
Biophysical Manipulation of the Extracellular Environment by Eurotium halophilicum

 , , ,
, , ,
Abstract
:1. Introduction
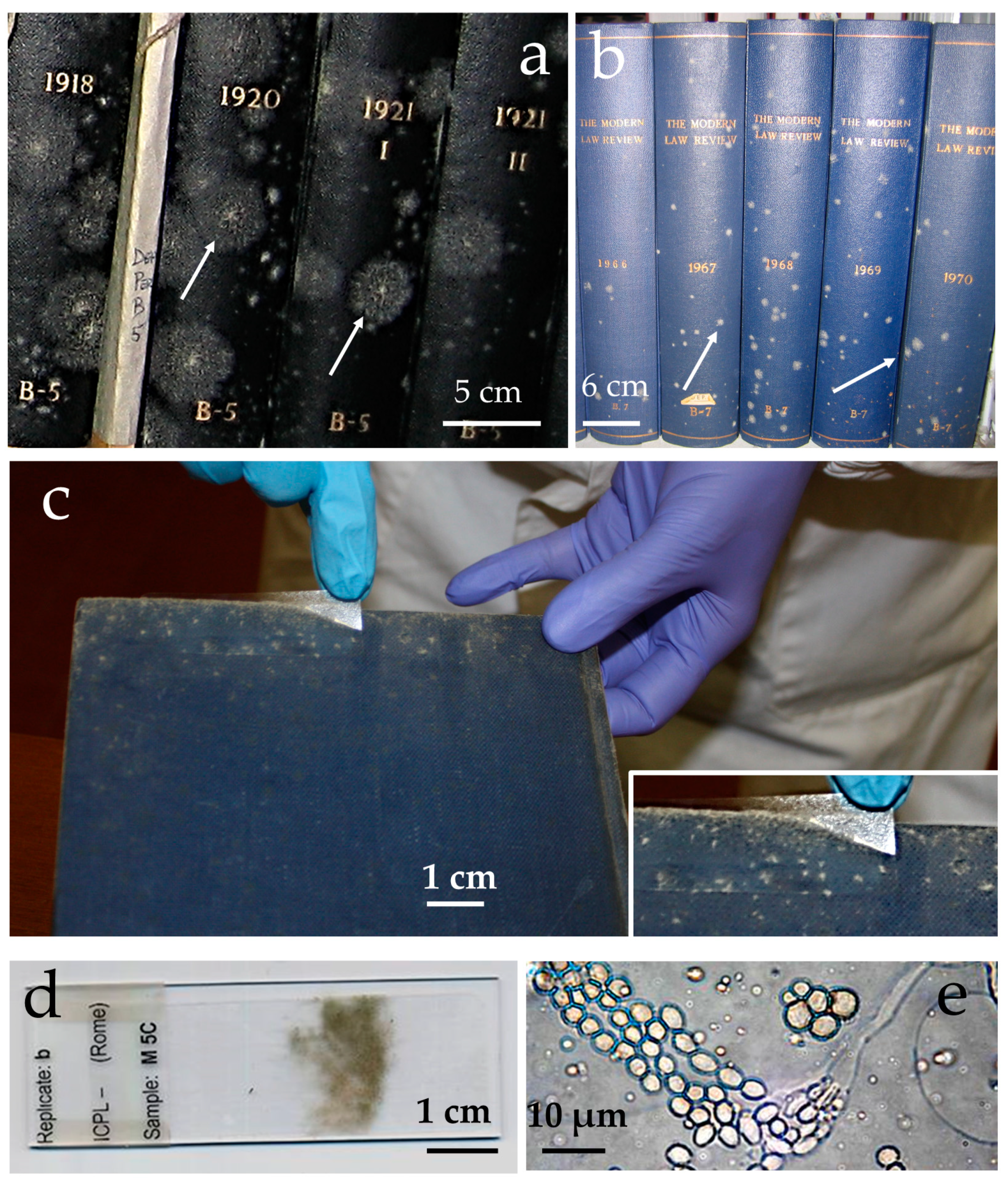
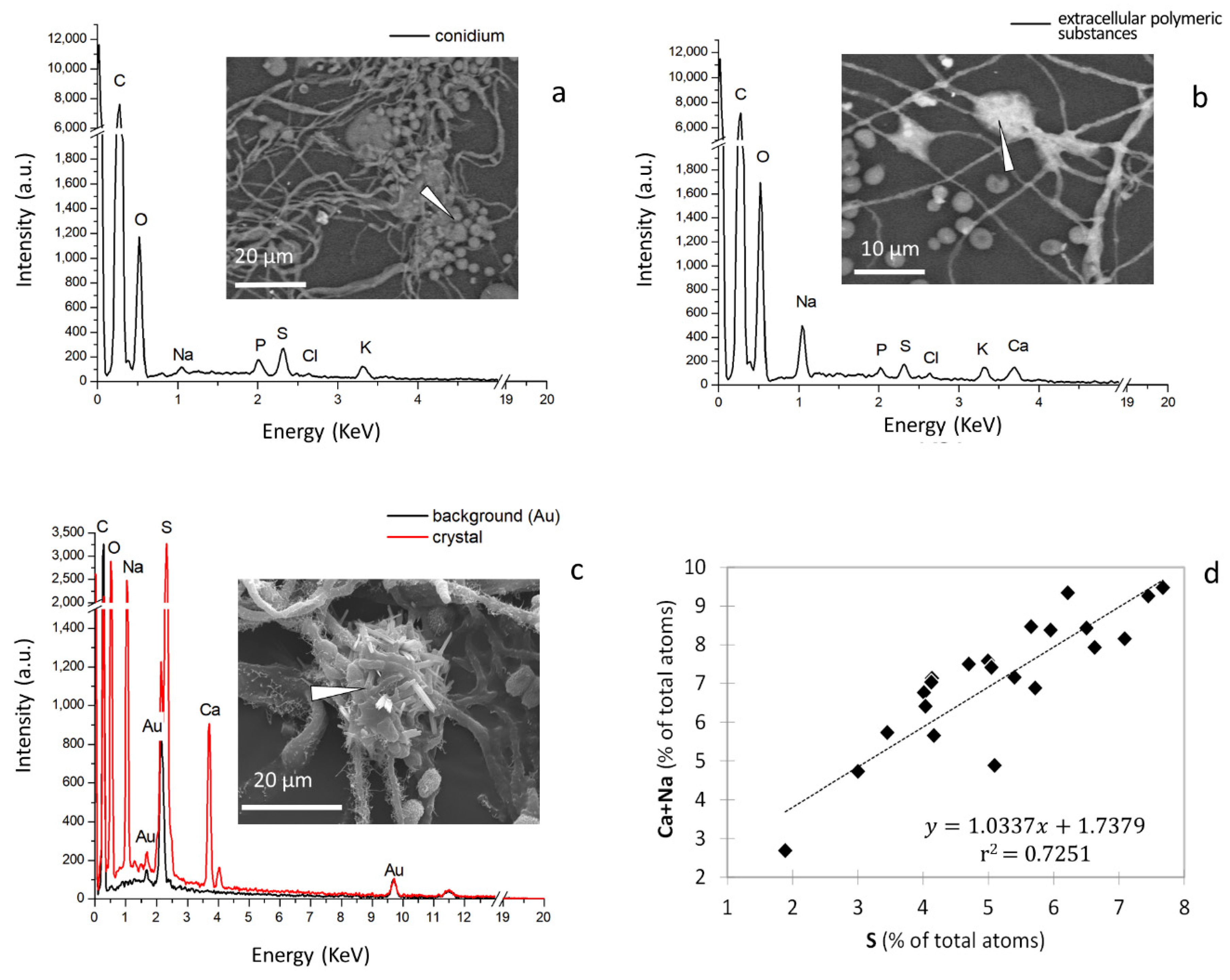
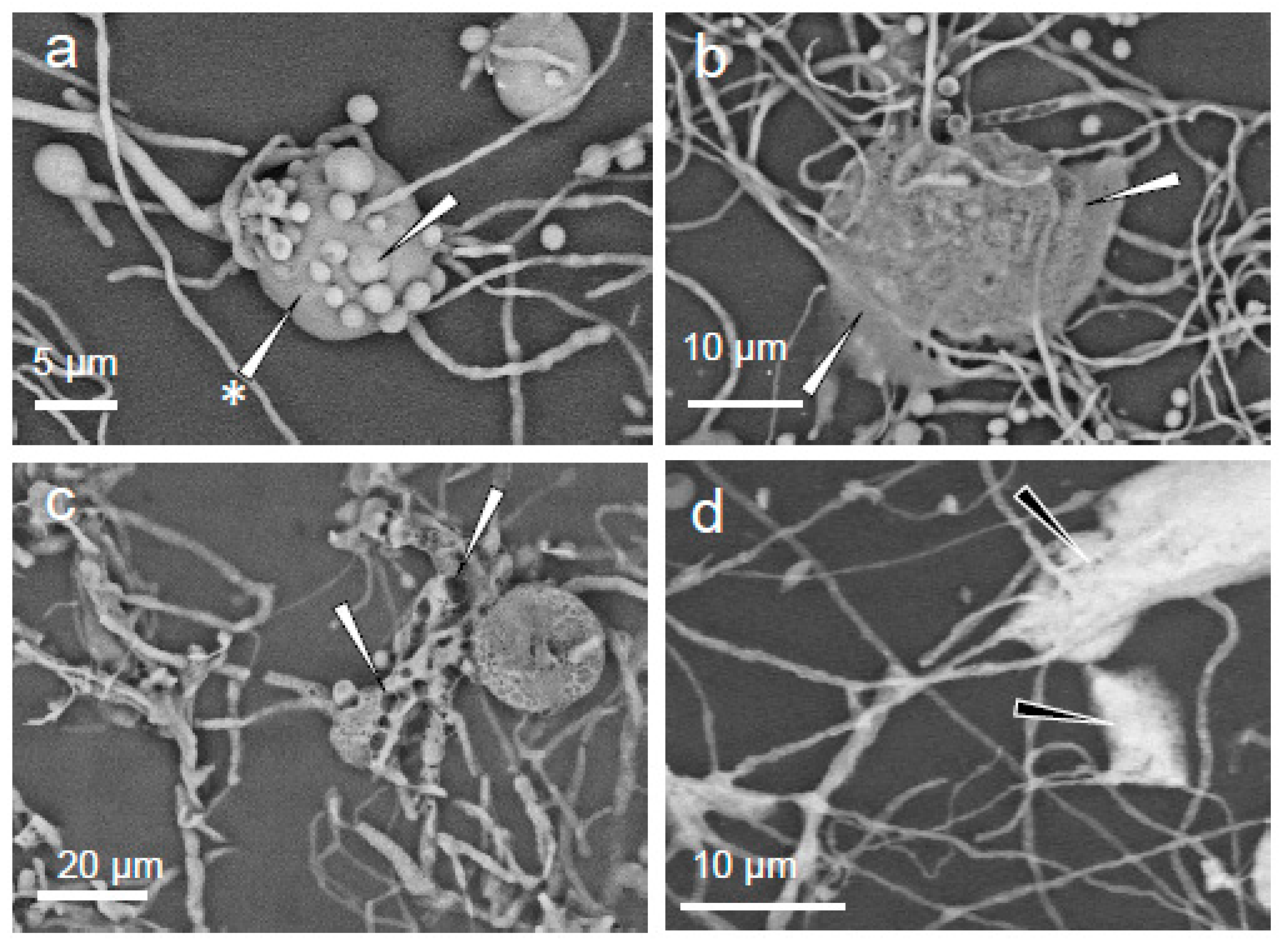
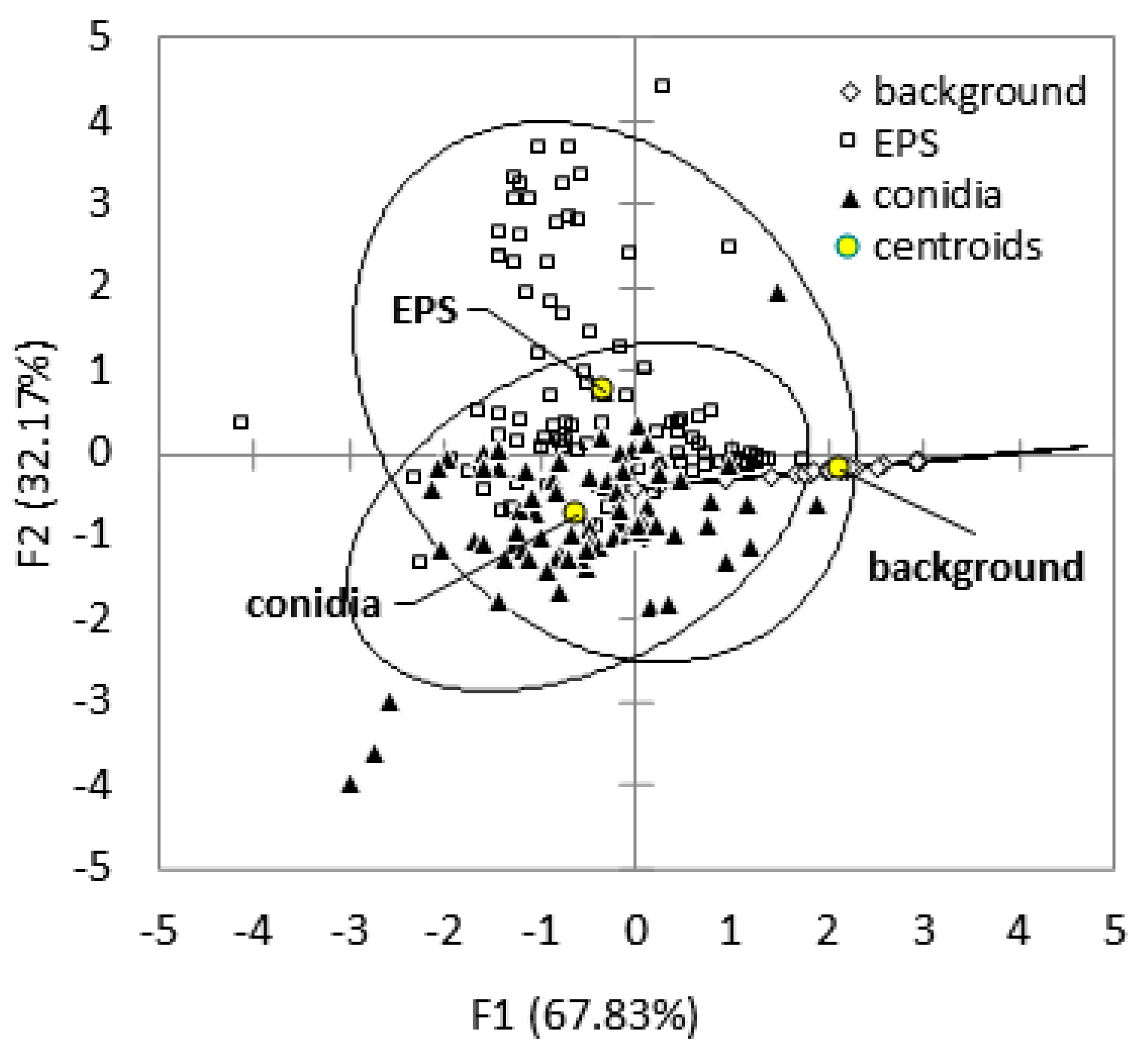
2. Results
3. Discussion
4. Materials and Methods
4.1. Sampling Procedure
4.2. Scanning Electron Microscopy
4.3. X-ray Powder Diffraction
4.4. Energy-Dispersive X-ray Spectroscopy
4.5. Statistical Analysis of Energy-Dispersive X-ray Spectroscopy Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, J.; Zhou, L.; Foltz, G.R.; Qu, X.; Ying, J.; Tokinaga, H.; Mechoso, C.R.; Li, J.; Gu, X. Hydrological cycle changes under global warming and their effects on multiscale climate variability. Ann. N. Y. Acad. Sci. 2020, 1472, 21–48. [Google Scholar] [CrossRef] [PubMed]
- Seager, R.; Henderson, N.; Vecchi, G. Thermodynamic and dynamic mechanisms for large-scale changes in the hydrological cycle in response to global warming. J. Clim. 2010, 23, 4651–4668. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, C.M.; Papavizas, G.C.; Benjamin, C.R. A new halophilic species of Eurotium. Mycologia 1959, 51, 636–640. [Google Scholar] [CrossRef]
- Hocking, A.D.; Pitt, J.I. Two new species of xerophilic fungi and a further record of Eurotium halophilicum. Mycologia 1988, 80, 82–88. [Google Scholar] [CrossRef]
- Abdel-Hafez, S.I.I.; Moubasher, A.H.; Barakat, A. Keratinophilic fungi and other moulds associated with air-dust particles from Egypt. Folia Microbiol. 1990, 35, 311–325. [Google Scholar] [CrossRef]
- Samson, R.A.; van der Lustgraaf, B. Aspergillus penicilloides and Eurotium halophilicum in association with house-dust mites. Mycopathologia 1978, 64, 13–16. [Google Scholar] [CrossRef]
- Huerta, T.; Sanchis, V.; Hernandez, J.; Hernandez, E. Mycoflora of dry-salted Spanish ham. Microbiol. Alim. Nutr. 1987, 5, 247–252. [Google Scholar]
- Xu, A.; Wang, Y.; Wen, J.; Liu, P.; Liu, Z.; Li, Z. Fungal community associated with fermentation and storage of Fuzhuan brick-tea. Int. J. Food Microbiol. 2011, 146, 14–22. [Google Scholar] [CrossRef]
- Pinzari, F.; Montanari, M. Mould growth on library materials stored in Compactus-type shelving units. In Sick Building Syndrome: Public Buildings and Workplaces; Abdul-Wahab Al-Sulaiman, S.A., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 193–206. [Google Scholar]
- Montanari, M.; Melloni, V.; Pinzari, F.; Innocenti, G. Fungal biodeterioration of historical library materials stored in Compactus movable shelves. Int. Biodeter. Biodegr. 2012, 75, 83–88. [Google Scholar] [CrossRef]
- Micheluz, A.; Manente, S.; Tigini, V.; Prigione, V.; Pinzari, F.; Ravagnan, G.; Varese, G.C. The extreme environment of a library: Xerophilic fungi inhabiting indoor niches. Int. Biodeter. Biodegr. 2015, 99, 1–7. [Google Scholar] [CrossRef]
