
Effect of Temperature on the Expression of Classical Enterotoxin Genes among Staphylococci Associated with Bovine Mastitis


Abstract
:1. Introduction
2. Results
2.1. Distribution of Classical SE Genes among Staphylococcal Isolates
2.2. Growth of Staphylococci in Milk
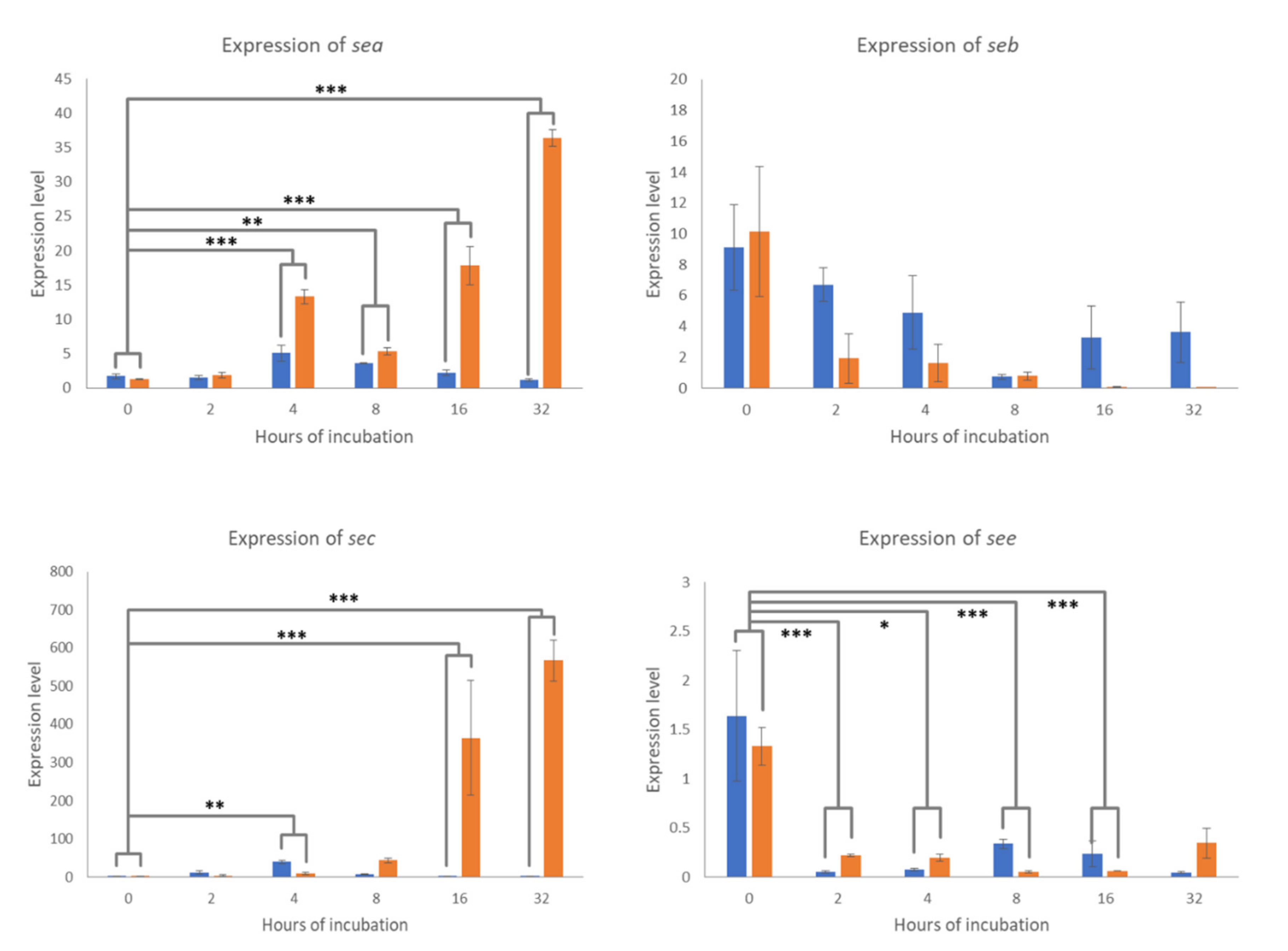
2.3. Expression of Classical SE Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Characterization of the Genus Staphylococcus
4.3. Genomic DNA Extraction
4.4. Genotypic Identification of Staphylococci
4.5. Detection of Classical SE Genes
4.6. Growth Curve Evaluation of Staphylococci in Milk
4.7. RNA Extraction and cDNA Synthesis
4.8. Quantitative Real-Time PCR (qPCR) Assay
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hennekinne, J.A.; De Buyser, M.L.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef] [Green Version]
- Podkowik, M.; Park, J.Y.; Seo, K.S.; Bystroń, J.; Bania, J. Enterotoxigenic Potential of Coagulase-Negative Staphylococci. Int. J. Food Microbiol. 2013, 163, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Park, J.Y.; Fox, L.K.; Seo, K.S.; McGuire, M.A.; Park, Y.H.; Rurangirwa, F.R.; Sischo, W.M.; Bohach, G.A. Detection of Classical and Newly Described Staphylococcal Superantigen Genes in Coagulase-Negative Staphylococci Isolated from Bovine Intramammary Infections. Vet. Microbiol. 2011, 147, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Fetsch, A.; Johler, S. Staphylococcus aureus as a foodborne pathogen. Curr. Clin. Microbiol. Rep. 2018, 5, 88–96. [Google Scholar] [CrossRef]
- Wieneke, A.A.; Roberts, D.; Gilbert, R.J. Staphylococcal Food Poisoning in the United Kingdom, 1969–90. Epidemiol. Infect. 1993, 110, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.P.; Barros, S.L.S.; Silva, V.C.; Cirqueira, M.G. Study of Staphylococcus aureus in raw and pasteurized milk consumed in the Reconcavo area of the state of Bahia, Brazil. J. Food Process. Technol. 2011, 02, 128. [Google Scholar] [CrossRef] [Green Version]
- Necidová, L.; Bogdanovičová, K.; Haruštiaková, D.; Bártová, K. Short communication: Pasteurization as a means of inactivating staphylococcal enterotoxins A, B, and C in milk. J. Dairy Sci. 2016, 99, 8638–8643. [Google Scholar] [CrossRef] [Green Version]
- Necidová, L.; Bursová, Š.; Haruštiaková, D.; Bogdanovičová, D.; Lačanin, I. Effect of heat treatment on activity of staphylococcal enterotoxins of type A, B, and C in milk. J. Dairy Sci. 2019, 102, 3924–3932. [Google Scholar] [CrossRef]
- Schmitt, M.; Schuler-Schmid, U.; Schmidt-Lorenz, W. Temperature Limits of Growth, TNase and Enterotoxin Production of Staphylococcus aureus Strains Isolated from Foods. Int. J. Food Microbiol. 1990, 11, 1–11. [Google Scholar] [CrossRef]
