
Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology



Abstract
:1. Introduction
2. Results
2.1. Museum Specimen Survey
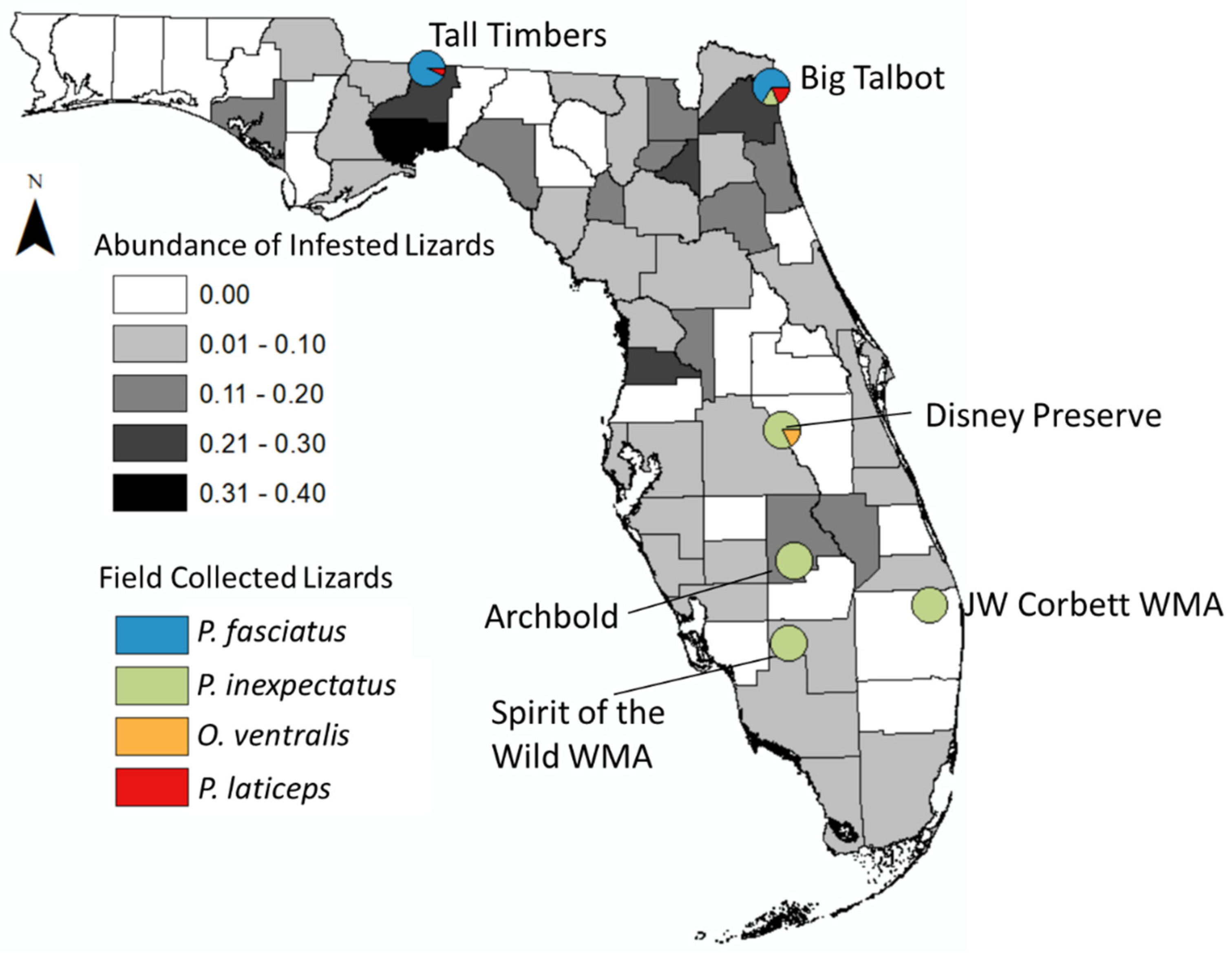
2.2. Field Collected Lizards and Ticks
2.3. Pathogen Screening
3. Discussion
4. Materials and Methods
4.1. Museum Samples
4.2. Field Sampling of Lizard and Ticks
4.3. PCR Testing and Sequencing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kugeler, K.J.; Schwartz, A.M.; Delorey, M.J.; Mead, P.S.; Hinckley, A.F. Estimating the Frequency of Lyme Disease Diagnoses, United States, 2010–2018. Emerg. Infect. Dis. 2021, 27, 616. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Updates on Borrelia burgdorferi sensu lato Complex with Respect to Public Health. Ticks Tick Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, I.; Yoshimizu, M.H.; Bonilla, D.L.; Fedorova, N.; Lane, R.S.; Padgett, K.A. Phylogeography of Borrelia Spirochetes in Ixodes pacificus and Ixodes spinipalpis Ticks Highlights Differential Acarological Risk of Tick-Borne Disease Transmission in Northern Versus Southern California. PLoS ONE 2019, 14, e0214726. [Google Scholar] [CrossRef]
- Wolcott, K.A.; Margos, G.; Fingerle, V.; Becker, N.S. Host Association of Borrelia burgdorferi sensu lato: A Review. Ticks Tick Borne Dis. 2021, 101766. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wielstra, B.; Rich, S.M. Northern and Southern Blacklegged (Deer) Ticks Are Genetically Distinct with Different Histories and Lyme Spirochete Infection Rates. Sci. Rep. 2020, 10, 10289. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Hickling, G.J.; Burke, R.L.; Ogden, N.H.; Beati, L.; LeBrun, R.A.; Arsnoe, I.M.; Gerhold, R.; Han, S.; Jackson, K.; et al. Why Lyme Disease Is Common in the Northern US, but Rare in the South: The Roles of Host Choice, Host-Seeking Behavior, and Tick Density. PLoS Biol. 2021, 19, e3001066. [Google Scholar] [CrossRef] [PubMed]
- Lehane, A.; Maes, S.E.; Graham, C.B.; Jones, E.; Delorey, M.; Eisen, R.J. Prevalence of Single and Coinfections of Human Pathogens in Ixodes ticks from Five Geographical Regions in the United States, 2013–2019. Ticks Tick Borne Dis. 2021, 12, 101637. [Google Scholar] [CrossRef]
- Johnson, T.L.; Graham, C.B.; Hojgaard, A.; Breuner, N.E.; Maes, S.E.; Boegler, K.A.; Eisen, R.J. Isolation of the Lyme Disease Spirochete Borrelia mayonii from Naturally Infected Rodents in Minnesota. J. Med. Entomol. 2017, 54, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Halsey, S.J.; Allan, B.F.; Miller, J.R. The Role of Ixodes scapularis, Borrelia burgdorferi and Wildlife Hosts in Lyme Disease Prevalence: A Quantitative Review. Ticks Tick Borne Dis. 2018, 9, 1103–1114. [Google Scholar] [CrossRef]