- Fiorillo, F.; Fiorentino, S.; Montanari, M.; Roversi Monaco, C.; Del Bianco, A.; Vandini, M. Learning from the past, intervening in the present: The role of conservation science in the challenging restoration of the wall painting Marriage at Cana by Luca Longhi (Ravenna, Italy). Herit. Sci. 2020, 8, 10. [Google Scholar] [CrossRef]
- Kavkler, K.; Humar, M.; Kržišnik, D.; Turk, M.; Tavzes, Č.; Gostinčar, C.; Džeroski, S.; Popov, S.; Penko, A.; Gunde–Cimerman, N.; et al. A multidisciplinary study of biodeteriorated Celje Ceiling, a tempera painting on canvas. Int. Biodeter. Biodegr. 2022, 170, 105389. [Google Scholar] [CrossRef]
- Bastholm, C.J.; Madsen, A.M.; Andersen, B.; Frisvad, J.C.; Richter, J. The mysterious mould outbreak—A comprehensive fungal colonisation in a climate-controlled museum repository challenges the environmental guidelines for heritage collections. J. Cult. Herit. 2022, 55, 78–87. [Google Scholar] [CrossRef]
- Andrews, S.; Pitt, J.I. Further studies on the water relations of xerophilic fungi, including some halophiles. J. Gen. Microbiol. 1987, 133, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, A.; Hamill, P.G.; Medina, Á.; Kminek, G.; Rummel, J.D.; Dijksterhuis, J.; Timson, D.J.; Magan, N.; Leong, S.L.; Hallsworth, J.E. Glycerol enhances fungal germination at the water-activity limit for life. Environ. Microbiol. 2017, 19, 947–967. [Google Scholar] [CrossRef] [Green Version]
- Piñar, G.; Tafer, H.; Sterflinger, K.; Pinzari, F. Amid the possible causes of a very famous foxing: Molecular and microscopic insight into Leonardo da Vinci’s self-portrait. Environ. Microbiol. Rep. 2015, 7, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Y.; Zhang, F.; Hu, C.; Liu, G.; Pan, J. Microbial community analyses of the deteriorated storeroom objects in the Tianjin Museum using culture-independent and culture-dependent approaches. Front. Microbiol. 2018, 9, 802. [Google Scholar] [CrossRef]
- Chin, J.P.; Megaw, J.; Magill, C.L.; Nowotarski, K.; Williams, J.P.; Bhaganna, P.; Linton, M.; Patterson, M.F.; Underwood, G.J.; Mswaka, A.Y.; et al. Solutes determine the temperature windows for microbial survival and growth. Proc. Natl. Acad. Sci. USA 2010, 107, 7835–7840. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, A.; Cray, J.A.; Williams, J.P.; Santos, R.; Sahay, R.; Neuenkirchen, N.; McClure, C.D.; Grant, I.R.; Houghton, J.D.; Quinn, J.P.; et al. Is there a common water-activity limit for the three domains of life? ISME J. 2015, 9, 1333–1351. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, A.; Burkhardt, J.; Cockell, C.S.; Cray, J.A.; Dijksterhuis, J.; Fox-Powell, M.; Kee, T.P.; Kminek, G.; McGenity, T.J.; Timmis, K.N.; et al. Multiplication of microbes below 0.690 water activity: Implications for terrestrial and extraterrestrial life. Environ. Microbiol. 2015, 17, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Chiang, Y.W.; Santos, R.M. X-ray diffraction techniques for mineral characterization: A review for engineers of the fundamentals, applications, and research directions. Minerals 2022, 12, 205. [Google Scholar] [CrossRef]
