
Canine Staphylococcus argenteus: Case Report from The Netherlands

 ,
,  ,
,
Abstract
:1. Introduction
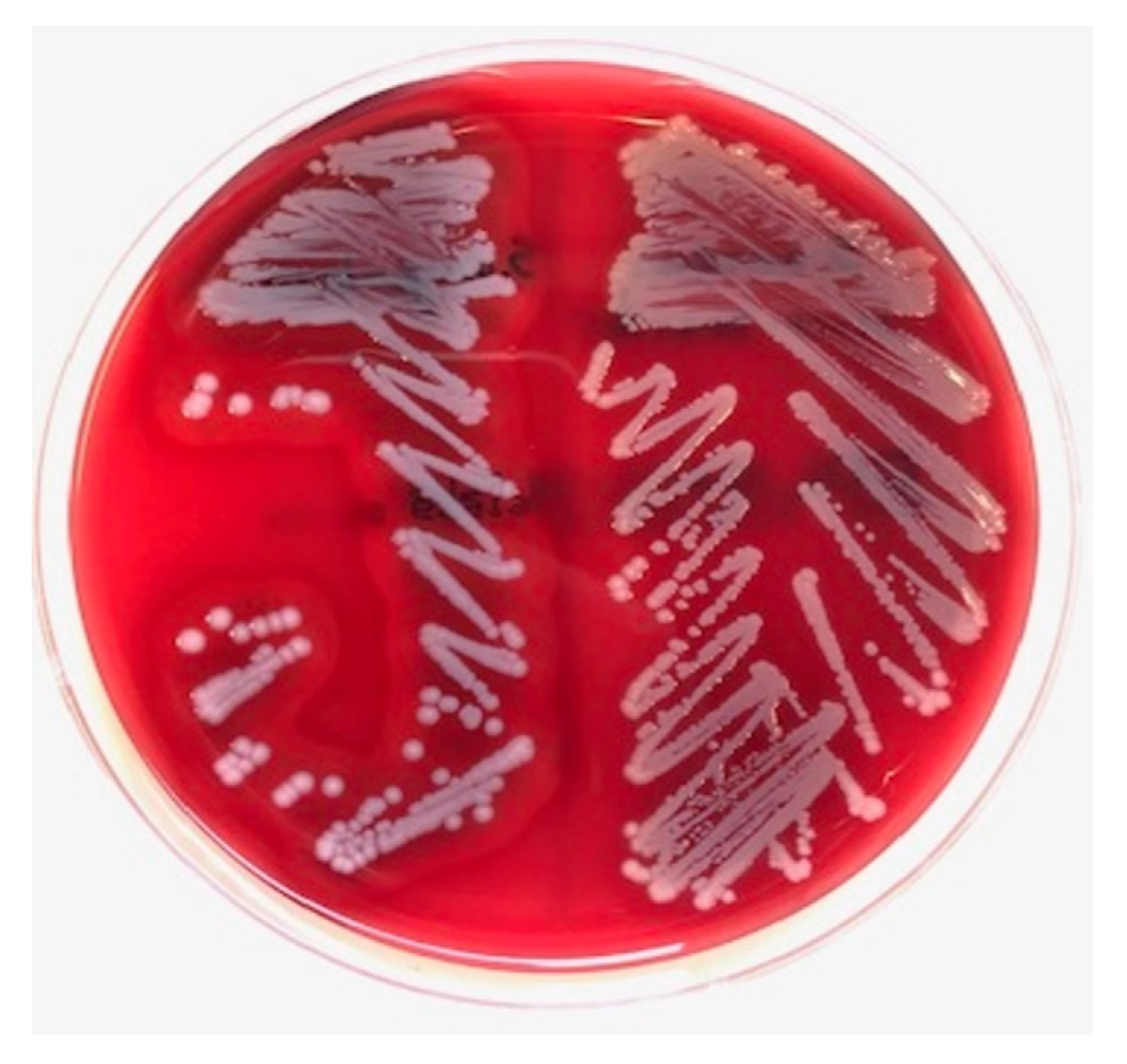
2. Case
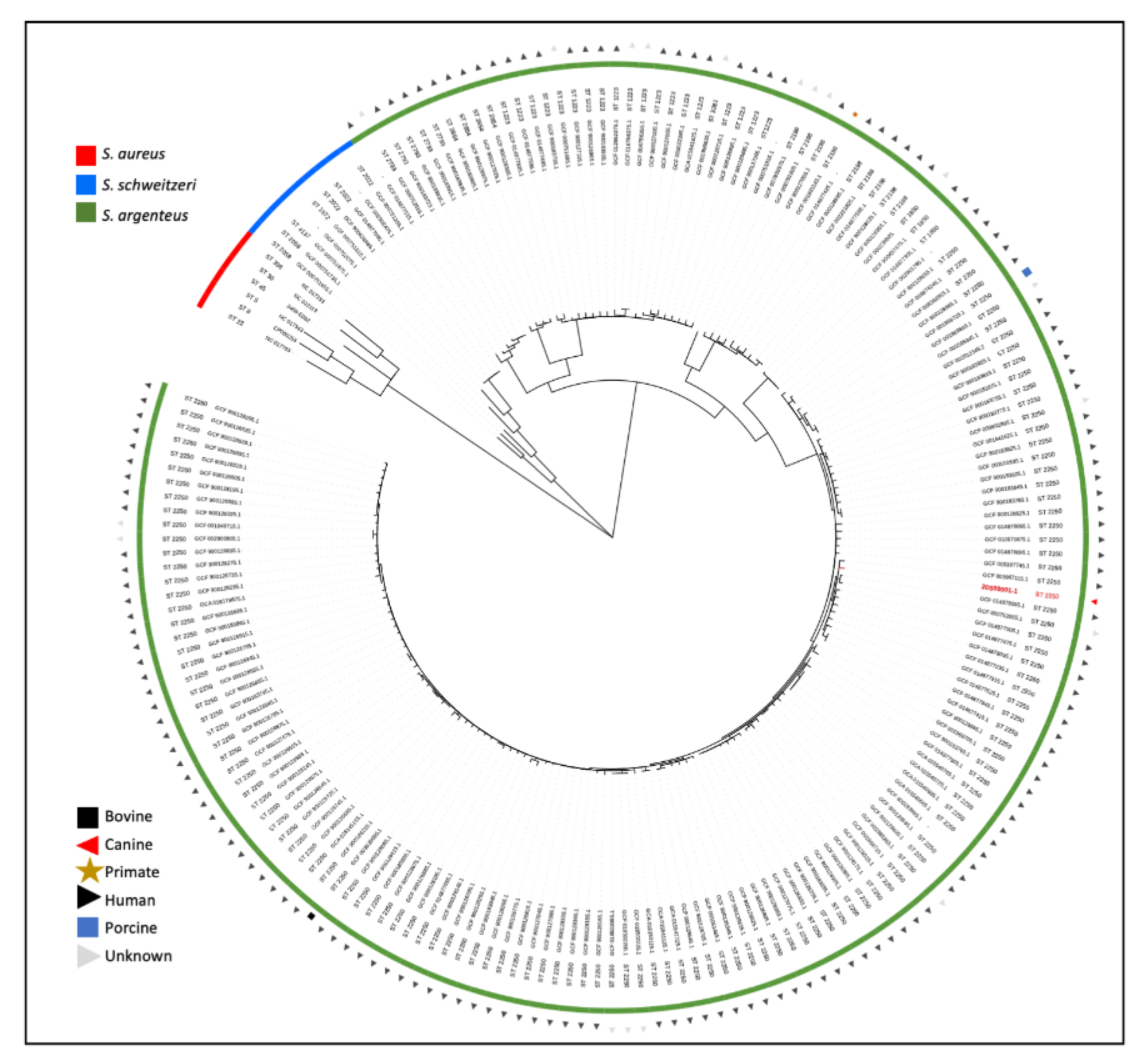
3. Materials and Methods
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tong, S.Y.C.; Schaumburg, F.; Ellington, M.J.; Corander, J.; Pichon, B.; Leendertz, F.; Bentley, S.D.; Parkhill, J.; Holt, D.; Peters, G.; et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: The non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 1, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Schaumburg, F.; Kearns, A.; Larsen, A.; Lindsay, J.; Skov, R.; Westh, H. Implications of identifying the recently defined members of the Staphylococcus aureus complex S. argenteus and S. schweitzeri: A position paper of members of the ESCMID Study Group for Staphylococci and Staphylococcal Diseases (ESGS). Clin. Microbiol. Infect. 2019, 25, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Bank, L.E.A.; Bosch, T.; Schouls, L.M.; Weersink, A.J.L.; Witteveen, S.; Wolffs, P.F.G.; Nijhuis, R.H.T. Methicillin-resistant Staphylococcus argenteus in the Netherlands: Not a new arrival. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1583–1585. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, A.; Froböse, N.J.; Mellmann, A.; Alabi, A.S.; Schaumburg, F.; Niemann, S. An in vitro study on Staphylococcus schweitzeri virulence. Sci. Rep. 2021, 11, 1157. [Google Scholar] [CrossRef]
- Zhang, D.-F.; Zhi, X.-Y.; Zhang, J.; Paoli, G.C.; Cui, Y.; Shi, C.; Shi, X. Preliminary comparative genomics revealed pathogenic potential and international spread of Staphylococcus argenteus. BMC Genom. 2017, 18, 808. [Google Scholar] [CrossRef] [Green Version]
- Thaipadungpanit, J.; Amornchai, P.; Nickerson, E.K.; Wongsuvan, G.; Wuthiekanun, V.; Limmathurotsakul, D.; Peacock, S.J. Clinical and Molecular Epidemiology of Staphylococcus argenteus Infections in Thailand. J. Clin. Microbiol. 2015, 53, 1005. [Google Scholar] [CrossRef] [Green Version]