- Arsnoe, I.M.; Tsao, J.I.; Hickling, G.J. Nymphal Ixodes scapularis Questing Behavior Explains Geographic Variation in Lyme Borreliosis Risk in the Eastern United States. Ticks Tick Borne Dis. 2019, 10, 553–563. [Google Scholar] [CrossRef]
- Keirans, J.E.; Hutcheson, H.J.; Durden, L.A.; Klompen, J.S. Ixodes (Ixodes) scapularis (Acari: Ixodidae): Redescription of All Active Stages, Distribution, Hosts, Geographical Variation, and Medical and Veterinary Importance. J. Med. Entomol. 1996, 33, 297–318. [Google Scholar] [CrossRef]
- NieLin, G.; Kocan, A.A. Experimental Borrelia burgdorferi Infections in the White-Footed Mouse, Deer Mouse, and Fulvous Harvest Mouse Detected by Needle Aspiration of Spirochetes. J. Wildl. Dis. 1993, 29, 214–218. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.H., Jr.; Lin, T.; Gao, L.; Clark, K.L.; Banks, C.W.; Durden, L.A.; James, A.M.; Chandler, F.W., Jr. An Enzootic Transmission Cycle of Lyme Borreliosis Spirochetes in the Southeastern United States. Proc. Natl. Acad. Sci. USA 2003, 100, 11642–11645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, C.T.; Bursey, C.R.; Connior, M.B.; Durden, L.A.; Robison, H.W. Helminth and Arthropod Parasites of the Ground Skink, Scincella lateralis (Sauria: Scincidae), from Arkansas and Oklahoma, U.S.A. Comp. Parasitol. 2014, 81, 210–219. [Google Scholar] [CrossRef]
- Rogers, A.J. A Study of the Ixodid Ticks of Northern Florida, Including the Biology and Life History of Ixodes scapularis Say (Ixodidae: Acarina). Ph.D. Thesis, University of Maryland, College Park, MD, USA, 1953. [Google Scholar]
- Apperson, C.S.; Levine, J.F.; Evans, T.L.; Braswell, A.; Heller, J. Relative Utilization of Reptiles and Rodents as Hosts by Immature Ixodes scapularis (Acari: Ixodidae) in the Coastal Plain of North Carolina, USA. Exp. Appl. Acarol. 1993, 17, 719–731. [Google Scholar]
- Oliver, J.H., Jr.; Cummins, G.A.; Joiner, M.S. Immature Ixodes scapularis (Acari: Ixodidae) Parasitizing Lizards from the Southeastern U.S.A. J. Parasitol. 1993, 79, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Zikeli, S.; Zohdy, S. Why Research on Lyme Disease and Ixodes scapularis Needs Wildlife Ecology without Taxonomic Bias: A Review. Southeast. Nat. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Levine, J.F.; Apperson, C.S.; Howard, P.; Washburn, M.; Braswell, A.L. Lizards as Hosts for Immature Ixodes scapularis (Acari: Ixodidae) in North Carolina. J. Med. Entomol. 1997, 34, 594–598. [Google Scholar] [CrossRef]