- Audra, P.; Nobécourt, J.-C. Rare sulfates (mirabilite, eugsterite) in the dry microclimate of Chamois Cave (Alpes-de-Haute-Provence, France). In Proceedings of the 16th International Congress of Speleology, Brno, Czech Republic, 21–28 July 2013; Volume 3. [Google Scholar]
- Steiger, M. Salts in porous materials: Thermodynamics of phase transitions, modeling and preventive conservation. RBM 2005, 11, 419–432. [Google Scholar] [CrossRef]
- Mills, S.J.; Hatert, F.; Nickel, E.H.; Ferraris, G. The standardisation of mineral group hierarchies: Application to recent nomenclature proposals. Eur. J. Miner. 2009, 21, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Fox-Powell, M.G.; Hallsworth, J.E.; Cousins, C.R.; Cockell, C.S. Ionic strength is a barrier to the habitability of Mars. Astrobiology 2016, 16, 427–442. [Google Scholar] [CrossRef]
- Hallsworth, J.E. Salt deliquescence can support extraterrestrial life. Nat. Astron. 2020, 4, 739–740. [Google Scholar] [CrossRef]
- Rivera-Valentín, E.G.; Chevrier, V.F.; Soto, A.; Martínez, G. Distribution and habitability of (meta)stable brines on present-day Mars. Nat. Astron. 2020, 4, 756–761. [Google Scholar] [CrossRef]
- Davila, A.F.; Duport, L.G.; Melchiorri, R.; Jänchen, J.; Valea, S.; de Los Rios, A.; Fairén, A.G.; Möhlmann, D.; McKay, C.P.; Ascaso, C.; et al. Hygroscopic salts and the potential for life on Mars. Astrobiology 2010, 10, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Davila, A.F.; Hawes, I.; Ascaso, C.; Wierzchos, J. Photosynthesis in the Atacama Desert. Environ. Microbiol. Rep. 2013, 5, 583–587. [Google Scholar] [CrossRef]
- Wierzchos, J.; Davila, A.F.; Sánchez-Almazo, I.M.; Hajnos, M.; Swieboda, R.; Ascaso, C. Novel water source for endolithic life in the hyperarid core of the Atacama Desert. Biogeosciences 2012, 9, 2275–2286. [Google Scholar] [CrossRef] [Green Version]
- Micheluz, A.; Manente, S.; Prigione, V.; Tigini, V.; Varese, G.C.; Ravagnan, G. The effects of books disinfection to the airborne microbiological community in a library environment. Aerobiologia 2018, 34, 29–44. [Google Scholar] [CrossRef]
- Steiger, M.; Asmussen, S. Crystallization of sodium sulfate phases in porous materials: The phase diagram of Na2SO4-H2O and the generation of stress. Geochim. Cosmochim. Acta 2008, 72, 4291–4306. [Google Scholar] [CrossRef]
- Peng, C.; Chen, L.; Tang, M. A database for deliquescence and efflorescence relative humidities of compounds with atmospheric relevance. Fundam. Res. 2022, 2, 578–587. [Google Scholar] [CrossRef]
- Gough, R.V.; Chevrier, V.F.; Tolbert, M.A. Formation of aqueous solutions on Mars via deliquescence of chloride-perchlorate binary mixtures. Earth Planet. Sci. Lett. 2014, 393, 73–82. [Google Scholar] [CrossRef]
- Chevrier, V.F.; Fitting, A.F.; Rivera-Valentín, E.G. Limited stability of multicomponent brines on the surface of Mars. Planet. Sci. J. 2022, 3, 125. [Google Scholar] [CrossRef]