- Etter, D.; Schelin, J.; Schuppler, M.; Johler, S. Staphylococcal enterotoxin C-an update on SEC variants, their structure and properties, and their role in foodborne insecticides. Toxin 2020, 12, 584. [Google Scholar] [CrossRef] [PubMed]
- Bania, J.; Dabrowska, A.; Bystron, J.; Korzekwa, K.; Chrzanowska, J.; Molenda, J. Distribution of Newly Described Enterotoxin-Like genes in Staphylococcus aureus from food. Int. J. Food Microbiol. 2006, 108, 36–41. [Google Scholar] [CrossRef]
- Tremaine, M.T.; Brockman, D.K.; Betley, M.J. Staphylococcal Enterotoxin A Gene (Sea) Expression Is Not Affected by the Accessory Gene Regulator (Agr). Infect. Immun. 1993, 61, 356–359. [Google Scholar] [CrossRef] [Green Version]
- Srednik, M.E.; Usongo, V.; Lépine, S.; Janvier, X.; Archambault, M.A.; Gentilini, E.R. Characterization of Staphylococcus aureus strains isolated from mastitis bovine milk in Argentina. J. Dairy Res. 2018, 85, 57–63. [Google Scholar] [CrossRef]
- Boss, R.; Cosandey, A.; Luini, M.; Artursson, K.; Bardiau, M.; Breitenwieser, F.; Hehenberger, E.; Lam, T.; Mansfeld, M.; Michel, A.; et al. Bovine Staphylococcus aureus: Subtyping, Evolution, and Zoonotic Transfer. J. Dairy Sci. 2016, 99, 515–528. [Google Scholar] [CrossRef] [Green Version]
- De Freitas Guimarães, F.; Nóbrega, D.B.; Richini-Pereira, V.B.; Marson, P.M.; de Figueiredo Pantoja, J.C.; Langoni, H. Enterotoxin genes in coagulase-negative and coagulase-positive staphylococci isolated from bovine milk. J. Dairy Sci. 2013, 96, 2866–2872. [Google Scholar] [CrossRef] [Green Version]
- Piechota, M.; Kot, B.; Zdunek, E.; Mitrus, J.; Wicha, J.; Wolska, M.K.; Sachanowicz, K. Distribution of Classical Enterotoxin Genes in Staphylococci from Milk of Cows with- and Without Mastitis and the Cowshed Environment. Pol. J. Vet. Sci. 2014, 17, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Rall, V.L.M.; Sforcin, J.M.; de Deus, M.F.R.; de Sousa, D.C.; Camargo, C.H.; Godinho, N.C.; Galindo, L.A.; Soares, T.C.; Araújo, J.P. Polymerase Chain Reaction Detection of Enterotoxins Genes in Coagulase-Negative Staphylococci Isolated from Brazilian Minas Cheese. Foodborne Pathog. Dis. 2010, 7, 1121–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernozy-Rozand, C.; Mazuy, C.; Prevost, G.; Lapeyre, C.; Bes, M.; Brun, Y.; Fleurette, J. Enterotoxin Production by Coagulase-Negative Staphylococci Isolated from Goats’ Milk and Cheese. Int. J. Food Microbiol. 1996, 30, 271–280. [Google Scholar] [CrossRef]
- Bautista, L.; Gaya, P.; Medina, M.; Nuñez, M. A quantitative study of enterotoxin production by sheep milk staphylococci. Appl. Environ. Microbiol. 1988, 54, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; Keller, B.; von Eiff, C.; Brück, M.; Lubritz, G.; Etienne, J.; Peters, G. Enterotoxigenic Potential of Staphylococcus intermedius. Appl. Environ. Microbiol. 2001, 67, 5551–5557. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, H.J.; Mørk, T.; Høgåsen, H.R.; Rørvik, L.M. enterotoxigenic Staphylococcus aureus in bulk milk in Norway. J. Appl. Microbiol. 2005, 99, 158–166. [Google Scholar] [CrossRef]