- Schuster, D.; Rickmeyer, J.; Gajdiss, M.; Thye, T.; Lorenzen, S.; Reif, M.; Josten, M.; Szekat, C.; Melo, L.D.; Schmithausen, R.M.; et al. Differentiation of Staphylococcus argenteus (formerly: Staphylococcus aureus clonal complex 75) by mass spectrometry from S. aureus using the first strain isolated from a wild African great ape. Int. J. Med Microbiol. 2017, 307, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Pumipuntu, N.; Tunyong, W.; Chantratita, N.; Diraphat, P.; Pumirat, P.; Sookrung, N.; Chaicumpa, W.; Indrawattana, N. Staphylococcus spp. associated with subclinical bovine mastitis in central and northeast provinces of Thailand. PeerJ 2019, 7, e6587. [Google Scholar] [CrossRef] [Green Version]
- Indrawattana, N.; Pumipuntu, N.; Suriyakhun, N.; Jangsangthong, A.; Kulpeanprasit, S.; Chantratita, N.; Sookrung, N.; Chaicumpa, W.; Buranasinsup, S. Staphylococcus argenteusfrom rabbits in Thailand. Microbiologyopen 2018, 8, e00665. [Google Scholar] [CrossRef] [Green Version]
- Kaden, R.; Engstrand, L.; Rautelin, H.; Johansson, C. Which methods are appropriate for the detection of Staphylococcus argenteus and is it worthwhile to distinguish S. argenteus from S. aureus? Infect. Drug Resist. 2018, 11, 2335–2344. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Geno-type-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2014, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0. 2021. Available online: http://www.eucast.org (accessed on 25 January 2022).
- CLSI. Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; CLSI Supplement VET01S; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2020. [Google Scholar]
- Chen, S.-Y.; Lee, H.; Teng, S.-H.; Wang, X.-M.; Lee, T.-F.; Huang, Y.-C.; Liao, C.-H.; Teng, L.-J.; Hsueh, P.-R. Accurate differentiation of novel Staphylococcus argenteus from Staphylococcus aureus using MALDI-TOF MS. Future Microbiol. 2018, 13, 997–1006. [Google Scholar] [CrossRef]
- Jones, D.; Meijer, E.F.J.; Blatter, C.; Liao, S.; Pereira, E.R.; Bouta, E.M.; Jung, K.; Chin, S.M.; Huang, P.; Munn, L.L.; et al. Methicillin-resistant Staphylococcus aureus causes sustained collecting lymphatic vessel dysfunction. Sci. Transl. Med. 2018, 10, eaam7964. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Hooper, D.C. Effect of Staphylococcus aureus Tet38 native efflux pump on in vivo response to tetracycline in a murine subcutaneous abscess model. J. Antimicrob. Chemother. 2017, 73, 720–723. [Google Scholar] [CrossRef]