- Alava, M.; Niskanen, K. The physics of paper. Rep. Prog. Phys. 2006, 69, 669–723. [Google Scholar] [CrossRef]
- Cray, J.A.; Russell, J.T.; Timson, D.J.; Singhal, R.S.; Hallsworth, J.E. A universal measure of chaotropicity and kosmotropicity. Environ. Microbiol. 2013, 15, 287–296. [Google Scholar] [CrossRef]
- Pitt, J.I. Xerophilic fungi and the spoilage of foods of plant origin. In Water Relations of Foods; Duckworth, R.B., Ed.; Academic Press: London, UK, 1975; pp. 273–307. [Google Scholar]
- De Hoog, S.; Zalar, P.; Van Den Ende, B.G.; Gunde-Cimerman, N. Relation of halotolerance to human-pathogenicity in the fungal tree of life: An overview of ecology and evolution under stress. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya. Cellular Origin, Life in Extreme Habitats and Astrobiology; Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherland, 2005; Volume 9. [Google Scholar] [CrossRef]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [Green Version]
- Nazareth, S.; Gonsalves, V. Aspergillus penicillioide–A true halophile existing in hypersaline and polyhaline econiches. Ann. Microbiol. 2014, 64, 397–402. [Google Scholar] [CrossRef]
- Stevenson, A.; Hamill, P.G.; O’Kane, C.J.; Kminek, G.; Rummel, J.D.; Voytek, M.A.; Dijksterhuis, J.; Hallsworth, J.E. Aspergillus penicillioides differentiation and cell division at 0.585 water activity. Environ. Microbiol. 2017, 19, 687–697. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 2nd ed.; Blackie Academic and Professional: London, UK, 1997. [Google Scholar]
- Polo, A.; Cappitelli, F.; Villa, F.; Pinzari, F. Biological invasion in the indoor environment: The spread of Eurotium halophilicum on library materials. Int. Biodeter. Biodegr. 2017, 118, 34–44. [Google Scholar] [CrossRef]
- Stevenson, A.; Hamill, P.G.; Dijksterhuis, J.; Hallsworth, J.E. Water-, pH- and temperature relations of germination for the extreme xerophiles Xeromyces bisporus (FRR 0025), Aspergillus penicillioides (JH06THJ) and Eurotium halophilicum (FRR 2471). Microb. Biotechnol. 2017, 10, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Magan, N.; Lacey, J. Effect of temperature and pH on water relations of field and storage fungi. Trans. Br. Mycol. Soc. 1984, 82, 71–81. [Google Scholar] [CrossRef]
- Kashangura, C.; Hallsworth, J.E.; Mswaka, A.Y. Phenotypic diversity amongst strains of Pleurotus sajor-caju: Implications for cultivation in arid environments. Mycol. Res. 2006, 110, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.P.; Hallsworth, J.E. Limits of life in hostile environments: No barriers to biosphere function? Environ. Microbiol. 2009, 11, 3292–3308. [Google Scholar] [CrossRef] [Green Version]
- Flipphi, M.; Sun, J.; Robellet, X.; Karaffa, L.; Fekete, E.; Zeng, A.P.; Kubicek, C.P. Biodiversity and evolution of primary carbon metabolism in Aspergillus nidulans and other Aspergillus spp. Fungal Genet. Biol. 2009, 46, S19–S44. [Google Scholar] [CrossRef]
