
Patterns of Kdr-L995F Allele Emergence Alongside Detoxifying Enzymes Associated with Deltamethrin Resistance in Anopheles gambiae s.l. from North Cameroon

 ,
,  , , ,
, , ,
Abstract
:1. Background
2. Results
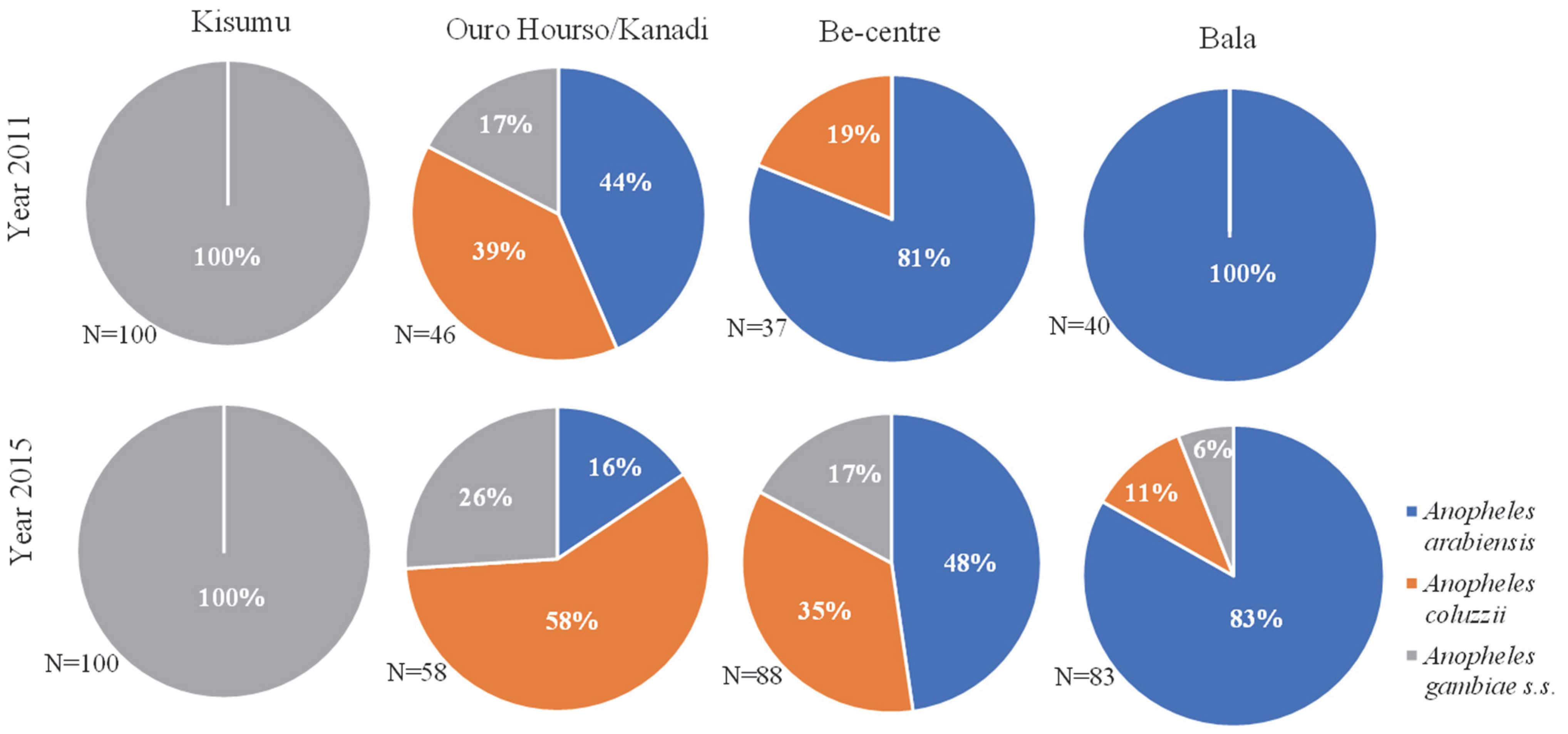
2.1. Distribution of Mosquito Species across the Study Sites
2.2. Deltamethrin Resistance Profiles
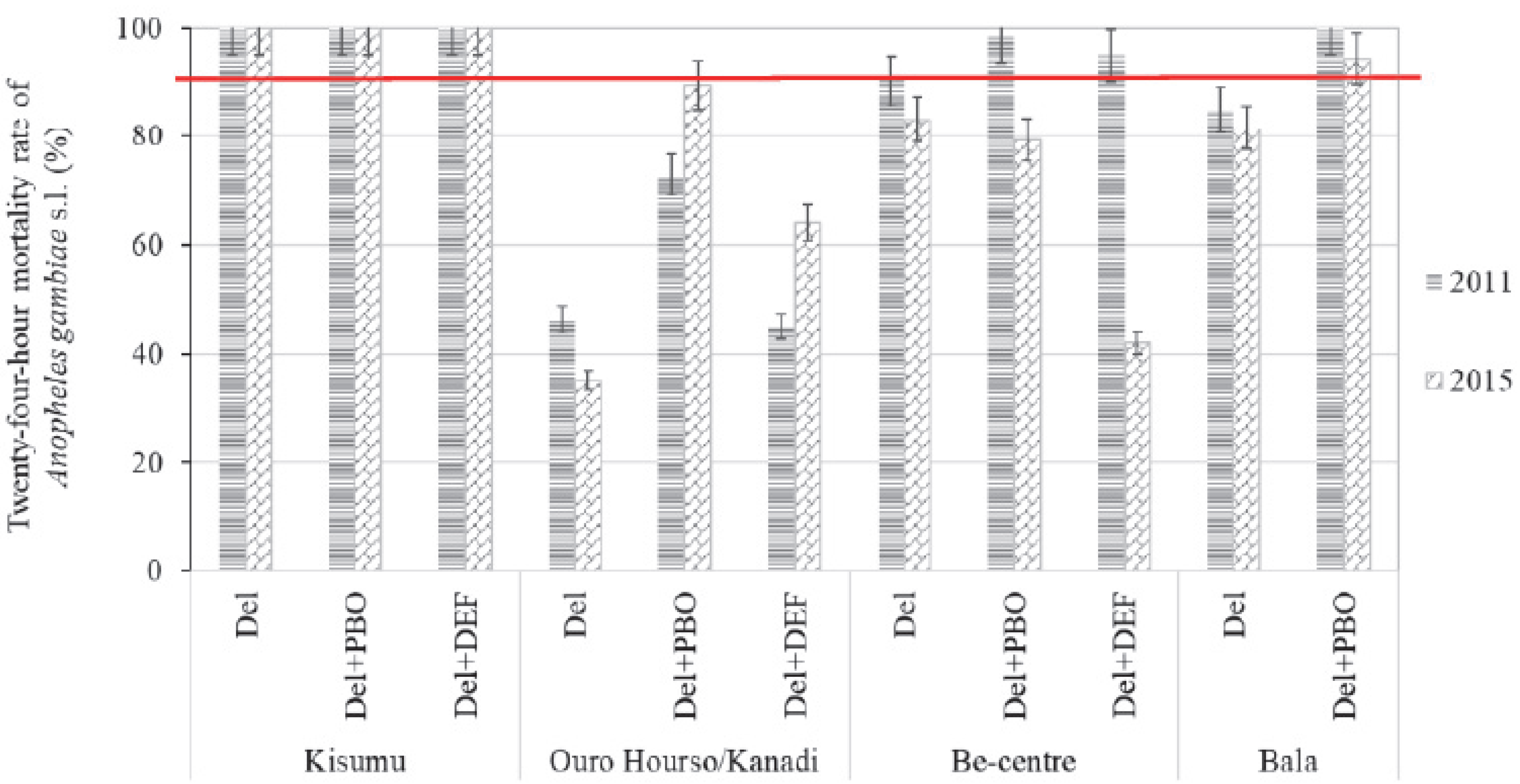
2.3. Metabolic Resistance Indicators: Effects of Synergists on Deltamethrin Resistance