- Goswami, C.; Fox, S.; Holden, M.; Leanord, A.; Evans, T.J. Genomic Analysis of Global Staphylococcus argenteus Strains Reveals Distinct Lineages With Differing Virulence and Antibiotic Resistance Gene Content. Front. Microbiol. 2021, 12, 795173. [Google Scholar] [CrossRef] [PubMed]
- DiCicco, M.; Weese, S.; Neethirajan, S.; Rousseau, J.; Singh, A. Fosfomycin susceptibility of canine methicillin-resistant Staphylococcus pseudintermedius isolates. Res. Vet. Sci. 2014, 96, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Helmann, J.D. Bacillithiol, a New Player in Bacterial Redox Homeostasis. Antioxid. Redox Signal. 2011, 15, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Hendlin, D.; Stapley, E.O.; Jackson, M.; Wallick, H.; Miller, A.K.; Wolf, F.J.; Miller, T.W.; Chaiet, L.; Kahan, F.M.; Foltz, E.L.; et al. Phosphonomycin, a New Antibiotic Produced by Strains of Streptomyces. Science 1969, 166, 122–123. [Google Scholar] [CrossRef]
- Woodyer, R.D.; Shao, Z.; Thomas, P.M.; Kelleher, N.L.; Blodgett, J.A.; Metcalf, W.W.; van der Donk, W.A.; Zhao, H. Heterologous Production of Fosfomycin and Identification of the Minimal Biosynthetic Gene Cluster. Chem. Biol. 2006, 13, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-C.; Chen, P.-Y.; Wang, J.-T.; Chang, S.-C. Prevalence of fosfomycin resistance and gene mutations in clinical isolates of methicillin-resistant Staphylococcus aureus. Antimicrob. Resist. Infect. Control 2020, 9, 135. [Google Scholar] [CrossRef]
- Hoefer, A.; Boyen, F.; Beierschmitt, A.; Moodley, A.; Roberts, M.; Butaye, P. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts. Antibiotics 2021, 10, 290. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Takemoto, K.; Iwasaki, S.; Yajima, T.; Kido, A.; Yamauchi, A.; Kuroiwa, K.; Kumai, Y.; Yoshihara, S.; Tokumoto, H.; et al. Isolation and characterization of Staphylococcus argenteus strains from retail foods and slaughterhouses in Japan. Int. J. Food Microbiol. 2021, 363, 109503. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antibiotic | MIC or Disk Zone | Interpretation |
|---|---|---|