- Cray, J.A.; Bell, A.N.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The biology of habitat dominance; can microbes behave as weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef] [Green Version]
- Dijksterhuis, J.; van Egmond, W.; Yarwood, A. From colony to rodlet: “A six meter long portrait of the xerophilic fungus Aspergillus restrictus decorates the hall of the Westerdijk institute”. Fungal Biol. 2020, 124, 509–515. [Google Scholar] [CrossRef]
- Sklenář, F.; Jurjević, Ž.; Zalar, P.; Frisvad, J.C.; Visagie, C.M.; Kolařík, M.; Houbraken, J.; Chen, A.J.; Yilmaz, N.; Seifert, K.A.; et al. Phylogeny of xerophilic aspergilli (subgenus Aspergillus) and taxonomic revision of section Restricti. Stud. Mycol. 2017, 88, 161–236. [Google Scholar] [CrossRef] [Green Version]
- Bartnicki-Garcia, S. Chitosomes: Past, present and future. FEMS Yeast Res. 2006, 6, 957–965. [Google Scholar] [CrossRef]
- Brown, A.D. Microbial water stress. Bacteriol Rev. 1976, 40, 803–846. [Google Scholar] [CrossRef] [PubMed]
- Treseder, K.K.; Lennon, J.T. Fungal traits that drive ecosystem dynamics on land. Microbiol. Mol. Biol. Rev. 2015, 79, 243–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajc, J.; Zalar, P.; Plemenitaš, A.; Gunde-Cimerman, N. The mycobiota of the salterns. In Biology of Marine Fungi; Raghukumar, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 133–158. [Google Scholar]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Fungal adaptation to extremely high salt concentrations. Adv. Appl. Microbiol. 2011, 77, 71–96. [Google Scholar] [CrossRef] [PubMed]
- De Lima Alves, F.; Stevenson, A.; Baxter, E.; Gillion, J.L.; Hejazi, F.; Hayes, S.; Morrison, I.E.; Prior, B.A.; McGenity, T.J.; Rangel, D.E.; et al. Concomitant osmotic and chaotropicity-induced stresses in Aspergillus wentii: Compatible solutes determine the biotic window. Curr. Genet. 2015, 61, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Kralj Kunčič, M.; Kogej, T.; Drobne, D.; Gunde-Cimerman, N. Morphological response of the halophilic fungal genus Wallemia to high salinity. Appl. Environ. Microbiol. 2010, 76, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Varliero, G.; Hallsworth, J.E.; Dallas, T.D.; Hopkins, D.; Frey, B.; Kong, W.; Lebre, P.; Makhalanyane, T.P.; Cowan, D.A. Microbial anhydrobiosis. Environ. Microbiol. 2021, 23, 6377–6390. [Google Scholar] [CrossRef]
- Breierová, E.; Stratilová, E.; Šajbidor, J. Salinity affects fatty acid and extracellular glycoprotein composition of Dipodascus australiensis. Food Technol. Biotechnol. 1997, 35, 237–241. [Google Scholar]
- Breierová, E.; Kačcuráková, M.; Stratilová, E. The effect of high concentration of NaCl in the cultivation medium on changes in the composition of extracellular yeast glycoproteins. System. Appl. Microbiol. 1997, 20, 348–355. [Google Scholar] [CrossRef]
- Hallsworth, J.E. Wooden owl that redefines Earth’s biosphere may yet catapult a fungus into space. Environ. Microbiol. 2019, 21, 2202–2211. [Google Scholar] [CrossRef]