2.3.1. Effects of Synergists on Mosquito Knockdown to Deltamethrin
2.3.2. Effects of Synergists on Mosquito Mortality to Deltamethrin
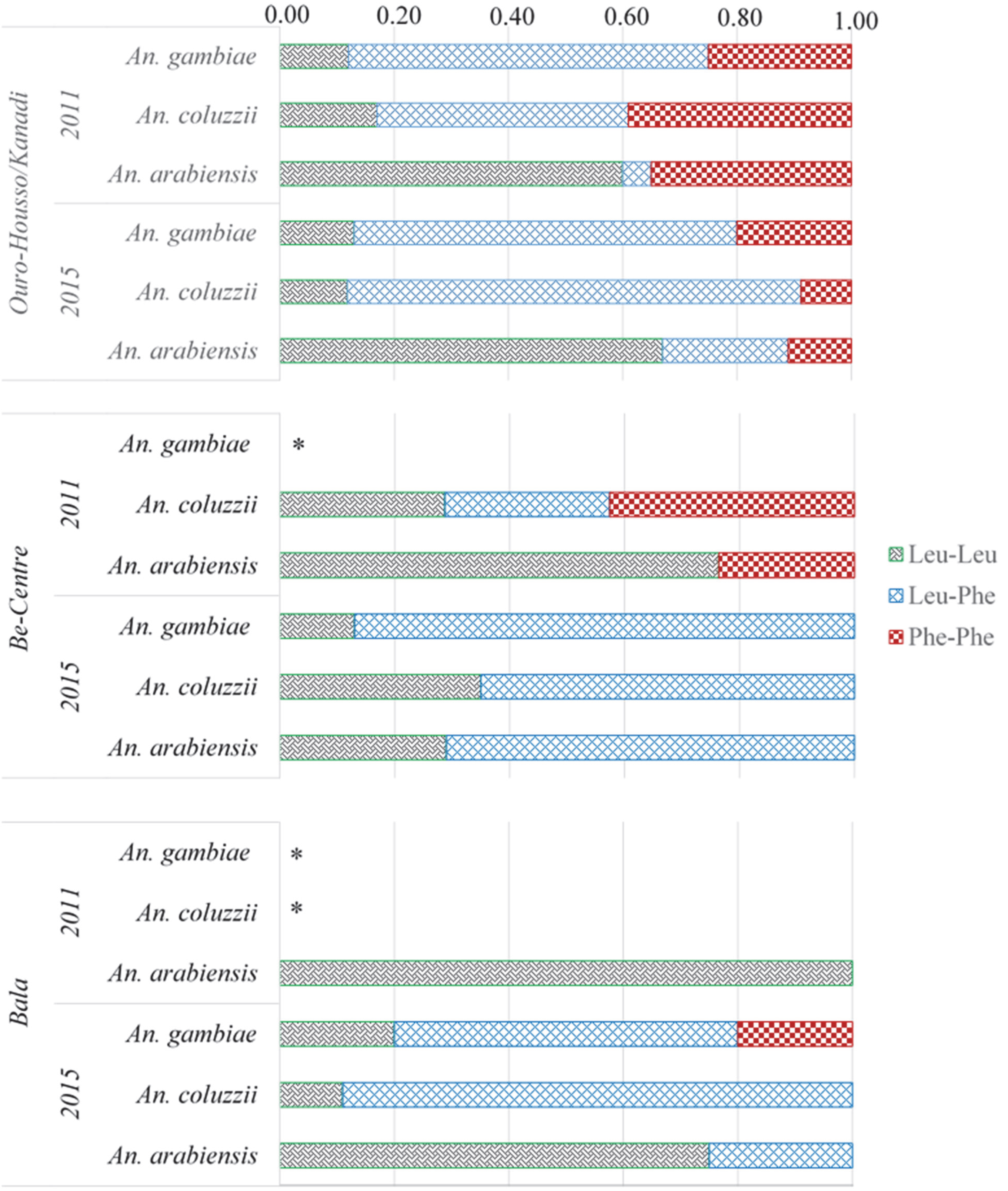
2.4. Genotypic and Allelic Frequencies at the Kdr 995 Locus
2.5. Interactions between Metabolic and kdr 995F Resistance Mechanisms
3. Discussion
4. Materials and Methods
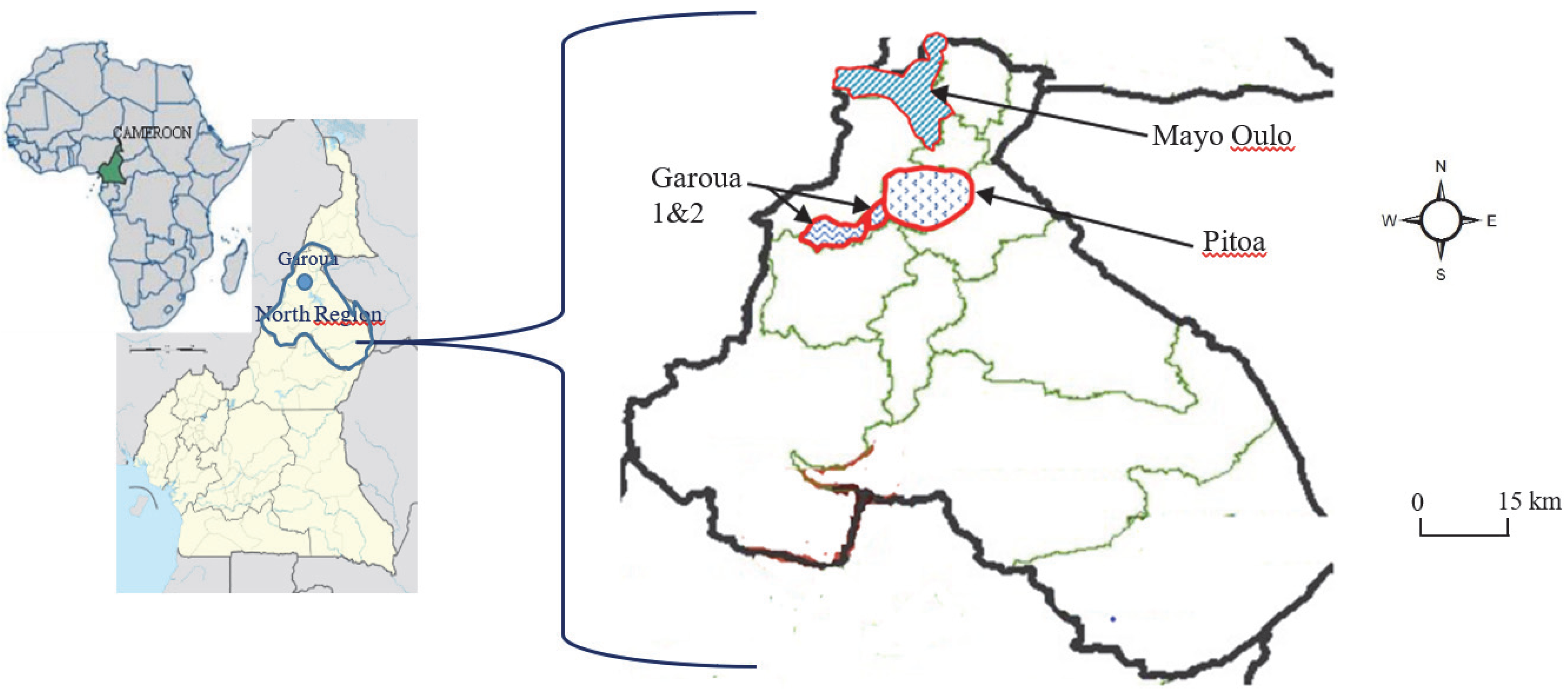
4.1. Study Period and Sites
4.2. Mosquito Collection and Processing
4.3. Bioassays
- One assay with sub-samples of mosquitoes not exposed to synergists prior to susceptibility tests, for establishing their resistance status to deltamethrin;
- Two assays with sub-samples of mosquitoes pre-exposed to synergists for one hour, i.e., piperonyl butoxide (PBO, Sigma Milwaukee, WI) and SSS-tributyl-phosphoro thrithioate (DEF, Sigma Milwaukee, WI), for detection of increased activity of two detoxifying enzymes, namely cytochrome P450 oxidases and carboxylesterases, respectively.