| Agar diffusion (mm) | ||
| Cefoxitin | 29 | Susceptible § |
| Clindamycin | 28 | Susceptible § |
| Rifampicin | 32 | Susceptible § |
| Trimethoprim | 26 | Susceptible § |
| Automated antibiogram (mg/L) | ||
| Ceftaroline | 0.5 | Susceptible § |
| Chloramphenicol | 8 | Susceptible § |
| Ciprofloxacin | ≤0.5 | Susceptible § |
| Daptomycin | 0.5 | Susceptible § |
| Erythromycin | ≤0.2 | Susceptible § |
| Fusidic acid | ≤0.5 | Susceptible § |
| Gentamycin | ≤1 | Susceptible § |
| Levofloxacin | ≤0.5 | Susceptible § |
| Linezolid | 2 | Susceptible § |
| Moxifloxacin | ≤0.25 | Susceptible § |
| Mupirocin | ≤0.5 | Susceptible § |
| Penicillin | 0.125 | Susceptible § |
| Quinupristine/Dalfopristine | ≤0.5 | Susceptible § |
| Teicoplanin | ≤0.5 | Susceptible § |
| Tetracycline | ≤0.5 | Susceptible § |
| Tigecycline | ≤0.125 | Susceptible § |
| Tobramycin | ≤1 | Susceptible § |
| Trimethoprim/sulfamethoxazole | ≤0.5 | Susceptible § |
| Vancomycin | 1 | Susceptible § |
| Microdilution (mg/L) | ||
| Enrofloxacin | ≤0.25 | Susceptible ‡ |
| Kanamycin | ≤16 | Susceptible ‡ |
| Neomycin | ≤8 | Susceptible ‡ |
| Epsilometer test (mg/L) | ||
| Fosfomycin | 64 | Resistant § |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meijer, E.F.J.; van Renssen, A.; Maat, I.; van der Graaf-van Bloois, L.; Duim, B.; Broens, E.M. Canine Staphylococcus argenteus: Case Report from The Netherlands. Pathogens 2022, 11, 153. https://doi.org/10.3390/pathogens11020153
Meijer EFJ, van Renssen A, Maat I, van der Graaf-van Bloois L, Duim B, Broens EM. Canine Staphylococcus argenteus: Case Report from The Netherlands. Pathogens. 2022; 11(2):153. https://doi.org/10.3390/pathogens11020153
Chicago/Turabian StyleMeijer, Eelco F. J., Anne van Renssen, Ianthe Maat, Linda van der Graaf-van Bloois, Birgitta Duim, and Els M. Broens. 2022. "Canine Staphylococcus argenteus: Case Report from The Netherlands" Pathogens 11, no. 2: 153. https://doi.org/10.3390/pathogens11020153
APA StyleMeijer, E. F. J., van Renssen, A., Maat, I., van der Graaf-van Bloois, L., Duim, B., & Broens, E. M. (2022). Canine Staphylococcus argenteus: Case Report from The Netherlands. Pathogens, 11(2), 153. https://doi.org/10.3390/pathogens11020153