- Grau-Bové, J.; Strlič, M. Fine particulate matter in indoor cultural heritage: A literature review. Herit. Sci. 2013, 1, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Drougka, F.; Liakakou, E.; Sakka, A.; Mitsos, D.; Zacharias, N.; Mihalopoulos, N.; Gerasopoulos, E. Indoor Air Quality Assessment at the Library of the National Observatory of Athens, Greece. Aerosol Air Qual. Res. 2020, 20, 889–903. [Google Scholar] [CrossRef]
- Bücking, H.; Heyser, W. Elemental composition and function of polyphosphates in ectomycorrhizal fungi—An X-ray microanalytical study. Mycol. Res. 1999, 103, 31–39. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hijikata, N.; Yokoyama, K.; Ohtomo, R.; Handa, Y.; Kawaguchi, M.; Saito, K.; Ezawa, T. Polyphosphate accumulation is driven by transcriptome alterations that lead to near-synchronous and near-equivalent uptake of inorganic cations in an arbuscular mycorrhizal fungus. New Phytol. 2014, 204, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Bradfield, G.; Somerfield, P.; Meyn, T.; Holby, M.; Babcock, D.; Bradley, D.; Segel, I.H. Regulation of sulfate transport in filamentous fungi. Plant Physiol. 1970, 46, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D. Papermaking, the History and Technique of an Ancient Craft; Dover Publication: New York, NY, USA, 1974. [Google Scholar]
- Reed, R. The Nature and Making of Parchment; The Elmete Press: Leeds, UK, 1975. [Google Scholar]
- Pinzari, F. Microbial processes involved in the deterioration of paper and parchment. In Biodeterioration and Preservation in Art, Archaeology and Architecture; Mitchell, R., Clifford, J., Eds.; Publications Ltd.: London, UK, 2018; pp. 33–56. [Google Scholar]
- Bicchieri, M.; Biocca, P.; Colaizzi, P.; Pinzari, F. Microscopic observations of paper and parchment: The archaeology of small objects. Herit. Sci. 2019, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Kumagai, H.; Oohashi, K.; Sakamoto, K.; Inubushi, K.; Enomoto, S. Transport of trace elements through the hyphae of an arbuscular mycorrhizal fungus into marigold determined by the multitracer technique. Soil Sci. Plant Nutr. 2001, 47, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.S.; Jellison, J. Extraction and translocation of calcium from gypsum during wood biodegradation by oxalate-producing fungi. Int. Biodeterior. Biodegr. 2007, 60, 8–15. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Olsson, S. Fungal translocation—Creating and responding to environmental heterogeneity. Mycologist 2004, 18, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Pinzari, F.; Cuadros, J.; Migliore, M.; Napoli, R.; Najorka, J. Manganese translocation and concentration on Quercus cerris decomposing leaf and wood litter by an ascomycetous fungus: An active process with ecosystem consequences? FEMS Microbiol. Ecol. 2018, 94, fiy111. [Google Scholar] [CrossRef] [Green Version]
- Boswell, G.P.; Jacobs, H.; Davidson, F.A.; Gadd, G.M.; Ritz, K. Growth and function of fungal mycelia in heterogeneous environments. Bull. Math. Biol. 2003, 65, 447–477. [Google Scholar] [CrossRef]
