
Adaptive Interactions of Achromobacter spp. with Pseudomonas aeruginosa in Cystic Fibrosis Chronic Lung Co-Infection

 ,
, 
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
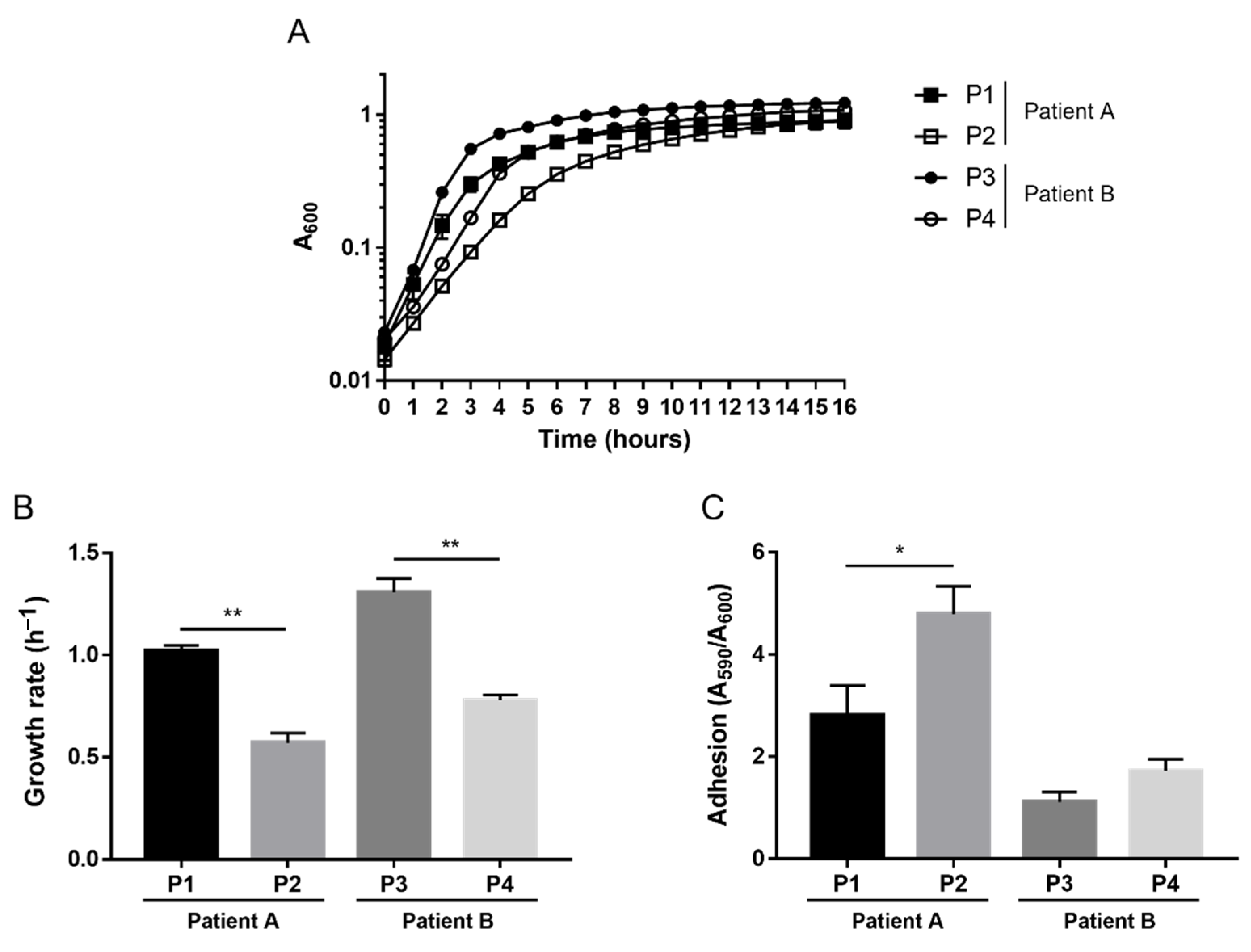
2.1. Phenotypic Variations
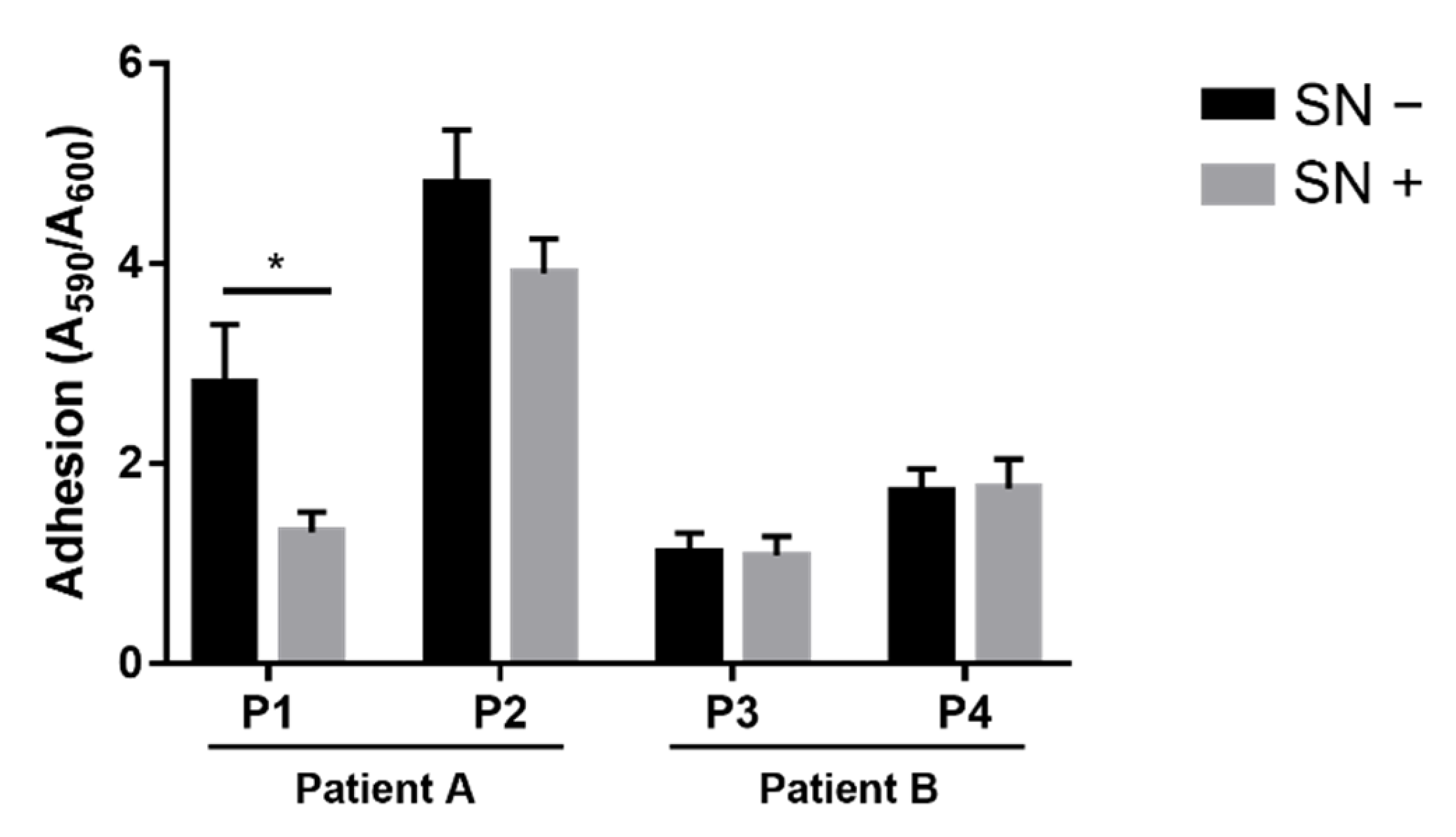
2.2. Effects of Achromobacter spp. on P. aeruginosa Adhesion
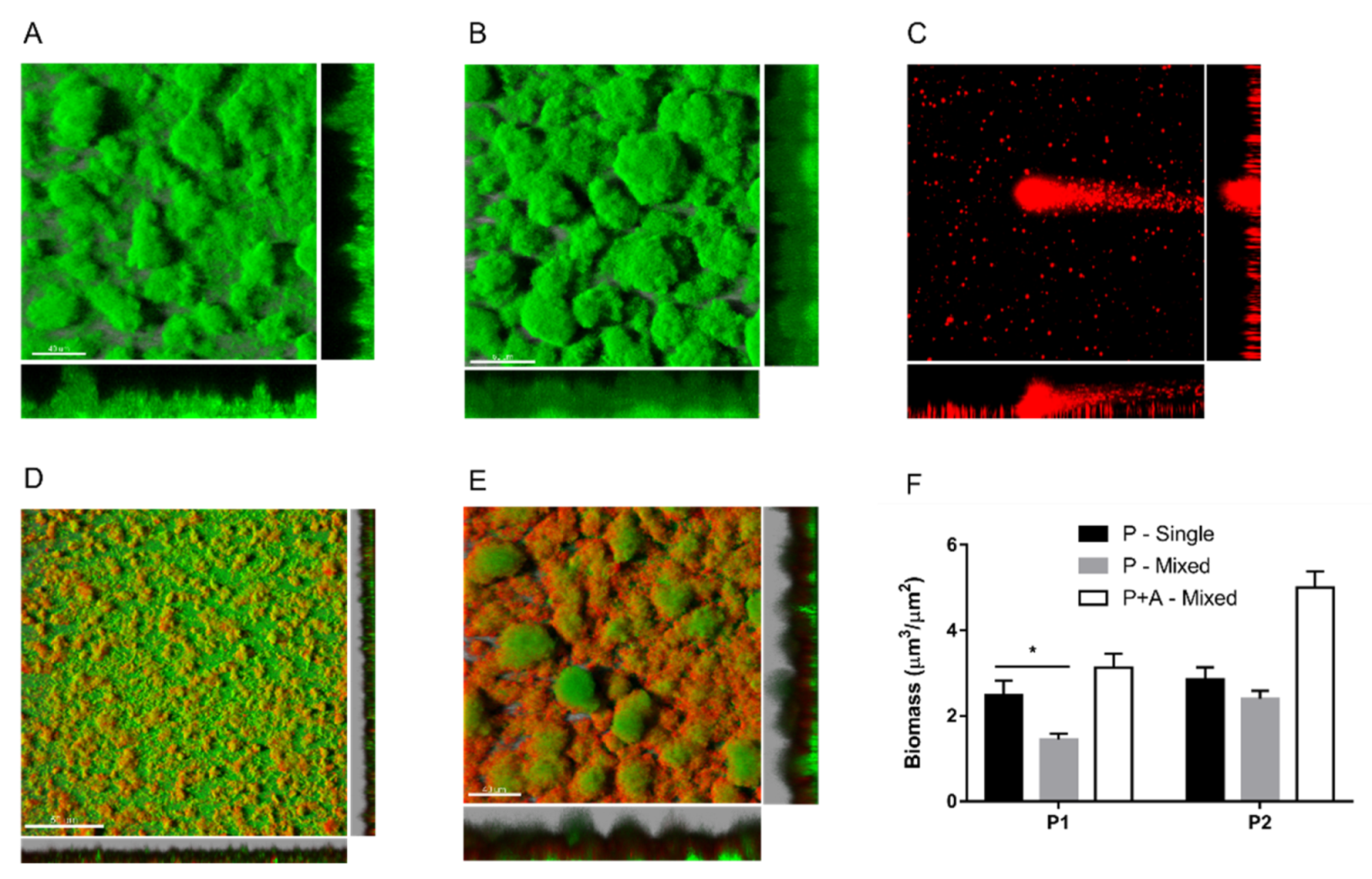
2.3. Effects of Achromobacter sp. on P. aeruginosa Biofilm Formation
2.4. Genetic Variants in Achromobacter sp. Virulence Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Growth Curves
4.3. Culture Supernatant Collection
4.4. Adhesion Assay
4.5. GFP Tagging
4.6. Biofilm Formation Assay
4.7. Genomic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rowe, S.M.; Miller, S.; Sorscher, E.J. Cystic Fibrosis. N. Engl. J. Med. 2005, 352, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Pillarisetti, N.; Williamson, E.; Linnane, B.; Skoric, B.; Robertson, C.F.; Robinson, P.; Massie, J.; Hall, G.; Sly, P.; Stick, S.; et al. Infection, Inflammation, and Lung Function Decline in Infants with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2011, 184, 75–81. [Google Scholar] [CrossRef]
- Ciofu, O.; Hansen, C.R.; Høiby, N. Respiratory bacterial infections in cystic fibrosis. Curr. Opin. Pulm. Med. 2013, 19, 251–258. [Google Scholar] [CrossRef]
- Aanaes, K.; Rickelt, L.F.; Johansen, H.K.; Von Buchwald, C.; Pressler, T.; Høiby, N.; Jensen, P.Ø. Decreased mucosal oxygen tension in the maxillary sinuses in patients with cystic fibrosis. J. Cyst. Fibros. 2011, 10, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Hansen, C.; Pressler, T.; Nielsen, K.G.; Jensen, P.Ø.; Bjarnsholt, T.; Høiby, N. Inflammation in Achromobacter xylosoxidans infected cystic fibrosis patients. J. Cyst. Fibros. 2010, 9, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambiase, A.; Catania, M.R.; Del Pezzo, M.; Rossano, F.; Terlizzi, V.; Sepe, A.; Raia, V. Achromobacter xylosoxidans respiratory tract infection in cystic fibrosis patients. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 973–980. [Google Scholar] [CrossRef] [Green Version]
