
Characterization of a Nigerian Lumpy Skin Disease Virus Isolate after Experimental Infection of Cattle

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
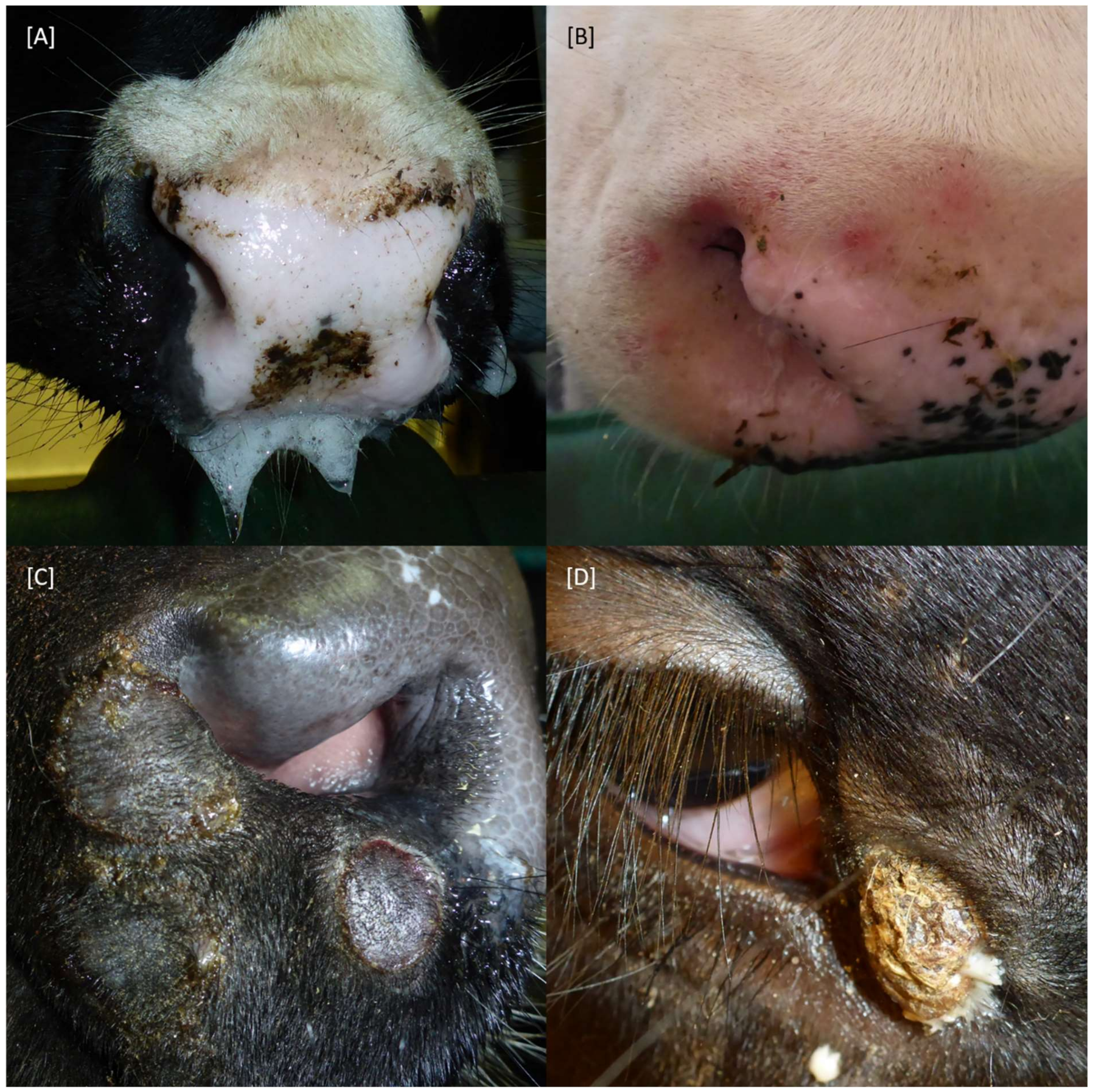
2.1. Clinical Signs
2.2. Viremia and Viral Shedding
2.3. Viral Genome Load in Selected Organ Samples
2.4. Serological Response
2.5. Phylogenetic Analysis of LSDV-V/281-Nigeria
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design and Sample Collection
4.3. Molecular Diagnostics
4.4. Serological Examination
4.5. Full-Length Genome Sequencing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2019 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 9 December 2020).