5. Species Identification and Kdr Genotyping
6. Data Analysis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van den Berg, H.; Bezerra, H.S.S.; Al-Eryani, S.; Chanda, E.; Nagpal, B.N.; Knox, T.B.; Velayudhan, R.; Yadav, R.S. Recent trends in global insecticide use for disease vector control and potential implications for resistance management. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Vector-Borne Diseases. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 27 December 2021).
- World Health Organization. World Malaria Report 2021: An in-Depth Update on Global and Regional Malaria Data and Trends. p. 263. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (accessed on 11 January 2022).
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Bertozzi-Villa, A.; Bever, C.A.; Koenker, H.; Weiss, D.J.; Vargas-Ruiz, C.; Nandi, A.K.; Gibson, H.S.; Harris, J.; Battle, K.E.; Rumisha, S.F.; et al. Maps and metrics of insecticide-treated net access, use, and nets-per-capita in Africa from 2000–2020. Nat. Commun. 2021, 12, 3589. [Google Scholar] [CrossRef] [PubMed]
- Riveron, J.M.; Tchouakui, M.; Mugenzi, L.; Menze, B.D.; Chiang, M.C.; Wondji, C.S. Insecticide Resistance in Malaria Vectors: An Update at a Global Scale; Chapter Towards Malaria Elimination; Manguin, S., Ed.; Intech Open: London, UK, 2018; pp. 149–175. [Google Scholar]
- Martinez Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Ins. Molec. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Report on Insecticide Resistance in Malaria Vectors: 2010–2016; World Health Organization: Geneva, Switzerland, 2018; Available online: https://creativecommons.org/licenses/by-nc-sa/3.0/igo (accessed on 27 December 2021).
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Warr, E.; Stevenson, B.J.; Pignatelli, P.M.; Morgan, J.C.; Steven, A. Field-caught permethrin-resistant Anopheles gambiae overexpress CYP6P3, a P450 that metabolises pyrethroids. PLoS Genet. 2008, 4, e1000286. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.J.; Bibby, J.; Pignatelli, P.; Muangnoicharoen, S.; O’Neill, P.M.; Lian, L.Y.; Muller, P.; Nikou, D.; Steven, A.; Hemingway, J.; et al. Cytochrome P450 6M2 from the malaria vector Anopheles gambiae metabolizes pyrethroids: Sequential metabolism of deltamethrin revealed. Insect Biochem. Molec. Biol. 2011, 41, 492–502. [Google Scholar] [CrossRef]
- Vontas, J.; Katsavou, E.; Mavridis, K. Cytochrome P450-based metabolic insecticide resistance in Anopheles and Aedes mosquito vectors: Muddying the waters. Pestic. Biochem. Physiol. 2020, 170, 104666. [Google Scholar] [CrossRef]
- Vulule, J.M.; Beach, R.F.; Atieli, F.K.; McAllister, J.C.; Brogdon, W.G.; Roberts, J.M.; Mwangi, R.W.; Hawley, W.A. Elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae from Kenyan villages using permethrin-impregnated nets. Med. Vet. Entomol. 1999, 13, 239–244. [Google Scholar] [CrossRef]
- Chandor-Proust, A.; Bibby, J.; R’Egent-Kloekner, M.; Roux, J.; Guittard-Crilat, E.; Poupardin, R.; Riaz, M.A.; Paine, M.; Dauphin-Villemant, C.; Reynaud, S.; et al. The central role of mosquito cytochrome P450 CYP6Zs in insecticide detoxification revealed by functional expression and structural modelling. Biochem. J. 2013, 455, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.A.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar]
- Nwane, P.; Etang, J.; Chouaїbou, M.; Toto, J.C.; Koffi, A.; Mimpfoundi, R.; Simar, F. Multiple insecticide resistance mechanisms in Anopheles gambiae s.l. populations from Cameroon, Central Africa. Paras. Vect. 2013, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisinza, W.N.; Nkya, T.E.; Kabula, B.; Overgaard, H.J.; Massue, D.J.; Mageni, Z. Multiple insecticide resistance in Anopheles gambiae from Tanzania: A major concern for malaria vector control. Malar. J. 2017, 16, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oumbouke, W.A.; Pignatelli, P.; Barreaux, A.M.G.; Tia, I.Z.; Koffi, A.A.; Alou, L.P.; Sternberg, E.D.; Thomas, M.B.; Weetman, D.; N’Guessan, R.; et al. Fine scale spatial investigation of multiple insecticide resistance and underlying target-site and metabolic mechanisms in Anopheles gambiae in central Côte d’Ivoire. Sci. Rep. 2020, 10, 15066. [Google Scholar] [CrossRef]
- Idowu, O.A.; Taiwo, A.A.; Olatunbosun, O.A.; Olukayode, A.; Oluwakemi, A.; Romoke, J.T.; AbdullahiIdowu, G.; Taiwo, A.S. Multiple insecticide resistance mechanisms in urban population of Anopheles coluzzii (Diptera: Culicidae) from Lagos, South-West Nigeria. Acta Tropica 2021, in press. [CrossRef]
- Czeher, C.; Labbo, R.; Arzika, I.; Duchemin, J.-B. Evidence of increasing Leu-Phe knockdown resistance mutation in Anopheles gambiae from Niger following a nationwide long-lasting insecticide-treated nets implementation. Malar. J. 2008, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Kleinschmidt, I.; Bradley, J.; Knox, T.B.; Mnzava, A.P.; Kafy, H.T.; Mbogo, C.; Ismail, B.A.; Bigoga, J.D.; Adechoubou, A.; Raghavendra, K.; et al. Implications of Insecticide Resistance Consortium, Implications of insecticide resistance for malaria vector control with long-lasting insecticidal nets: Trends in pyrethroid resistance during a WHO-coordinated multi-country prospective study. Paras. Vect. 2018, 11, 550. [Google Scholar]
- World Health Organisation. Global Plan for Insecticide Resistance Management in Malaria Vectors (GPIRM); WHO: Geneva, Switzerland, 2012; Available online: https://apps.who.int/iris/handle/10665/44846 (accessed on 14 February 2022).