- Edwards, B.D.; Greysson-Wong, J.; Somayaji, R.; Waddell, B.; Whelan, F.J.; Storey, D.; Rabin, H.R.; Surette, M.G.; Parkins, M.D. Prevalence and Outcomes of Achromobacter Species Infections in Adults with Cystic Fibrosis: A North American Cohort Study. J. Clin. Microbiol. 2017, 55, 2074–2085. [Google Scholar] [CrossRef] [Green Version]
- Firmida, M.; Pereira, R.; Silva, E.; Marques, E.; Lopes, A. Clinical impact of Achromobacter xylosoxidans colonization/infection in patients with cystic fibrosis. Braz. J. Med. Biol. Res. 2016, 49, 5097. [Google Scholar] [CrossRef] [Green Version]
- Hansen, C.R.; Pressler, T.; Høiby, N.; Gormsen, M. Chronic infection with Achromobacter xylosoxidans in cystic fibrosis patients; a retrospective case control study. J. Cyst. Fibros. 2006, 5, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhu, Y.; Ma, Y.; Liu, F.; Lu, N.; Yang, X.; Luan, C.; Yi, Y.; Zhu, B. Genomic Insights into Intrinsic and Acquired Drug Resistance Mechanisms in Achromobacter xylosoxidans. Antimicrob. Agents Chemother. 2014, 59, 1152–1161. [Google Scholar] [CrossRef] [Green Version]
- Traglia, G.M.; Almuzara, M.; Merkier, A.K.; Adams, C.; Galanternik, L.; Vay, C.; Centron, D.; Ramírez, M.S. Achromobacter xylosoxidans: An Emerging Pathogen Carrying Different Elements Involved in Horizontal Genetic Transfer. Curr. Microbiol. 2012, 65, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Gade, S.S.; Nørskov-Lauritsen, N.; Ridderberg, W. Prevalence and species distribution of Achromobacter sp. cultured from cystic fibrosis patients attending the Aarhus centre in Denmark. J. Med. Microbiol. 2017, 66, 686–689. [Google Scholar] [CrossRef]
- Papalia, M.; Steffanowski, C.; Traglia, G.M.; Almuzara, M.; Martina, P.; Galanternik, L.; Vay, C.; Gutkind, G.; Ramírez, M.S.; Radice, M. Diversity of Achromobacter species recovered from patients with cystic fibrosis, in Argentina. Rev. Argent. Microbiol. 2020, 52, 13–18. [Google Scholar] [CrossRef]
