
Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice



Abstract
:1. Introduction
2. Results
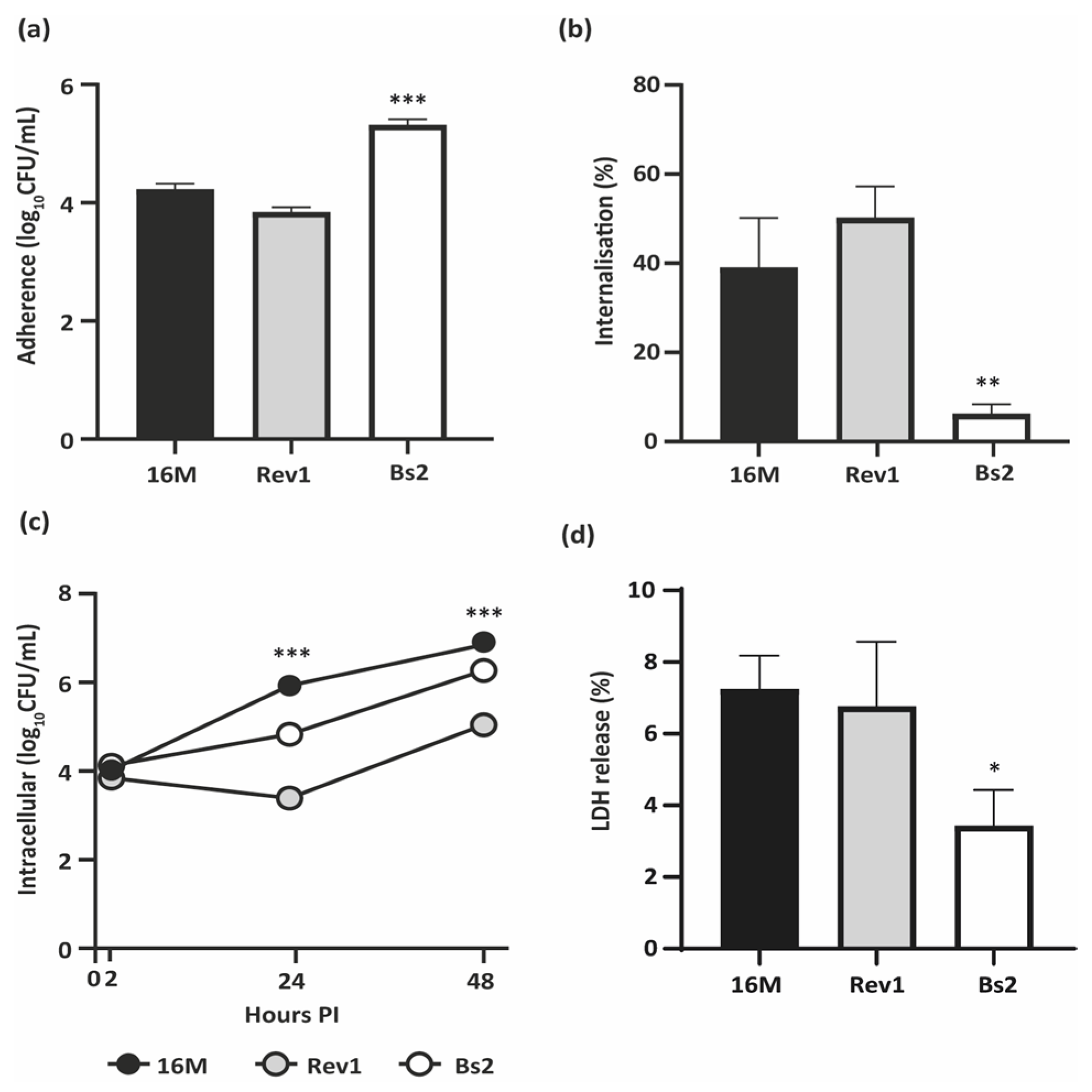
2.1. B. melitensis 16M and Rev1 Internalise More Efficiently and Induce Higher Cytotoxicity in BeWo Cells than B. suis bv2
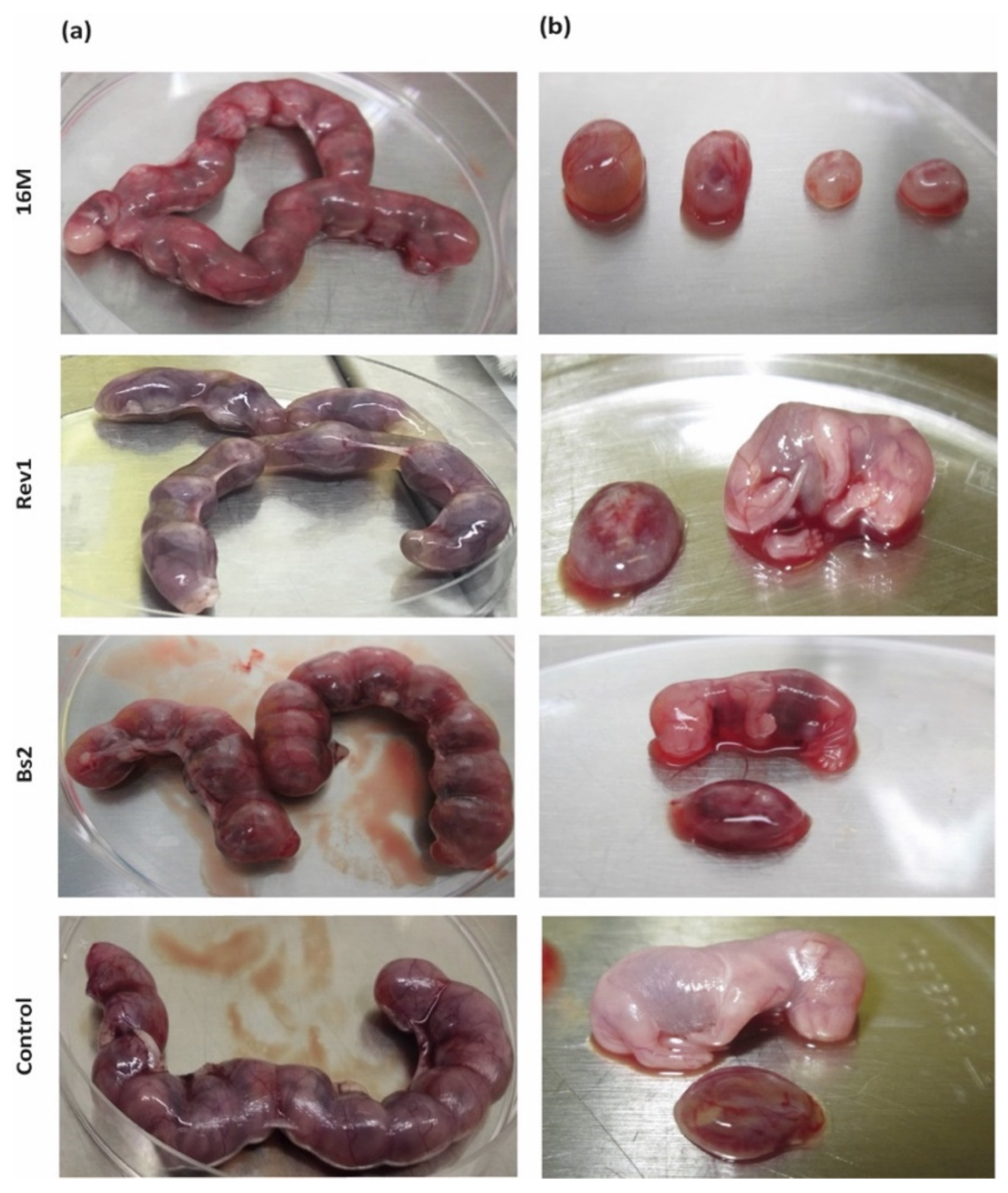
2.2. B. melitensis 16M and Rev1 Exhibit More Tropism for Mouse Placentas Than B. suis bv2
2.3. Kinetics of B. Melitensis and B. suis bv2 Infections in Spleens, Blood, Placentas and Vaginal Shedding and Cytokines Triggered by Infection, along Pregnancy in Mice
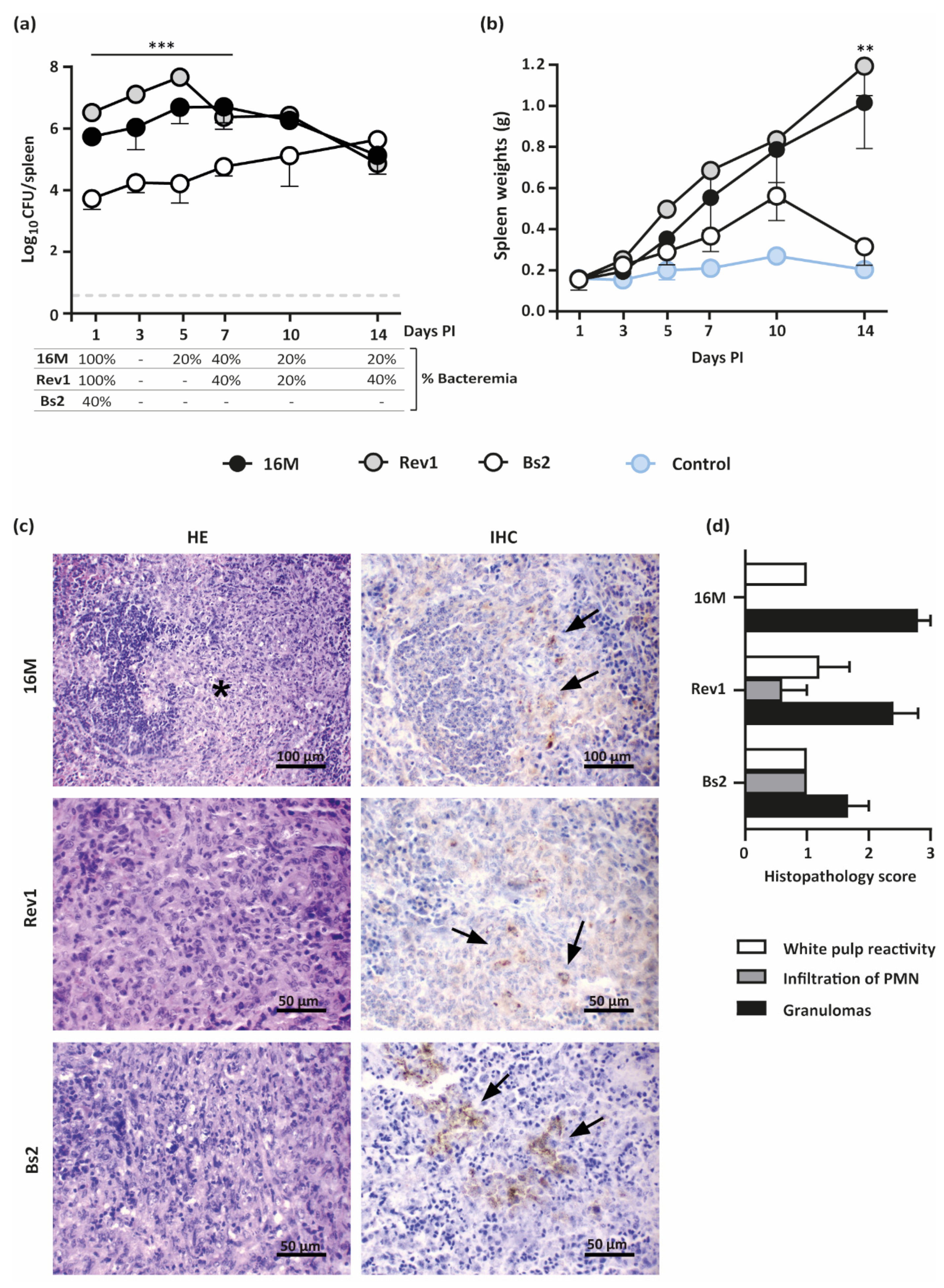
2.3.1. B. melitensis 16M and Rev1 Induce Higher Splenomegaly than B. suis bv2 in Pregnant Mice, Associated to Granulomas
2.3.2. B. melitensis Strains but Not B. suis bv2 Display a Th-1 Mediated Immune Response in Pregnant Mice
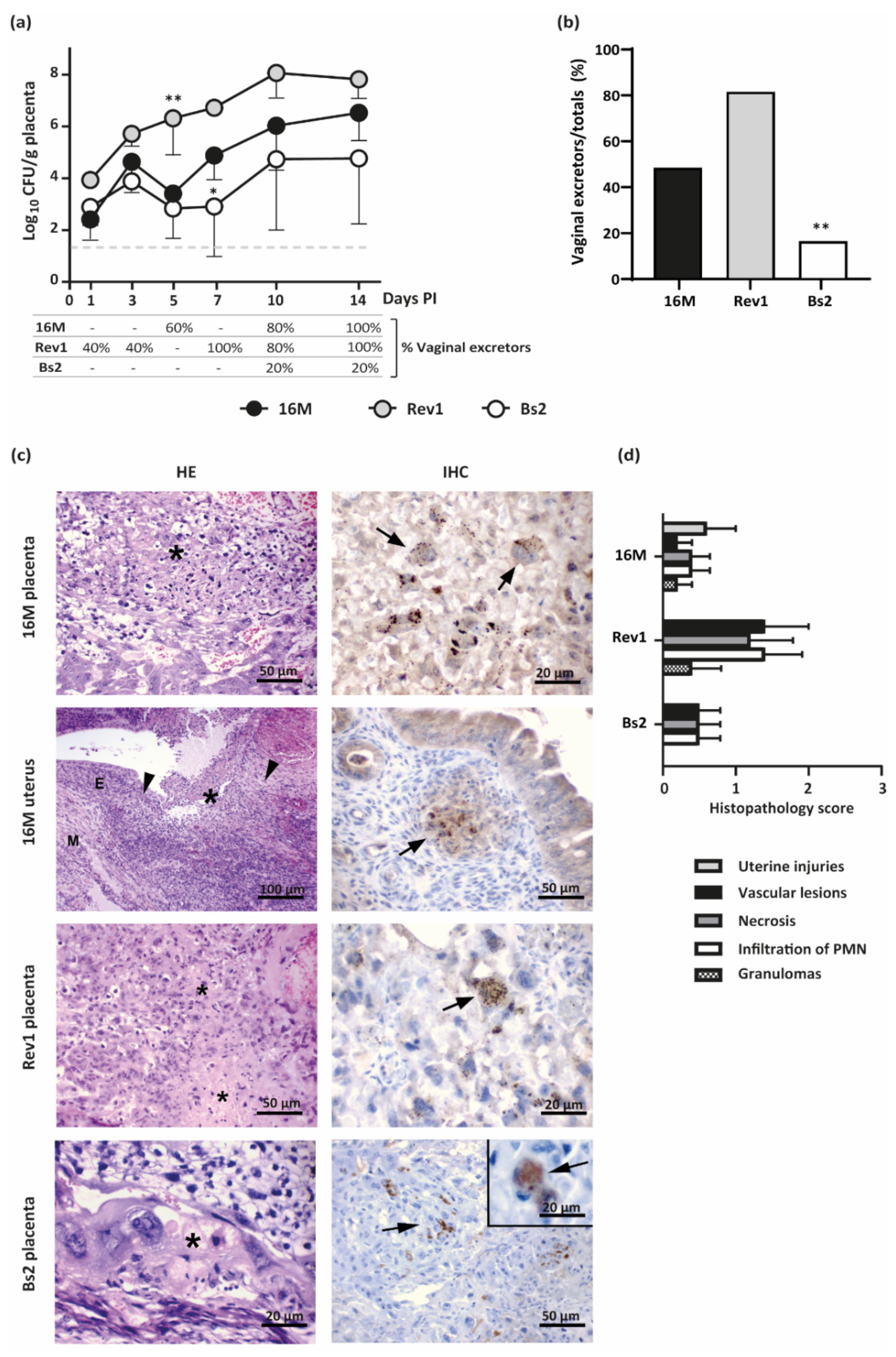
