10, 1028. https://doi.org/10.3390/pathogens10081028
Linsuwanon P, Wongwairot S, Auysawasdi N, Monkanna T, Richards AL, Leepitakrat S, Sunyakumthorn P, Im-Erbsin R, Poole-Smith K, McCardle P. Establishment of a Rhesus Macaque Model for Scrub Typhus Transmission: Pilot Study to Evaluate the Minimal Orientia tsutsugamushi Transmission Time by Leptotrombidium chiangraiensis Chiggers. Pathogens. 2021; 10(8):1028. https://doi.org/10.3390/pathogens10081028
Chicago/Turabian StyleLinsuwanon, Piyada, Sirima Wongwairot, Nutthanun Auysawasdi, Taweesak Monkanna, Allen L. Richards, Surachai Leepitakrat, Piyanate Sunyakumthorn, Rawiwan Im-Erbsin, Katie Poole-Smith, and Patrick McCardle. 2021. "Establishment of a Rhesus Macaque Model for Scrub Typhus Transmission: Pilot Study to Evaluate the Minimal Orientia tsutsugamushi Transmission Time by Leptotrombidium chiangraiensis Chiggers" Pathogens 10, no. 8: 1028. https://doi.org/10.3390/pathogens10081028
APA StyleLinsuwanon, P., Wongwairot, S., Auysawasdi, N., Monkanna, T., Richards, A. L., Leepitakrat, S., Sunyakumthorn, P., Im-Erbsin, R., Poole-Smith, K., & McCardle, P. (2021). Establishment of a Rhesus Macaque Model for Scrub Typhus Transmission: Pilot Study to Evaluate the Minimal Orientia tsutsugamushi Transmission Time by Leptotrombidium chiangraiensis Chiggers. Pathogens, 10(8), 1028. https://doi.org/10.3390/pathogens10081028