- Durden, L.A.; Oliver, J.H., Jr.; Banks, C.W.; Vogel, G.N. Parasitism of Lizards by Immature Stages of the Blacklegged Tick, Ixodes scapularis (Acari, Ixodidae). Exp. Appl. Acarol. 2002, 26, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Quistad, G.B. Borreliacidal Factor in the Blood of the Western Fence Lizard (Sceloporus occidentalis). J. Parasitol. 1998, 84, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Hendricks, A.; Burge, D. Molecular Identification and Analysis of Borrelia burgdorferi sensu lato in Lizards in the Southeastern United States. Appl. Environ. Microbiol. 2005, 71, 2616–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.; Levine, J.F.; Yang, S.; Howard, P.; Apperson, C.S. Reservoir Competence of the Southeastern Five-Lined Skink (Eumeces inexpectatus) and the Green Anole (Anolis carolinensis) for Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1996, 54, 92–97. [Google Scholar] [CrossRef]
- Moody, T.D. Prevalence and Transmission Potential of Borrelia burgdorferi in Three Species of Wildcaught Plestiodon spp. Skinks of the Southeastern United States. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 2013. [Google Scholar]
- Krysko, K.L.; Burgess, J.P.; Rochford, M.R.; Gillette, C.R.; Cueva, D.; Enge, K.M.; Somma, L.A.; Stabile, J.L.; Smith, D.C.; Wasilewski, J.A.; et al. Verified Non-Indigenous Amphibians and Reptiles in Florida from 1863 through 2010: Outlining the Invasion Process and Identifying Invasion Pathways and Stages. Zootaxa 2011, 3028, 1–64. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Eisen, L.; Beard, C.B. County-Scale Distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the Continental United States. J. Med. Entomol. 2016, 53, 349–386. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Eisen, L.; Ogden, N.H.; Beard, C.B. Linkages of Weather and Climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), Enzootic Transmission of Borrelia Burgdorferi, and Lyme Disease in North America. J. Med. Entomol. 2016, 53, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Florida Department of Health. Annual Morbidity Statistics Report; Technical Report for Florida Department of Health; Florida Department of Health: Tallahassee, FL, USA, 2011–2017.