- FY-PMI. The PMI VectorLink Cameroon Annual Entomology Report; The PMI VectorLink Project; Abt Associates: Rockville, ML, USA, 2020; p. 74. [Google Scholar]
- Institut National de la Statistique (INS); ICF. Enquête Démographique et de Santé du Cameroun 2018. Indicateurs Clés. Yaoundé, Cameroun, et Rockville, Maryland, USA, 2019. 2019, p. 60. Available online: http://cdnss.minsante.cm/sites/default/files/eds18.pdf (accessed on 14 February 2022).
- Etang, J.; Fondjo, E.; Chandre, F.; Morlais, I.; Brengues, C.; Nwane, P.; Chouaïbou, M.; Djemaï, A.; Simard, F. First report of the kdr mutations in the malaria vector Anopheles gambiae from Cameroon. Am. J. Trop. Med. Hyg. 2006, 74, 795–797. [Google Scholar] [CrossRef]
- Etang, J.; Manga, L.; Toto, J.C.; Guillet, P.; Fondjo, E.; Chandre, F. Spectrum of metabolic-based resistance to DDT and pyrethroids in Anopheles gambiae s.l. populations from Cameroon. J. Vect. Ecolog. 2007, 32, 123–133. [Google Scholar] [CrossRef]
- Müller, P.; Chouaibou, M.; Pignatelli, P.; Etang, J.; Walker, E.D.; Donnelly, M.J.; Simard, F.; Ranson, H. Pyrethroid tolerance is associated with elevated expression of antioxidants and agricultural practice in Anopheles arabiensis sampled from an area of cotton fields in Northern Cameroon. Mol. Ecol. 2008, 17, 1145–1155. [Google Scholar] [CrossRef]
- Mandeng, S.E.; Awono-Ambene, H.P.; Bigoga, J.D.; Eyisap Ekoko, W.E.; Binyang, J.; Piameu, M.; Mbakop, L.R.; Fesuh, B.N.; Mvondo, N.; Tabue, R.; et al. Spatial and temporal development of deltamethrin resistance in malaria vectors of the Anopheles gambiae complex from North Cameroon. PLoS ONE 2019, 14, e0212024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt, I.; Mnzava, A.P.; Kafy, H.T.; Mbogo, C.; Bashir, A.I.; Bigoga, J.; Adechoubou, A.; Raghavendra, K.; Knox, T.B.; Nkuni, Z.J.; et al. Design of a study to determine the impact of insecticide resistance on malaria vector control: A multi- country investigation. Malar. J. 2015, 14, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Test Procedures: For Insecticide Resistance Monitoring in Malaria Vector Mosquitoes; WHO: Geneva, Switzerland, 2018; p. 56. [Google Scholar]
- Wondji, C.; Simard, F.; Petrarca, V.; Etang, J.; Santolamazza, F.; Della Torre, A.; Fontenille, D. Species and populations of the Anopheles gambiae complex in Cameroon with special emphasis on chromosomal and molecular forms of Anopheles gambiae s.s. J. Med. Entomol. 2005, 42, 998–1005. [Google Scholar] [CrossRef]
- Chouaïbou, M.; Etang, J.; Brévault, T.; Nwane, P.; Kérah Hinzoumbé, C.; Mimpfoundi, R.; Simard, F. The dynamics of insecticide resistance in the malaria vector Anopheles gambiae s.l. from an area of extensive cotton cultivation in Northern Cameroon. Trop. Med. Int. Health. 2008, 13, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Ayala, D.; Kamdem, G.C.; Pombi, M.; Etouna, J.; Ose, K.; Fotsing, J.M.; Fontenille, D.; Besansky, N.J.; Costantini, C. Ecological niche partitioning between Anopheles gambiae molecular forms in Cameroon: The ecological side of speciation. BMC Ecol. 2009, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Gimonneau, G.; Pombi, M.; Choisy, M.; Morand, S.; Dabiré, R.K.; Simard, F. Larval habitat segregation between the molecular forms M and S of Anopheles gambiae in a rice field area of Burkina Faso. Med. Vet. Entomo. 2011, 26, 17. [Google Scholar]