- Spilker, T.; Vandamme, P.; LiPuma, J.J. Identification and distribution of Achromobacter species in cystic fibrosis. J. Cyst. Fibros. 2013, 12, 298–301. [Google Scholar] [CrossRef] [Green Version]
- Barrado, L.; Brañas, P.; Orellana, M.D.L.A.; Martínez, M.T.; García, G.; Otero, J.R.; Chaves, F. Molecular Characterization of Achromobacter Isolates from Cystic Fibrosis and Non-Cystic Fibrosis Patients in Madrid, Spain. J. Clin. Microbiol. 2013, 51, 1927–1930. [Google Scholar] [CrossRef] [Green Version]
- Amoureux, L.; Bador, J.; Zouak, F.B.; Chapuis, A.; de Curraize, C.; Neuwirth, C. Distribution of the species of Achromobacter in a French Cystic Fibrosis Centre and multilocus sequence typing analysis reveal the predominance of A. xylosoxidans and clonal relationships between some clinical and environmental isolates. J. Cyst. Fibros. 2016, 15, 486–494. [Google Scholar] [CrossRef] [Green Version]
- McGuigan, L.; Callaghan, M. The evolving dynamics of the microbial community in the cystic fibrosis lung. Environ. Microbiol. 2015, 17, 16–28. [Google Scholar] [CrossRef]
- Tashiro, Y.; Yawata, Y.; Toyofuku, M.; Uchiyama, H.; Nomura, N. Interspecies Interaction between Pseudomonas aeruginosa and Other Microorganisms. Microbes Environ. 2013, 28, 13–24. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R.V.D. Interactions among cariogenic bacterial species in oral biofilm. In The Battle against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2015; Volume 1, pp. 438–444. [Google Scholar]
- Kuramitsu, H.K.; He, X.; Lux, R.; Anderson, M.H.; Shi, W. Interspecies Interactions within Oral Microbial Communities. Microbiol. Mol. Biol. Rev. 2007, 71, 653–670. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Lieleg, O.; Caldara, M.; Baumgärtel, R.; Ribbeck, K. Mechanical robustness of Pseudomonas aeruginosa biofilms. Soft Matter 2011, 7, 3307–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS 2013, 121, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Lowery, C.A.; Park, J.; Gloeckner, C.; Meijler, M.M.; Mueller, R.S.; Boshoff, H.I.; Ulrich, R.L.; Barry, C.; Bartlett, D.H.; Kravchenko, V.V.; et al. Defining the Mode of Action of Tetramic Acid Antibacterials