- Culbertson, K.A.; Herrmann, N.C. Asymmetric Interference Competition and Niche Partitioning Between Native and Invasive Anolis Lizards. Oecologia 2019, 190, 811–820. [Google Scholar] [CrossRef]
- Wehi, P.M.; Whaanga, H.; Trewick, S.A. Artefacts, Biology and Bias in Museum Collection Research. Mol. Ecol. 2012, 21, 3103–3109. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.M.; Gough, L. The Role of Temperature in Determining Distributions and Coexistence of Three Species of Plestiodon. J. Therm. Biol. 2012, 37, 374–379. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Albert, M.; Acevedo, L.; Dyer, M.C.; Arsnoe, I.M.; Tsao, J.I.; Mather, T.N.; LeBrun, R.A. Environmental Factors Affecting Survival of Immature Ixodes scapularis and Implications for Geographical Distribution of Lyme Disease: The Climate/Behavior Hypothesis. PLoS ONE 2017, 12, e0168723. [Google Scholar] [CrossRef] [PubMed]
- Arsnoe, I.M.; Hickling, G.J.; Ginsberg, H.S.; McElreath, R.; Tsao, J.I. Different Populations of Blacklegged Tick Nymphs Exhibit Differences in Questing Behavior that have Implications for Human Lyme Disease Risk. PLoS ONE. 2015, 10, e0127450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitt, L.J.; Cooper, W.E. Foraging and Diet of a Diurnal Predator (Eumeces laticeps) Feeding on Hidden Prey. J. Herpetol. 1986, 20, 408–415. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Neary, D.G.; Harris, L.D. Effect of High-Intensity Wildfire and Silvicultural Treatments on Reptile Communities in Sand-Pine Scrub. Conserv. Biol. 1994, 8, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Swei, A.; Ostfeld, R.S.; Lane, R.S.; Briggs, C.J. Impact of the Experimental Removal of Lizards on Lyme Disease Risk. Proc. Biol. Sci. 2011, 278, 2970–2978. [Google Scholar] [CrossRef]
- Slowik, T.J.; Lane, R.S. Feeding Preferences of the Immature Stages of Three Western North American Ixodid Ticks (Acari) for Avian, Reptilian, or Rodent Hosts. J. Med. Entomol. 2009, 46, 115–122. [Google Scholar] [CrossRef]
- James, A.M.; Oliver, J.H., Jr. Feeding and Host Preference of Immature Ixodes dammini, I. scapularis, and I. pacificus (Acari: Ixodidae). J. Med. Entomol. 1990, 27, 324–330. [Google Scholar] [CrossRef]
- De Jesus, C.E.; Ganser, C.; Kessler, W.H.; White, Z.S.; Bhosale, C.R.; Glass, G.E.; Wisely, S.M. A Survey of Tick-Borne Bacterial Pathogens in Florida. Insects 2019, 10, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giery, S.T.; Ostfeld, R.S. The Role of lizards in the Ecology of Lyme Disease in Two Endemic Zones of the Northeastern United States. J. Parasitol. 2007, 93, 511–517. [Google Scholar] [CrossRef]
- Rulison, E.L.; Kerr, K.T.; Dyer, M.C.; Han, S.; Burke, R.L.; Tsao, J.I.; Ginsberg, H.S. Minimal Role of Eastern Fence Lizards in Borrelia burgdorferi Transmission in Central New Jersey Oak/Pine Woodlands. J. Parasitol. 2014, 100, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.I.; Norris, D.E. Detection of Borrelia burgdorferi DNA in Lizards from Southern Maryland. Vector Borne Zoonotic Dis. 2007, 7, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Ostfeld, R.S.; Keesing, F. Biodiversity and Disease Risk: The Case of Lyme Disease. Conserv. Biol. 2000, 14, 722–728. [Google Scholar] [CrossRef]