- Etang, J.; Mbida Mbida, A.; Ntonga Akono, P.; Binyang, A.J.; Eboumbou Moukoko, C.E.; Lehman, L.G. Anopheles coluzzii larval habitat and insecticide resistance in the island area of Manoka, Cameroon. BMC Infect. Dis. 2016, 16, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tene Fossog, B.; Antonio-Nkondjio, C.; Kengne, P.; Njiokou, F.; Besansky, N.J.; Costantini, C. Physiological correlates of ecological divergence along an urbanization gardient: Differential tolerance to ammonia among molecular forms of the malaria mosquito Anopheles gambiae. BMC Ecol. 2013, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Nwane, P.; Etang, J.; Chouaibou, M.; Toto, J.C.; Mimpfoundi, R.; Awono-Ambene, H.P.; Kerah Hinzoumbé, C.; Simard, F. Trends in DDT and pyrethroid resistance in Anopheles gambiae s.s. populations from urban and agro-industrial settings in southern Cameroon. BMC Infect Dis. 2009, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Ndjemaï, H.N.M.; Patchoké, S.; Atangana, J.; Etang, J.; Simard, F.; Bilong Bilong, C.F.; Reimer, L.; Cornel, A.; Lanzaro, G.C.; Fondjo, E.; et al. The distribution of insecticide resistance in Anopheles gambiae s.l populations from Cameroon: An update. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 1127–1138. [Google Scholar] [CrossRef] [Green Version]
- Nwane, P.; Etang, J.; Chouaїbou, M.; Toto, J.C.; Mimpfoundi, R.; Simard, F. Kdr-based insecticide resistance in Anopheles gambiae s.s. populations in Cameroon: Spread of the L1014F and L1014S mutations. BMC Res. Notes 2011, 4, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diabaté, A.; Baldet, T.; Chandre, F.; Akogbéto, M.; Guiguemde, T.R.; Darriet, F.; Brengues, C.; Guillet, P.; Hemingway, J.; Small, G.J.; et al. The role of agricultural use of insecticides in resistance to pyrethroids in Anopheles gambiae s.l. in Burkina Faso. Am. J. Trop. Med. Hyg. 2002, 67, 617–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wounang, R.; Ketchoum Ngahane, G.C. Enquête Démographique et de Santé et à Indicateurs Multiples (EDS-MICS). Paludisme 2011, 11, 187–204. Available online: http://onsp.minsante.cm/sites/default/files/publications/28/FR260.pdf (accessed on 3 November 2021).
- Bowen, H. Cameroon Malaria Knowledge, Attitudes, and Practices Progress from 2011 to 2012. Malaria No More, Final Evaluation Report, New York, NY 10016, USA, 2012, p. 45. Available online: https://www.comminit.com/content/cameroon-malaria-knowledge-attitudes-and-practices-progress-2011-2012 (accessed on 14 February 2022).
- Andrews, C.A. Natural Selection, Genetic Drift, and Gene Flow Do Not Act in Isolation in Natural Populations. Nat. Educ. Knowl. 2010, 3, 5. [Google Scholar]
- Platt, N.; Kwiatkowska, R.M.; Irving, H.; Diabate, A.; Dabire, R.; Wondji, C.S. Target site resistance mutations (kdr and RDL), but not metabolic resistance, negatively impact male mating competiveness in the malaria vector Anopheles gambiae. Heredity 2015, 115, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Medjigbodo, A.A.; Djogbenou, L.S.; Djihinto, O.Y.; Akoton, R.B.; Abbey, E.; Kakossou, R.M.; Sonounameto, E.G.; Salavi, E.B.J.; Djossou, L.; Badolo, A. Putative pleiotropic effects of the knockdown resistance (L1014F) allele on the life-history traits of Anopheles gambiae. Malar. J. 2021, 20, 480. [Google Scholar] [CrossRef]
- Etang, J.; Chouaibou, M.; Toto, J.C.; Faye, O.; Manga, L.; Samè-Ekobo, A.; Awono-Ambene, P.; Simard, F. A preliminary test of the protective efficacy of Permethrin-treated bed nets in an area of Anopheles gambiae metabolic resistance to pyrethroids in North Cameroon. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 881–884. [Google Scholar] [CrossRef]
- World Meteorological Organization (WMO). World Weather Information Service-Douala; World Meteorological Organization: Douala, Cameroon, 2018; Retrieved. [Google Scholar]
- Ekoko, W.E.; Awono-Ambene, H.P.; Bigoga, J.; Mandeng, S.E.; Piameu, M.; Nvondo, N.; Toto, J.C.; Nwane, P.; Patchoke, S.; Mbackop, L.R.; et al. Patterns of Anopheline feeding/resting behaviour and Plasmo-dium infections in North Cameroon, 2011-2014: Implications for malaria control. Parasites Vectors 2019, 12, 297. [Google Scholar] [CrossRef] [Green Version]