Derived from Pseudomonas aeruginosa Quorum Sensing Signals. J. Am. Chem. Soc. 2009, 131, 14473–14479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liu, Y.; Markussen, T.; Høiby, N.; Tolker-Nielsen, T.; Molin, S. Pattern differentiation in co-culture biofilms formed by Staphylococcus aureus and Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol. 2011, 62, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, L.R.; Déziel, E.; D’Argenio, D.A.; Lepine, F.; Emerson, J.; McNamara, S.; Gibson, R.L.; Ramsey, B.W.; Miller, S.I. Selection for Staphylococcus aureus small-colony variants due to growth in the presence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2006, 103, 19890–19895. [Google Scholar] [CrossRef] [Green Version]
- Hubert, D.; Réglier-Poupet, H.; Sermet-Gaudelus, I.; Ferroni, A.; Le Bourgeois, M.; Burgel, P.R.; Serreau, P.R.; Dusser, D.; Poyart, C.; Coste, J. Association between Staphylococcus aureus alone or combined with Pseudomonas aeruginosa and the clinical condition of patients with cystic fibrosis. J. Cyst. Fibros. 2013, 12, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Chattoraj, S.S.; Murthy, R.; Ganesan, S.; Goldberg, J.B.; Zhao, Y.; Hershenson, M.B.; Sajjan, U.S. Pseudomonas aeruginosa Alginate Promotes Burkholderia cenocepacia Persistence in Cystic Fibrosis Transmembrane Conductance Regulator Knockout Mice. Infect. Immun. 2010, 78, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Bragonzi, A.; Farulla, I.; Paroni, M.; Twomey, K.B.; Pirone, L.; Lorè, N.I.; Bianconi, I.; Dalmastri, C.; Ryan, R.; Bevivino, A. Modelling Co-Infection of the Cystic Fibrosis Lung by Pseudomonas aeruginosa and Burkholderia cenocepacia Reveals Influences on Biofilm Formation and Host Response. PLoS ONE 2012, 7, e52330. [Google Scholar] [CrossRef] [Green Version]
- El-Halfawy, O.; Valvano, M.A. Chemical Communication of Antibiotic Resistance by a Highly Resistant Subpopulation of Bacterial Cells. PLoS ONE 2013, 8, e68874. [Google Scholar] [CrossRef] [Green Version]