- Arai, H. Foxing caused by fungi: Twenty-five years of study. Int. Biodeter. Biodegr. 2000, 46, 181–188. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Kaminskyj, S.G.; Dahms, T.E. High spatial resolution surface imaging and analysis of fungal cells using SEM and AFM. Micron 2008, 39, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.I.; Newbury, D.E.; Michael, J.R.; Ritchie, N.W.M.; Scott, J.H.J.; Joy, D.C. Scanning Electron Microscopy and X-ray Microanalysis, 4th ed.; Springer: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Massart, D.L.; Vandeginste, B.G.M.; Buydens, L.M.C.; de Jong, S.; Lewi, P.J.; Smeyers Verbeke, J. Handbook of Chemometrics and Qualimetrics. Part B; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Reimann, C.; Filzmoser, P.; Garrett, R.G.; Dutter, R. Statistical Data Analysis Explained. Applied Environmental Statistics with R; Wiley: Chichester, UK, 2008. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Mancinelli, R.L.; Conley, C.A.; Dallas, T.D.; Rinaldi, T.; Davila, A.F.; Benison, K.C.; Rapoport, A.; Cavalazzi, B.; Selbmann, L.; et al. Astrobiology of life on Earth. Environ. Microbiol. 2021, 23, 3335–3344. [Google Scholar] [CrossRef] [PubMed]
- Hallsworth, J.E. Water is a preservative of microbes. Microbial. Biotechnol. 2022, 15, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Kauserud, H.; Heegaard, E.; Semenov, M.A.; Boddy, L.; Halvorsen, R.; Stige, L.C.; Sparks, T.H.; Gange, A.C.; Stenseth, N.C. Climate change and spring-fruiting fungi. Proc. Biol. Sci. 2010, 277, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Větrovský, T.; Kohout, P.; Kopecký, M.; Machac, A.; Man, M.; Bahnmann, B.D.; Brabcová, V.; Choi, J.; Meszárošová, L.; Human, Z.R.; et al. A meta-analysis of global fungal distribution reveals climate-driven patterns. Nat. Commun. 2019, 10, 5142. [Google Scholar] [CrossRef] [Green Version]
- Baxter, B.; Butler, J. Great Salt Lake Biology; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Kumar, V.; Sarma, V.V.; Thambugala, K.M.; Huang, J.-J.; Li, X.-Y.; Hao, G.-F. Ecology and evolution of marine fungi with their adaptation to climate change. Front. Microbiol. 2021, 12, 2527. [Google Scholar] [CrossRef]
- Timmis, K.; Hallsworth, J.E. The darkest microbiome: A post-human biosphere. Microbial. Biotechnol. 2022, 15, 176–185. [Google Scholar] [CrossRef]
- Kosel, J.; Kvačič, M.; Legan, L.; Retko, K.; Ropret, P. Evaluating the xerophilic potential of moulds on selected egg tempera paints on glass and wooden supports using fluorescent microscopy. J. Cult. Herit. 2021, 52, 44–54. [Google Scholar] [CrossRef]
- Lintang, W.; Susetyo-Salim, T.; Oetari, A.; Sjamsuridzal, W. Isolation and characterization of fungi from deteriorated old manuscripts from Banyumas, collection of Library of Universitas Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 948, 012031. [Google Scholar] [CrossRef]