- Tabue, R.N.; Awono-Ambene, P.; Etang, J.; Atangana, J.; Antonio-Nkondjio, C.; Toto, J.C.; Leke, R.G.F.; Fondjo, E.; Mnzava, A.P.; Knox, T.B.; et al. Role of Anopheles (Cellia) rufipes (Gough, 1910) and other local anophelines in human malaria transmission in the northern savannah of Cameroon: A cross-sectional survey. Parasites Vectors 2017, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Tabue, R.; Alungamoh, B.N.; Zeukeng, F.; Esemu, L.F.; Fodjo Yetgang, A.B.; Kouambeng, C.; Awono-Ambene, H.P.; Fondjo, E.; Achu, D.; Leke, R.G.; et al. Case definition of clinical malaria in children from three health districts in the region of North Cameroon. Biomed. Res. Int. 2019, 9709013. [Google Scholar] [CrossRef] [Green Version]
- Lumsden, W.H.R.; Service, M.W. Mosquito Ecology. Field Sampling Methods. J. Appl. Ecol. 1977, 14, 651. [Google Scholar] [CrossRef]
- Gillies, M.T.; De Meillon, B. The anophelinae of Africa south of the Sahara. Pub. South. Afr. Inst. Med. Res. 1968, 54, 343. [Google Scholar]
- Gillies, M.T.; Coetzee, M. Supplement to the anophelinae of Africa south of the Sahara (afrotropical region). Pub. South Afr. Inst. Med. Res. 1987, 55, 143. [Google Scholar]
- Collins, F.H.; Mendez, M.A.; Razmussen, M.O.; Mehaffey, P.C.; Besansky, N.J.; Finnerty, V. A ribosomal RNA gene probe differentiates member species of Anopheles gambiae complex. Am. J. Trop. Med. Hyg. 1987, 37, 37–41. [Google Scholar] [CrossRef]
- Fanello, C.; Santolamazza, F.; Della Torré, A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complexe by PCR-RFLP. Med. Vet. Entomol. 2002, 16, 461–464. [Google Scholar] [CrossRef]
- Lynd, A.; Ranson, H.; McCall, P.J.; Randle, N.P.; Black, I.V.W.C.; Walker, E.D.; Donnelly, M.J. A simplified high-throughput method for pyrethroid knockdown resistance (kdr) detection in Anopheles gambiae. Malar. J. 2005, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Thomas, A.; Kumar, S.; Pillai, M.M.K. Pyperonyl butoxide as a counter measure for deltamethrin resistance in Culex quinquefasciatus Say. Entomon 1991, 18, 1–10. [Google Scholar]
- Rousset, F. GENEPOP ‘007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Res. 2008, 8, 103–106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality and District | Year | N | Tkd50 (CI), min | Tkd95 (CI), min | Resistance Status |
|---|---|---|---|---|---|
| Kisumu Reference strain | 2011 | 86 | 9.5 (8.4–10.8) | 17.3 (15.7–19.4) | S |
| 2015 | 86 | 9.4 (7.4–11.8) | 18.6 (14.7–19.7) | S | |
| Ouro-Housso/Kanadi Garoua district | 2011 | 93 | 51.5 (30.7–64.2) | >60 | R |
| 2015 | 100 | >60 | >60 | R | |
| Be-Centre Pitoa district | 2011 | 82 | 23.5 (20.7–26.1) | >60 | SR |
| 2015 | 95 | 37.4 (32.3–44.0) | >60 | R | |
| Bala Mayo-Oulo district | 2011 | 75 | 41.2 (37.0–45.3) | >60 | R |
| 2015 | 87 | 27.6 (25.9–29.3) | >60 | R |
| Locality and District | Year | Insecticide | N | Kdt50 (CI) (min) | Kdt95 (CI) (min) | Kdts50 (%) | Resistance Status |
|---|---|---|---|---|---|---|---|
| Kisumu Reference strain | 2011 | Del | 86 | 9.5 (8.4–10.8) | 17.3 (15.7–19.4) | - | S |
| Del+PBO | 88 | 9.2 (7.8–10.8) | 18.4 (17.2–20.4) | 3.2 | S | ||
| Del+DEF | 82 | 8.8 (7.1–9.3) | 16.8 (14.5–20.3) | 7.4 | S | ||
| 2015 | Del | 86 | 9.4 (7.4–11.8) | 18.6 (14.7–19.7) | - | S | |
| Del+PBO | 90 | 8.9 (6.8–10.3) | 19.4 (16.2–21.3) | 5.3 | S | ||
| Del+DEF | 84 | 8.6 (7.1–10.7) | 18.8 (15.5–20.6) | 8.5 | S | ||
| Ouro-Housso/Kanadi Garoua district | 2011 | Del | 93 | 51.5 (30.7–64.2) | >60 | - | R |
| Del+PBO | 81 | 28.7 (21.0–35.0) | >60 | 44.3 | R | ||
| Del+DEF | 70 | 59.1 (47.3–24,854.6) | >60 | <0 | R | ||
| 2015 | Del | 100 | >60 | >60 | - | R | |
| Del+PBO | 68 | 40.3 (35.4–46.8) | >60 | 55.5 | R | ||
| Del+DEF | 74 | 52.4 (45.2–68.7) | >60 | 30.0 | R | ||