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An emerging worldwide threat to sheep, goats and cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, F.G. Lumpy skin disease, an African capripox virus disease of cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Davies, F.G. Lumpy skin disease of cattle: A growing problem in Africa and the Near East. World Anim. Rev. 1991, 68, 37–42. [Google Scholar]
- Carn, V.M.; Kitching, R.P. An investigation of possible routes of transmission of lumpy skin disease virus (Neethling). Epidemiol. Infect. 1995, 114, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of lumpy skin disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.; Mellor, P.S.; Coetzer, J.A.; Venter, E.H. A potential role for ixodid (hard) tick vectors in the transmission of lumpy skin disease virus in cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haig, D.A. Lumpy skin disease. Bull Epizoot Dis. Afr. 1957, 5, 421–430. [Google Scholar]
- Rouby, S.; Aboulsoud, E. Evidence of intrauterine transmission of lumpy skin disease virus. Vet. J. 2016, 209, 193–195. [Google Scholar] [CrossRef]
- Annandale, C.H.; Holm, D.E.; Ebersohn, K.; Venter, E.H. Seminal transmission of lumpy skin disease virus in heifers. Transbound. Emerg. Dis. 2014, 61, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Weiss, K.E. Lumpy skin disease virus. Virol. Monogr. 1968, 3, 111–131. [Google Scholar]
- Wolff, J.; Krstevski, K.; Beer, M.; Hoffmann, B. Minimum Infective Dose of a Lumpy Skin Disease Virus Field Strain from North Macedonia. Viruses 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M.; Kitching, R.P. The clinical response of cattle experimentally infected with lumpy skin disease (Neethling) virus. Arch. Virol. 1995, 140, 503–513. [Google Scholar] [CrossRef]
- Möller, J.; Moritz, T.; Schlottau, K.; Krstevski, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental lumpy skin disease virus infection of cattle: Comparison of a field strain and a vaccine strain. Arch. Virol. 2019, 164, 2931–2941. [Google Scholar] [CrossRef]
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Manning, L.; Neufeld, J.; Embury-Hyatt, C.; Copps, J.; Boyle, D.B. Quantification of lumpy skin disease virus following experimental infection in cattle. Transbound. Emerg. Dis. 2008, 55, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.; Oura, C.A. Review: Lumpy skin disease: An emerging threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A. The detection of lumpy skin disease virus in samples of experimentally infected cattle using different diagnostic techniques. Onderstepoort J. Vet. Res. 2005, 72, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Diesel, A.M. The epizootiology of lumpy skin disease in South Africa. In Proceedings of the 14th International Veterinary Congress, London, UK, 8 October 1949; Volume 2, pp. 492–500. [Google Scholar]
- Thomas, A.D.; Mare, C.v.E. Knopvelsiekte. J. S. Afr. Vet. Assoc. 1945, 16, 36–43. [Google Scholar]
- Woods, J.A. Lumpy skin disease—A review. Trop Anim. Health Prod. 1988, 20, 11–17. [Google Scholar] [CrossRef]
- Hunter, P.; Wallace, D. Lumpy skin disease in southern Africa: A review of the disease and aspects of control. J. South Afr. Vet. Assoc. 2001, 72, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. OIE-Listed Diseases, Infections and Infestations in Force in 2020. Available online: https://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2020/ (accessed on 16 October 2020).