- Tollersrud, T.; Kenny, K.; Reitz, A.J.; Lee, J.C. Genetic and Serologic Evaluation of Capsule Production by Bovine Mammary Isolates of Staphylococcus aureus and Other Staphylococcus spp. from Europe and the United States. J. Clin. Microbiol. 2000, 38, 2998–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günaydın, B.; Aslantaş, Ö.; Demir, C. Detection of superantigenic toxin genes in Staphylococcus aureus strains from subclinical bovine mastitis. Trop. Anim. Health Prod. 2011, 43, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Boynukara, B.; Gulhan, T.; Alisarli, M.; Gurturk, K.; Solmaz, H. Classical enterotoxigenic characteristics of Staphylococcus aureus strains isolated from bovine subclinical mastitis in Van, Turkey. Int. J. Food Microbiol. 2008, 125, 209–211. [Google Scholar] [CrossRef]
- Diab, M.S.; Ibrahim, N.A.; Elnaker, Y.F.; Zidan, S.A.; Saad, M.A. Molecular detection of Staphylococcus aureus enterotoxin genes isolated from mastitic milk and humans in El-Behira, Egypt. Int. J. One Health 2021, 7, 70–77. [Google Scholar] [CrossRef]
- Fang, R.; Cui, J.; Cui, T.; Guo, H.; Ono, H.K.; Park, C.H.; Okamura, M.; Nakane, A.; Hu, D.L. Staphylococcal Enterotoxin C Is an Important Virulence Factor for Mastitis. Toxin 2019, 11, 141. [Google Scholar] [CrossRef] [Green Version]
- Babić, M.; Pajić, M.; Radinović, M.; Boboš, S.; Bulajić, S.; Nikolić, A.; Velebit, B. Effects of Temperature Abuse on the Growth and Staphylococcal Enterotoxin A Gene (Sea) Expression of Staphylococcus aureus in Milk. Foodborne Pathog. Dis. 2019, 16, 282–289. [Google Scholar] [CrossRef]
- Wallin-Carlquist, N.; Cao, R.; Márta, D.; Sant’Ana da Silva, A.; Schelin, J.; Rådström, P. Acetic acid increases the phage-encoded enterotoxin A expression in Staphylococcus aureus. BMC Microbiol. 2010, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Zeaki, N.; Wallin-Carlquist, N.; Skandamis, P.N.; Schelin, J.; Rådstrom, P. Elevated enterotoxin A expression and formation in Staphylococcus aureus and its association with prophage induction. Appl. Environ. Microbiol. 2012, 78, 4942–4948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeaki, N.; Susilo, Y.B.; Pregiel, A.; Rådstrom, P.; Schelin, J. Prophage-encoded staphylococcal enterotoxin A: Regulation of production in Staphylococcus aureus strains representing different sea regions. Toxins 2015, 7, 5359–5376. [Google Scholar] [CrossRef]