- Bakkal, S.; Robinson, S.M.; Ordonez, C.L.; Waltz, D.A.; Riley, M.A. Role of bacteriocins in mediating interactions of bacterial isolates taken from cystic fibrosis patients. Microbiology 2010, 156, 2058–2067. [Google Scholar] [CrossRef] [Green Version]
- De Baets, F.; Schelstraete, P.; Van Daele, S.; Haerynck, F.; Vaneechoutte, M. Achromobacter xylosoxidans in cystic fibrosis: Prevalence and clinical relevance. J. Cyst. Fibros. 2007, 6, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Tetart, M.; Wallet, F.; Kyheng, M.; Leroy, S.; Perez, T.; Le Rouzic, O.; Wallaert, B.; Prevotat, A. Impact of Achromobacter xylosoxidans isolation on the respiratory function of adult patients with cystic fibrosis. ERJ Open Res. 2019, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Menetrey, Q.; Dupont, C.; Chiron, R.; Jumas-Bilak, E.; Marchandin, H. High Occurrence of Bacterial Competition Among Clinically Documented Opportunistic Pathogens Including Achromobacter xylosoxidans in Cystic Fibrosis. Front. Microbiol. 2020, 11, 558160. [Google Scholar] [CrossRef] [PubMed]
- Veschetti, L.; Sandri, A.; Johansen, H.K.; Lleò, M.M.; Malerba, G. Hypermutation as an Evolutionary Mechanism for Achromobacter xylosoxidans in Cystic Fibrosis Lung Infection. Pathogens 2020, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Gabrielaite, M.; Nørskov-Lauritsen, N.; Nielsen, F.C.; Johansen, H.K.; Marvig, R.L. Genetic adaptation and transmission of Achromobacter in cystic fibrosis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Marvig, R.L.; Sommer, L.M.M.; Molin, S.; Johansen, H.K. Convergent evolution and adaptation of Pseudomonas aeruginosa within patients with cystic fibrosis. Nat. Genet. 2015, 47, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Ly, P.M.; Feldman, M.F. The Tip of the VgrG Spike Is Essential to Functional Type VI Secretion System Assembly in Acinetobacter baumannii. mBio 2020, 11, e02761-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Holland, I.B.; Schmitt, L. The Type 1 secretion pathway—The hemolysin system and beyond. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1843, 1629–1641. [Google Scholar] [CrossRef] [Green Version]