- Pepin, K.M.; Eisen, R.J.; Mead, P.S.; Piesman, J.; Fish, D.; Hoen, A.G.; Barbour, A.G.; Hamer, S.; Diuk-Wasser, M.A. Geographic Variation in the Relationship between Human Lyme Disease Incidence and Density of Infected Host-Seeking Ixodes scapularis Nymphs in the Eastern United States. Am. J. Trop. Med. Hyg. 2012, 86, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.; Allgower, R.; Matuschka, F.R. Co-feeding transmission and its contribution to the perpetuation of the Lyme disease spirochete Borrelia afzelii. Emerg. Infect. Dis. 2003, 9, 895–896. [Google Scholar] [CrossRef]
- Nelder, M.P.; Russell, C.B.; Sheehan, N.J.; Sander, B.; Moore, S.; Li, Y.; Johnson, S.; Patel, S.N.; Sider, D. Human Pathogens Associated with the Blacklegged Tick Ixodes scapularis: A Systematic Review. Parasit. Vectors 2016, 9, 265. [Google Scholar] [CrossRef] [Green Version]
- McAllister, C.T.; Lance, A.; Durden, M.; Connior, B.; Henry, W. Robison Parasitism of Reptiles by the Blacklegged Tick (Ixodes scapularis) and Western Blacklegged Tick (Ixodes pacificus) with New Records of I. scapularis from Arkansas and Oklahoma Lizards: Implications for Lyme Disease Epidemiology. Herpetol. Rev. 2013, 44, 572–579. [Google Scholar]
- Durden, L.A.; Keirans, J.E. Nymphs of the Genus Ixodes (Acari: Ixodidae) of the United States: Taxonomy, Identification Key, Distribution, Hosts, and Medical/Veterinary Importance; Entomological Society of America: Lanham, MD, USA, 1996. [Google Scholar]
- Keirans, J.E.; Litwak, T.R. Pictorial Key to the Adults of Hard Ticks, Family Ixodidae (Ixodida: Ixodoidea), East of the Mississippi River. J. Med. Entomol. 1989, 26, 435–448. [Google Scholar] [CrossRef]
- Keirans, J.E.; Durden, L.A. Illustrated Key to Nymphs of the Tick Genus Amblyomma (Acari: Ixodidae) Found in the United States. J. Med. Entomol. 1998, 35, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Meshaka, W.E., Jr.; Layne, J.N. The Herpetology of Southern Florida. Herpetol. Conserv. Biol. 2015, 10, 161–171. [Google Scholar]
- Durden, L.A.; Dotson, E.M.; Vogel, G.N. Two Efficient Techniques for Catching Skinks. Herpetol. Rev. 1995, 26, 137. [Google Scholar]
- Barbour, A.G.; Maupin, G.O.; Teltow, G.J.; Carter, C.J.; Piesman, J. Identification of an Uncultivable Borrelia Species in the Hard Tick Amblyomma americanum: Possible Agent of a Lyme Disease-like Illness. J. Infect. Dis. 1996, 173, 403–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence Typing Reveals Extensive Strain Diversity of the Lyme Borreliosis Agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [Green Version]
- Tabara, K.; Arai, S.; Kawabuchi, T.; Itagaki, A.; Ishihara, C.; Satoh, H.; Okabe, N.; Tsuji, M. Molecular survey of Babesia microti, Ehrlichia species and Candidatus Neoehrlichia mikurensis in Wild Rodents from Shimane Prefecture, Japan. Microbiol. Immunol. 2007, 51, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic Identification of Rickettsiae and Estimation of Intraspecies Sequence Divergence for Portions of Two Rickettsial Genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Species | Avg. Larvae | Avg. Nymphs | Abundance of Infested Lizards (CI) |