- Schelin, J.; Wallin-Carlquist, N.; Cohn, M.T.; Lindqvist, R.; Barker, G.C.; Rådstrom, P. The formation of Staphylococcus aureus enterotoxin in food environments and advances in risk assessment. Virulence 2011, 2, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Tsutsuura, S.; Shimamura, Y.; Murata, M. Temperature dependence of the production of staphylococcal enterotoxin A by Staphylococcus aureus. Biosci. Biotechnol. Biochem. 2013, 77, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valihrach, L.; Alibayov, B.; Zdenkova, K.; Demnerova, K. Expression and production of staphylococcal enterotoxin C is substantially reduced in milk. Food Microbiol. 2014, 44, 54–59. [Google Scholar] [CrossRef]
- Hunt, K.; Butler, F.; Jordan, K. Factors affecting staphylococcal enterotoxin Cbovine production in milk. Int. Dairy J. 2014, 39, 41–46. [Google Scholar] [CrossRef]
- Edwards, V.M.; Deringer, J.R.; Callantine, S.D.; Deobald, C.F.; Berger, P.H.; Kapur, V.; Stauffacher, C.V.; Bohach, G.A. Characterization of the canine type C enterotoxin produced by Staphylococcus intermedius pyoderma isolates. Infect. Immun. 1997, 65, 2346–2352. [Google Scholar] [CrossRef] [Green Version]
- Marr, J.C.; Lyon, J.D.; Roberson, J.R.; Lupher, M.; Davis, W.C.; Bohach, G.A. Characterization of novel type C staphylococcal enterotoxins: Biological and evolutionary implications. Infect. Immun. 1993, 61, 4254–4262. [Google Scholar] [CrossRef] [Green Version]
- Valihrach, L.; Alibayov, B.; Demnerova, K. Production of staphylococcal enterotoxin C in milk. Int. Dairy J. 2013, 30, 103–107. [Google Scholar] [CrossRef]
- Alibayov, B.; Zdenkova, K.; Sykorova, H.; Demnerova, K. Molecular analysis of Staphylococcus aureus pathogenicity islands (SaPI) and their superantigens combination of food samples. J. Microbiol. Methods 2014, 107, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P. Mobile genetic elements and bacterial toxins: The Superantigen-Encoding Pathogenicity islands of Staphylococcus aureus. Plasmid 2003, 49, 93–105. [Google Scholar] [CrossRef]
- Novick, R.P.; Schlievert, P.; Ruzin, A. Pathogenicity and resistance islands of staphylococci. Microbes Infect. 2001, 3, 585–594. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet Mol Res. 2003, 2, 63–76. [Google Scholar]
- Fraser, J.D.; Proft, T. The bacterial superantigen and superantigen-like proteins. Immunol. Rev. 2008, 225, 226–243. [Google Scholar] [CrossRef] [PubMed]
- Derzelle, S.; Dilasser, F.; Duquenne, M.; Deperrois, V. Differential temporal expression of the staphylococcal enterotoxins genes during cell growth. Food Microbiol. 2009, 26, 896–904. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Jeggo, M. The one health approach—why is it so important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Mastitis Council. Laboratory Handbook on Bovine Mastitis, 3rd ed.; National Mastitis Council: Verona, WI, USA, 2017; pp. 43–52. [Google Scholar]