- Hagiuda, R.; Oda, H.; Kawakami, Y.; Hirose, D. Species diversity based on revised systematics of xerophilic Aspergillus section Restricti isolated from storage rooms and houses in Japan. Biocontrol Sci. 2022, 27, 65–80. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories of Observations | C | O | Na | P | S | Cl | K | Ca |
|---|---|---|---|---|---|---|---|---|
| Conidia | 76.250 b | 21.954 b | 0.175 b | 0.233 a | 0.819 b | 0.173 a | 0.361 a | 0.036 b |
| Extracellular polymeric substances | 65.861 c | 25.654 a | 3.617 a | 0.013 b | 3.054 a | 0.145 a | 0.133 b | 1.524 a |
| Background | 92.379 a | 7.591 c | 0.000 c | 0.000 c | 0.031 c | 0.000 b | 0.000 c | 0.000 c |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0001 | <0.0001 | <0.0001 |
| Significant | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Parameters of the Principal Components | F1 | F2 | F3 |
|---|---|---|---|
| Eigenvalue | 3.615 | 2.072 | 1.048 |
| Variability (%) | 45.186 | 25.898 | 13.095 |
| Cumulative variability (%) | 45.186 | 71.084 | 84.179 |
| From\To | Conidia | Extracellular Polymeric Substances | Background | Total | Correct (%) |
|---|---|---|---|---|---|
| Conidia | 68 | 11 | 3 | 82 | 82.93 |
| Extracellular polymeric substances | 16 | 67 | 10 | 93 | 72.04 |
| Background | 5 | 1 | 32 | 38 | 84.21 |
| Total | 89 | 79 | 45 | 213 | 78.40 |
| Variable | C | O | Na | P | S | Cl | K | Ca |
|---|---|---|---|---|---|---|---|---|
| C | −0.92 | −0.77 | −0.20 | −0.71 | 0.04 | −0.18 | −0.63 | |
| O | <0.0001 | 0.56 | 0.13 | 0.45 | 0.03 | 0.12 | 0.44 | |
| Na | <0.0001 | <0.0001 | −0.12 | 0.82 | −0.12 | −0.13 | 0.61 | |
| P | 0.003 | 0.05 | 0.08 | −0.07 | −0.07 | 0.97 | −0.06 | |
| S | <0.0001 | <0.0001 | <0.0001 | 0.34 | −0.08 | −0.09 | 0.54 | |
| Cl | 0.59 | 0.66 | 0.08 | 0.31 | 0.22 | −0.06 | −0.15 | |
| K | 0.01 | 0.07 | 0.06 | <0.0001 | 0.18 | 0.36 | −0.07 | |
| Ca | <0.0001 | <0.0001 | <0.0001 | 0.41 | <0.0001 | 0.02 | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micheluz, A.; Pinzari, F.; Rivera-Valentín, E.G.; Manente, S.; Hallsworth, J.E. Biophysical Manipulation of the Extracellular Environment by Eurotium halophilicum. Pathogens 2022, 11, 1462. https://doi.org/10.3390/pathogens11121462
Micheluz A, Pinzari F, Rivera-Valentín EG, Manente S, Hallsworth JE. Biophysical Manipulation of the Extracellular Environment by Eurotium halophilicum. Pathogens. 2022; 11(12):1462. https://doi.org/10.3390/pathogens11121462
Chicago/Turabian StyleMicheluz, Anna, Flavia Pinzari, Edgard G. Rivera-Valentín, Sabrina Manente, and John E. Hallsworth. 2022. "Biophysical Manipulation of the Extracellular Environment by Eurotium halophilicum" Pathogens 11, no. 12: 1462. https://doi.org/10.3390/pathogens11121462
APA StyleMicheluz, A., Pinzari, F., Rivera-Valentín, E. G., Manente, S., & Hallsworth, J. E. (2022). Biophysical Manipulation of the Extracellular Environment by Eurotium halophilicum. Pathogens, 11(12), 1462. https://doi.org/10.3390/pathogens11121462