- Beard, P.M. Lumpy skin disease: A direct threat to Europe. Vet. Rec. 2016, 178, 557–558. [Google Scholar] [CrossRef] [Green Version]
- Carn, V.M. Control of capripoxvirus infections. Vaccine 1993, 11, 1275–1279. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Antoniou, S.E.; Tsiamadis, E.; Topkaridou, M.; Labus, T.; Debeljak, Z.; Plavšić, B.; Miteva, A.; Alexandrov, T.; Pite, L.; et al. Field observations and experiences gained from the implementation of control measures against lumpy skin disease in South-East Europe between 2015 and 2017. Prev. Vet. Med. 2020, 181, 104600. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G.; Mbugwa, G. The alterations in pathogenicity and immunogenicity of a Kenya sheep and goat pox virus on serial passage in bovine foetal muscle cell cultures. J. Comp. Pathol. 1985, 95, 565–572. [Google Scholar] [CrossRef]
- van Rooyen, P.J.; Munz, E.K.; Weiss, K.E. The optimal conditions for the multiplication of Neethling-type lumpy skin disease virus in embryonated eggs. Onderstepoort J. Vet. Res. 1969, 36, 165–174. [Google Scholar]
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling lumpy skin disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of lumpy skin disease—The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842. [Google Scholar] [CrossRef]
- MacDonald, R.A.S. Pseudo-urticaria of cattle. In Annual Report for 1930–1931; Department of Animal Health, Government of Northern Rhodesia: Lusaka, Zambia, 1931; pp. 20–21. [Google Scholar]
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Bertagnoli, S.; Meyer, G.; Ganiere, J.P.; Caufour, P.; De Clercq, K.; Jacquiet, P.; Fournie, G.; Hautefeuille, C.; Etore, F.; et al. Risk of introduction of lumpy skin disease in France by the import of vectors in animal trucks. PLoS ONE 2018, 13, e0198506. [Google Scholar] [CrossRef]
- Adedeji, A.; Ja, A.; Dogonyaro, B.; Kujul, N.; Tekki, I.; Asala, O.; Chima, N.; Dyek, D.; Maguda, A. Recurrent Outbreaks of Lumpy Skin Disease and its Economic Impact on a Dairy Farm in Jos, Plateau State, Nigeria. Niger. Vet. J. 2017, 38, 151–158. [Google Scholar]
- Nawathe, D.R.; Asagba, M.O.; Abegunde, A.; Ajayi, S.A.; Durkwa, L. Some observations on the occurrence of lumpy skin disease in Nigeria. Zent. Vet. B 1982, 29, 31–36. [Google Scholar] [CrossRef]
- Nawathe, D.R.; Gibbs, E.P.; Asagba, M.O.; Lawman, M.J. Lumpyskin disease in Nigeria. Trop. Anim. Health Prod. 1978, 10, 49–54. [Google Scholar] [CrossRef]
- Bida, S.A.; Ali, I.I.; Eid, F.L.A.; Mustapha, H.B. Lumpy skin disease outbreak in cattle of Northern Nigeria. J. Niger. Vet. Med. Assoc. 1976, 5, 21–26. [Google Scholar]
- Adedeji, A.J.; Möller, J.; Meseko, C.A.; Adole, J.A.; Tekki, I.S.; Shamaki, D.; Hoffmann, B. Molecular characterization of Capripox viruses obtained from field outbreaks in Nigeria between 2000 and 2016. Transbound. Emerg. Dis. 2019, 66, 1631–1641. [Google Scholar] [CrossRef]
- Osuagwuh, U.I.; Bagla, V.; Venter, E.H.; Annandale, C.H.; Irons, P.C. Absence of lumpy skin disease virus in semen of vaccinated bulls following vaccination and subsequent experimental infection. Vaccine 2007, 25, 2238–2243. [Google Scholar] [CrossRef] [Green Version]
- Wolff, J.; Moritz, T.; Schlottau, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines 2020, 9, 4. [Google Scholar] [CrossRef]
- Kononov, A.; Prutnikov, P.; Shumilova, I.; Kononova, S.; Nesterov, A.; Byadovskaya, O.; Pestova, Y.; Diev, V.; Sprygin, A. Determination of lumpy skin disease virus in bovine meat and offal products following experimental infection. Transbound. Emerg. Dis. 2019, 66, 1332–1340. [Google Scholar] [CrossRef]