- Ciftci, A.; Findik, A.; Onuk, E.E.; Savasan, S. Detection of methicillin resistance and slime factor production of Staphylococcus aureus in bovine mastitis. Braz. J. Microbiol. 1958, 40, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. Optimization of the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Russell, V.; Lenth. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The aligned rank transform for nonparametric factorial analyses using only ANOVA procedures. In Proceedings of the ACM Conference on Human Factors in Computing Systems (CHI ‘11), Vancouver, BC, Canada, 7–12 May 2011; ACM Press: New York, NY, USA; pp. 143–146. [Google Scholar]
- Elkin, L.A.; Kay, M.; Higgins, J.; Wobbrock, J.O. An aligned rank transform procedure for multifactor contrast tests. In Proceedings of the ACM Symposium on User Interface Software and Technology (UIST ‘21), New York, NY, USA, 23 February 2021; ACM Press: New York, NY, USA Epub ahead of printing. [Google Scholar]
- R Core Team, 2018. version 3.5.0. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org (accessed on 5 July 2021).


{kind=link}
| Identification (n) | Number of Isolates without Detected Gene | Number of Isolates Carrying Gene | |||||
|---|---|---|---|---|---|---|---|
| sea | seb | sec | sed | see | sea + sec | ||
| S. aureus (51) | 33 | – | – | 15 | – | – | 3 |
| S. chomogenes (24) | 21 | – | – | 3 | – | – | – |
| S. haemolyticus (7) | 6 | – | – | – | – | 1 | – |
| S. epidermidis (5) | 3 | – | 1 | – | – | 1 | – |
| Other CNS (11) | 11 | – | – | – | – | – | – |
| Total (98) | 74 | – | 1 | 18 | – | 2 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homsombat, T.; Boonyayatra, S.; Awaiwanont, N.; Pichpol, D. Effect of Temperature on the Expression of Classical Enterotoxin Genes among Staphylococci Associated with Bovine Mastitis. Pathogens 2021, 10, 975. https://doi.org/10.3390/pathogens10080975
Homsombat T, Boonyayatra S, Awaiwanont N, Pichpol D. Effect of Temperature on the Expression of Classical Enterotoxin Genes among Staphylococci Associated with Bovine Mastitis. Pathogens. 2021; 10(8):975. https://doi.org/10.3390/pathogens10080975
Chicago/Turabian StyleHomsombat, Theeyathart, Sukolrat Boonyayatra, Nattakarn Awaiwanont, and Duangporn Pichpol. 2021. "Effect of Temperature on the Expression of Classical Enterotoxin Genes among Staphylococci Associated with Bovine Mastitis" Pathogens 10, no. 8: 975. https://doi.org/10.3390/pathogens10080975
APA StyleHomsombat, T., Boonyayatra, S., Awaiwanont, N., & Pichpol, D. (2021). Effect of Temperature on the Expression of Classical Enterotoxin Genes among Staphylococci Associated with Bovine Mastitis. Pathogens, 10(8), 975. https://doi.org/10.3390/pathogens10080975