- Rabin, H.R.; Surette, M.G. The cystic fibrosis airway microbiome. Curr. Opin. Pulm. Med. 2012, 18, 622–627. [Google Scholar] [CrossRef]
- Lim, Y.W.; Evangelista, J.S.; Schmieder, R.; Bailey, B.; Haynes, M.; Furlan, M.; Maughan, H.; Edwards, R.; Rohwer, F.; Conrad, U.; et al. Clinical Insights from Metagenomic Analysis of Sputum Samples from Patients with Cystic Fibrosis. J. Clin. Microbiol. 2013, 52, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, A.P.; Lopes, S.; Pereira, M.O. Insights into Cystic Fibrosis Polymicrobial Consortia: The Role of Species Interactions in Biofilm Development, Phenotype, and Response to In-Use Antibiotics. Front. Microbiol. 2017, 7, 2146. [Google Scholar] [CrossRef] [Green Version]
- Trancassini, M.; Iebba, V.; Citerã, N.; Tuccio, V.; Magni, A.; Varesi, P.; De Biase, R.V.; Totino, V.; Santangelo, F.; Gagliardi, A.; et al. Outbreak of Achromobacter xylosoxidans in an Italian Cystic fibrosis center: Genome variability, biofilm production, antibiotic resistance, and motility in isolated strains. Front. Microbiol. 2014, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, T.H.; Hansen, M.A.; Jensen, P.Ø.; Hansen, L.; Riber, L.; Cockburn, A.; Kolpen, M.; Hansen, C.R.; Ridderberg, W.; Eickhardt, S.; et al. Complete Genome Sequence of the Cystic Fibrosis Pathogen Achromobacter xylosoxidans NH44784-1996 Complies with Important Pathogenic Phenotypes. PLoS ONE 2013, 8, e68484. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary forPseudomonas aeruginosabiofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Ridderberg, W.; Nielsen, S.M.; Nørskov-Lauritsen, N. Genetic Adaptation of Achromobacter sp. during Persistence in the Lungs of Cystic Fibrosis Patients. PLoS ONE 2015, 10, e0136790. [Google Scholar] [CrossRef] [Green Version]
- Dantam, J.; Subbaraman, L.N.; Jones, L. Adhesion of Pseudomonas aeruginosa, Achromobacter xylosoxidans, Delftia acidovorans, Stenotrophomonas maltophiliato contact lenses under the influence of an artificial tear solution. Biofouling 2020, 36, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.K.; Willcox, M. Adhesion of Stenotrophomonas maltophilia, Delftia acidovorans, and Achromobacter xylosoxidans to Contact Lenses. Eye Contact Lens Sci. Clin. Pract. 2018, 44, S120–S126. [Google Scholar] [CrossRef]