|---|---|---|---|
| Anolis carolinensis (18) | 0 | 0 | 0/1241 (0%) (±0.004) |
| Aspidoscelis sexlineata (2) | 0 | 0 | 0/538 (0%) (±0.004) |
| Plestiodon egregious (10) | 0 | 1 | 1/966 (0.01%) (±0.01) |
| Plestiodon fasciatus (4) | 1.8 (±1.19) | 2.1 (±1.62) | 8/43 (18.6%) (±0.12) |
| Plestiodon inexpectatus (6) | 4.7 | 1.2 | 125/753 (16.6%) (±0.03) |
| Plestiodon laticeps (6) | 5.4 (±0.73) | 2.2 (±0.33) | 108/432 (25.0%) (±0.04) |
| Ophisaurus attenatus (7) | 19.5 | 1.5 | 1/102 (0.98%) (±0.03) |
| Ophisaurus compressus (13) | 0 | 0 | 0/89 (0%) (±0.04) |
| Ophisaurus ventralis (22) | 7.1 (±1.82) | 1.7 (±0.38) | 24/381 (6.2%) (±0.03) |
| Scincella lateralis (10) | 0.7 (±0.41) | 0.4 (±0.25) | 8/668 (1.2%) (±0.01) |
| Sceloporus undulatus (5) | 2.5 (±0.79) | 0.3 (±0.15) | 11/615 (1.7%) (±0.01) |
| Location | P. inexpectatus | P. fasciatus | P. laticeps | # Ticks Collected |
|---|---|---|---|---|
| North | ||||
| Big Talbot | 3/7 (42.9%) | 24/32 (75.0%) | 9/9 (100%) | 137 |
| Tall Timbers | 0 | 24/40 (60.0%) | 3/3 (100%) | 89 |
| Central | ||||
| Disney Preserve | 1/10 (10.0%) | - | - | 1 |
| Archbold | 4/16 (25.0%) | - | - | 5 |
| South | ||||
| JW Corbett WMA | 0/1 (0%) | - | - | 0 |
| Spirit of the Wild WMA | 1/4 (25.0%) | - | - | 1 |
| Total | 9/38 (23.7%) | 48/72 (66.7%) | 12/12 (100%) | 233 |
| Species | Sex | Avg. Larvae | Avg. Nymphs | Avg. All Ticks | Total Ticks | Abundance of Infested Lizards |
|---|---|---|---|---|---|---|
| P. fasciatus | Male | 0.7 (±0.56) | 3.4 (±0.56) | 4.1 (±0.68) | 74 | 18/25 (72.0%) (±0.17) |
| Female | 0.3 (±0.11) | 3.1 (±0.58) | 3.4 (±0.57) | 78 | 23/31 (74.2%) (±0.15) | |
| Juvenile | 0.1 (±0.14) | 0.7 (±0.14) | 1.0 (±0.0) | 7 | 7/16 (43.8%) (±0.22) | |
| P. inexpectatus | Male | 0 | 3.5 (±0.75) | 3.5 (±0.75) | 7 | 2/17 (11.8%) (±0.15) |
| Female | 0.3 (±0.33) | 1.3 (±0.33) | 1.7 (±0.33) | 5 | 3/10 (30.0%) (±0.16) | |
| Juvenile | 0 | 0.8 (±0.50) | 0.8 (±0.50) | 3 | 4/12 (33.3%) (±0.24) | |
| P. laticeps | Male | 0 | 2.5 (±1.11) | 2.5 (±1.11) | 15 | 5/5 (100%) (±0.22) |
| Female | 1.1 (±0.59) | 5.3 (±1.22) | 6.4 (±1.55) | 45 | 7/7 (100%) (±0.18) | |
| Juvenile | NA | NA | NA | NA | NA | |
| All P. spp. | 0.4 (±0.16) | 3.0 (±0.30) | 3.4 (±0.38) | 233 | 69/123 (56.1%) (±0.09) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Jesus, C.; Bhosale, C.; Wilson, K.; White, Z.; Wisely, S.M. Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology. Pathogens 2021, 10, 999. https://doi.org/10.3390/pathogens10080999
De Jesus C, Bhosale C, Wilson K, White Z, Wisely SM. Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology. Pathogens. 2021; 10(8):999. https://doi.org/10.3390/pathogens10080999
Chicago/Turabian StyleDe Jesus, Carrie, Chanakya Bhosale, Kristen Wilson, Zoe White, and Samantha M. Wisely. 2021. "Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology" Pathogens 10, no. 8: 999. https://doi.org/10.3390/pathogens10080999
APA StyleDe Jesus, C., Bhosale, C., Wilson, K., White, Z., & Wisely, S. M. (2021). Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology. Pathogens, 10(8), 999. https://doi.org/10.3390/pathogens10080999