- Aleksandr, K.; Olga, B.; David, W.B.; Pavel, P.; Yana, P.; Svetlana, K.; Alexander, N.; Vladimir, R.; Dmitriy, L.; Alexander, S. Non-vector-borne transmission of lumpy skin disease virus. Sci. Rep. 2020, 10, 7436. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Campe, W.V.; Aerts, L.; Venter, E.; Tuppurainen, E.; Saegerman, C.; De Clercq, K. Comparative Evaluation of Lumpy Skin Disease Virus-Based Live Attenuated Vaccines. Vaccines 2021, 9, 473. [Google Scholar] [CrossRef]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical lumpy skin disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A.; Bell-Sakyi, L. Lumpy skin disease: Attempted propagation in tick cell lines and presence of viral DNA in field ticks collected from naturally-infected cattle. Ticks Tick Borne Dis. 2015, 6, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Wolff, J.; King, J.; Moritz, T.; Pohlmann, A.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental Infection and Genetic Characterization of Two Different Capripox Virus Isolates in Small Ruminants. Viruses 2020, 12, 1098. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Wolff, J.; Beer, M.; Hoffmann, B. Probe-Based Real-Time qPCR Assays for a Reliable Differentiation of Capripox Virus Species. Microorganisms 2021, 9, 765. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [Green Version]
- Dietze, K.; Moritz, T.; Alexandrov, T.; Krstevski, K.; Schlottau, K.; Milovanovic, M.; Hoffmann, D.; Hoffmann, B. Suitability of group-level oral fluid sampling in ruminant populations for lumpy skin disease virus detection. Vet. Microbiol. 2018, 221, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Spearman, C. The method of “right and wrong cases” (“constant stimuli”) without Gauss’s formulae. Br. J. Psychol. 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimentally Infected Animals | i-c Animals | |||||||
|---|---|---|---|---|---|---|---|---|
| Sample Material | R-81 | R-47 | R-84 | R-74 | R-41 | R-83 | R-32 | R-37 |
| Cervical lymph node | no Cq | no Cq | no Cq | 15.9 | 21.8 | no Cq | no Cq | no Cq |
| Mediastinal lymph node | 22.7 | 16.0 | ||||||
| Mesenterial lymph node | 37.3 | 28.3 | ||||||
| Liver | 29.5 | 21.8 | ||||||
| Spleen | 28.9 | 19.2 | ||||||
| Lung | no Cq | no Cq | 33.4 | 26.1 | 23.2 | 33.4 | no Cq | no Cq |
| Location of Skin Sample | ||||||||
| Anus | 25.3 | 17.8 | ||||||
| 24.6 | 16.3 | |||||||
| 24.2 | 18.5 | |||||||
| Muzzle | 22.4 | |||||||
| Subcutis | no Cq | |||||||
| 22.0 | ||||||||
| 23.6 | ||||||||
| Lung | 31.8 | |||||||
| 28.3 | ||||||||
| 27.1 | ||||||||
| 28.8 | ||||||||
| 28.4 | ||||||||
| Healed skin lesion (see Figure 2C) | 28.2 | |||||||
| 26.5 | ||||||||
| 28.5 | ||||||||
| 27.0 | ||||||||
| 34.0 | ||||||||
| Crust of skin lesion (see Figure 2D) | 19.7 | |||||||
| 20.2 | ||||||||
| 21.7 | ||||||||
| 18.3 | ||||||||
| 19.0 | ||||||||
| 18.1 | ||||||||
| 17.6 | ||||||||
| 21.4 | ||||||||
| 20.7 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolff, J.; Tuppurainen, E.; Adedeji, A.; Meseko, C.; Asala, O.; Adole, J.; Atai, R.; Dogonyaro, B.; Globig, A.; Hoffmann, D.; et al. Characterization of a Nigerian Lumpy Skin Disease Virus Isolate after Experimental Infection of Cattle. Pathogens 2022, 11, 16. https://doi.org/10.3390/pathogens11010016
Wolff J, Tuppurainen E, Adedeji A, Meseko C, Asala O, Adole J, Atai R, Dogonyaro B, Globig A, Hoffmann D, et al. Characterization of a Nigerian Lumpy Skin Disease Virus Isolate after Experimental Infection of Cattle. Pathogens. 2022; 11(1):16. https://doi.org/10.3390/pathogens11010016
Chicago/Turabian StyleWolff, Janika, Eeva Tuppurainen, Adeyinka Adedeji, Clement Meseko, Olayinka Asala, Jolly Adole, Rebecca Atai, Banenat Dogonyaro, Anja Globig, Donata Hoffmann, and et al. 2022. "Characterization of a Nigerian Lumpy Skin Disease Virus Isolate after Experimental Infection of Cattle" Pathogens 11, no. 1: 16. https://doi.org/10.3390/pathogens11010016
APA StyleWolff, J., Tuppurainen, E., Adedeji, A., Meseko, C., Asala, O., Adole, J., Atai, R., Dogonyaro, B., Globig, A., Hoffmann, D., Beer, M., & Hoffmann, B. (2022). Characterization of a Nigerian Lumpy Skin Disease Virus Isolate after Experimental Infection of Cattle. Pathogens, 11(1), 16. https://doi.org/10.3390/pathogens11010016