- Nielsen, S.M.; Nørskov-Lauritsen, N.; Bjarnsholt, T.; Meyer, R.L. Achromobacter Species Isolated from Cystic Fibrosis Patients Reveal Distinctly Different Biofilm Morphotypes. Microorganisms 2016, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, A.; Cantón, R.; Campo, P.; Baquero, F.; Blazquez, J. High Frequency of Hypermutable Pseudomonas aeruginosa in Cystic Fibrosis Lung Infection. Science 2000, 288, 1251–1253. [Google Scholar] [CrossRef]
- Sha, J.; Rosenzweig, J.A.; Kozlova, E.V.; Wang, S.; Erova, T.E.; Kirtley, M.L.; van Lier, C.J.; Chopra, A.K. Evaluation of the roles played by Hcp and VgrG type 6 secretion system effectors in Aeromonas hydrophila SSU pathogenesis. Microbiology 2013, 159, 1120–1135. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, A.L.; Racher, K.; Jamieson, L.; Blight, M.A.; Holland, I.B. Mutations in HlyD, Part of the Type 1 Translocator for Hemolysin Secretion, Affect the Folding of the Secreted Toxin. J. Bacteriol. 2005, 187, 7471–7480. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, J.; Duong, F.; Wandersman, C.; Murgier, M.; Lazdunski, A. The secretion genes of Pseudomonas aeruginosa alkaline protease are functionally related to those of Erwinia chrysanthemi proteases and Escherichia coli?-haemolysin. Mol. Microbiol. 1991, 5, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Veschetti, L.; Sandri, A.; Patuzzo, C.; Melotti, P.; Malerba, G.; Lleò, M.M. Mobilome Analysis of Achromobacter spp. Isolates from Chronic and Occasional Lung Infection in Cystic Fibrosis Patients. Microorganisms 2021, 9, 130. [Google Scholar] [CrossRef]
- Lambertsen, L.; Sternberg, C.; Molin, S. Mini-Tn7 transposons for site-specific tagging of bacteria with fluorescent proteins. Environ. Microbiol. 2004, 6, 726–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-H.; Schweizer, H.P. mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef]