| Be-Centre Pitoa district | 2011 | Del | 82 | 23.5 (20.7–26.1) | >60 | - | SR |
| Del+PBO | 70 | 10.8 (6.6–14.6) | 41.8 (28.4–94.4) | 54.0 | S | ||
| Del+DEF | 86 | 6.8 (4.3–8.8) | 24.7 (18.6–42.3) | 71.1 | SR | ||
| 2015 | Del | 95 | 37.4 (32.3–44.0) | >60 | - | R | |
| Del+PBO | 82 | 14.9 (12.0–17.7) | >60 | 60.2 | R | ||
| Del+DEF | 88 | >60 | >60 | <0 | R | ||
| Bala Mayo Oulo district | 2011 | Del | 75 | 41.2 (37.0–45.3) | >60 | - | R |
| Del+PBO | 51 | 38.1 (26.1–101.2) | 50 (23.0–249.0) | 7.5 | S | ||
| 2015 | Del | 87 | 27.6 (25.9–29.3) | >60 | - | R | |
| Del+PBO | 89 | 26.3 (24.6–28.0) | >60 | 4.7 | SR |
| Locality and District | Year | Species | N | Fs L995F (%) | Fc L995F (%) | p Value |
|---|---|---|---|---|---|---|
| Ouro-Housso/Kanadi Garoua district | 2011 | An. arabiensis | 20 | 37.50a | 50.00 | 0.88 |
| An. coluzzii | 18 | 61.11c | ||||
| An. gambiae | 8 | 56.25g | ||||
| 2015 | An. arabiensis | 9 | 22.22a | 45.69 | ||
| An. coluzzii | 34 | 48.53c | ||||
| An. gambiae | 15 | 53.33g | ||||
| Be-Centre Pitoa district | 2011 | An. arabiensis | 30 | 3.33a | 13.5 | 0.02 |
| An. coluzzii | 7 | 57.14c | ||||
| An. gambiae | 0 | - | ||||
| 2015 | An. arabiensis | 42 | 35.71a* | 35.80 | ||
| An. coluzzii | 31 | 32.26c | ||||
| An. gambiae | 15 | 43.33 | ||||
| Bala Mayo-Oulo district | 2011 | An. arabiensis | 40 | 0.00a | 0.00 | 0.01 |
| An. coluzzii | 0 | - | ||||
| An. gambiae | 0 | - | ||||
| 2015 | An. arabiensis | 69 | 12.32a* | 18.10 | ||
| An. coluzzii | 9 | 44.44 | ||||
| An. gambiae | 5 | 50.00 |
| Locality | Year | Species and Kdr L995F freq. | PBO Effect | DEF Effect | Kdr L995F Allele | Resistance Status | ||
|---|---|---|---|---|---|---|---|---|
| Kdts50 | Mrv. | Kdts50 | Mrv. | |||||
| Ouro-Housso/Kanadi | 2011 | Aa **, Ac **, Ag ** | + | + | - | - | +++ | R |
| 2015 | Aa *, Ac **, Ag ** | ++ | ++ | + | + | ++ | R | |
| Be-Centre | 2011 | Aa *, Ac *** | ++ | + | ++ | +/- | ++ | RP |
| 2015 | Aa **, Ac **, Ag ** | ++ | - | - | - | +++ | R | |
| Bala | 2011 | Aa | +/- | + | N/A | N/A | - | R |
| 2015 | Aa *, Ac **, Ag ** | +/- | + | N/A | N/A | +++ | R | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etang, J.; Mandeng, S.E.; Nwane, P.; Awono-Ambene, H.P.; Bigoga, J.D.; Ekoko, W.E.; Binyang, A.J.; Piameu, M.; Mbakop, L.R.; Mvondo, N.; et al. Patterns of Kdr-L995F Allele Emergence Alongside Detoxifying Enzymes Associated with Deltamethrin Resistance in Anopheles gambiae s.l. from North Cameroon. Pathogens 2022, 11, 253. https://doi.org/10.3390/pathogens11020253
Etang J, Mandeng SE, Nwane P, Awono-Ambene HP, Bigoga JD, Ekoko WE, Binyang AJ, Piameu M, Mbakop LR, Mvondo N, et al. Patterns of Kdr-L995F Allele Emergence Alongside Detoxifying Enzymes Associated with Deltamethrin Resistance in Anopheles gambiae s.l. from North Cameroon. Pathogens. 2022; 11(2):253. https://doi.org/10.3390/pathogens11020253
Chicago/Turabian StyleEtang, Josiane, Stanislas Elysée Mandeng, Philippe Nwane, Herman Parfait Awono-Ambene, Jude D. Bigoga, Wolfgang Eyisap Ekoko, Achille Jerome Binyang, Michael Piameu, Lili Ranaise Mbakop, Narcisse Mvondo, and et al. 2022. "Patterns of Kdr-L995F Allele Emergence Alongside Detoxifying Enzymes Associated with Deltamethrin Resistance in Anopheles gambiae s.l. from North Cameroon" Pathogens 11, no. 2: 253. https://doi.org/10.3390/pathogens11020253
APA StyleEtang, J., Mandeng, S. E., Nwane, P., Awono-Ambene, H. P., Bigoga, J. D., Ekoko, W. E., Binyang, A. J., Piameu, M., Mbakop, L. R., Mvondo, N., Tabue, R., Mimpfoundi, R., Toto, J. C., Kleinschmidt, I., Knox, T. B., Mnzava, A. P., Donnelly, M. J., & Fondjo, E. (2022). Patterns of Kdr-L995F Allele Emergence Alongside Detoxifying Enzymes Associated with Deltamethrin Resistance in Anopheles gambiae s.l. from North Cameroon. Pathogens, 11(2), 253. https://doi.org/10.3390/pathogens11020253