- Tolker-Nielsen, T.; Sternberg, C. Growing and Analyzing Biofilms in Flow Chambers. Curr. Protoc. Microbiol. 2011, 21, 1B.2.1–1B.2.17. [Google Scholar] [CrossRef]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program comstat. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Patient | Isolate | Species | Year of Isolation | Other Characteristics |
|---|---|---|---|---|
| A | A1 | A. insuavis | 2005 | Hypermutator |
| A | P1 | P. aeruginosa | 2005 | DK08 clone type |
| A | A2 | A. insuavis | 2008 | |
| A | P2 | P. aeruginosa | 2008 | DK08 clone type |
| B | A3 * | A. xylosoxidans | 2008 | |
| B | P3 | P. aeruginosa | 2008 | |
| B | A4 * | A. xylosoxidans | 2014 | |
| B | P4 | P. aeruginosa | 2014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandri, A.; Haagensen, J.A.J.; Veschetti, L.; Johansen, H.K.; Molin, S.; Malerba, G.; Signoretto, C.; Boaretti, M.; Lleo, M.M. Adaptive Interactions of Achromobacter spp. with Pseudomonas aeruginosa in Cystic Fibrosis Chronic Lung Co-Infection. Pathogens 2021, 10, 978. https://doi.org/10.3390/pathogens10080978
Sandri A, Haagensen JAJ, Veschetti L, Johansen HK, Molin S, Malerba G, Signoretto C, Boaretti M, Lleo MM. Adaptive Interactions of Achromobacter spp. with Pseudomonas aeruginosa in Cystic Fibrosis Chronic Lung Co-Infection. Pathogens. 2021; 10(8):978. https://doi.org/10.3390/pathogens10080978
Chicago/Turabian StyleSandri, Angela, Janus Anders Juul Haagensen, Laura Veschetti, Helle Krogh Johansen, Søren Molin, Giovanni Malerba, Caterina Signoretto, Marzia Boaretti, and Maria M. Lleo. 2021. "Adaptive Interactions of Achromobacter spp. with Pseudomonas aeruginosa in Cystic Fibrosis Chronic Lung Co-Infection" Pathogens 10, no. 8: 978. https://doi.org/10.3390/pathogens10080978
APA StyleSandri, A., Haagensen, J. A. J., Veschetti, L., Johansen, H. K., Molin, S., Malerba, G., Signoretto, C., Boaretti, M., & Lleo, M. M. (2021). Adaptive Interactions of Achromobacter spp. with Pseudomonas aeruginosa in Cystic Fibrosis Chronic Lung Co-Infection. Pathogens, 10(8), 978. https://doi.org/10.3390/pathogens10080978