2.3.3. Placental Infections Increase Progressively throughout Pregnancy, More Markedly in B. melitensis than That of B. suis bv2
2.3.4. B. melitensis 16M and Rev1 but Scarcely B. suis bv2 Are Vertically Transmitted and the Infections Affect Foetal Viability
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Media, and Culture Conditions
4.2. BeWo Infections
4.3. Mice Experiments
4.3.1. Ethics and Animal Welfare Statements
4.3.2. Animals and Biosafety
4.3.3. Pregnancies Synchronisation and Experimental Infections of Pregnant Mice
4.3.4. Screening of Infection in Pregnant Mice
4.3.5. Kinetics of Infection during Pregnancy
- (i)
- Blood samples were obtained by retroorbital sinus puncture under anaesthesia and used for bacteraemia detection and for cytokine analysis in serum. For bacteraemia, blood samples were collected with 10% of 50 mM EDTA (Sigma-Aldrich), diluted in PBS-0.1% Triton-100× (Sigma-Aldrich) as described elsewhere [72], and 100 µL were plated in BAB and cultured (37 °C, 5 days).
- (ii)
- Vaginal fluid was collected individually by vaginal washes with 100 µL of sterile PBS previous external decontamination of vulva with povidone–iodine 10% (Betadine®). Vaginal excretion of Brucella was determined by plating of collected fluid in CITA selective medium [73].
- (iii)
- At necropsy, spleens were aseptically removed, individually weighted, and processed. Placental discs were detached from uterine walls and considered as a pool of each dam. When possible, the foetus was dissected from placental envelops tissue. The pool of pups was externally washed with ethanol (70%) and air dried in order to avoid overestimation of foetal infection from amniotic fluid. Each organ or pool was homogenised in sterile PBS, and ten-fold dilutions were plated in BAB to determine viable bacterial counts as previously described [70].
4.4. Serum Cytokine Analysis
4.5. Histopathological Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreno, E. Retrospective and prospective perspectives on zoonotic brucellosis. Front. Microbiol. 2014, 5, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, E.; Moriyón, I. The Genus Brucella. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 315–456. [Google Scholar] [CrossRef]
- Roop, R.M.; Barton, I.S.; Hopersberger, D.; Martin, D.W. Uncovering the Hidden Credentials of Brucella Virulence. Microbiol. Mol. Biol. Rev. 2021, 85, e00021-19. [Google Scholar] [CrossRef] [PubMed]
- Barbier, T.; Machelart, A.; Zúñiga-Ripa, A.; Plovier, H.; Hougardy, C.; Lobet, E.; Willemart, K.; Muraille, E.; De Bolle, X.; Van Schaftingen, E.; et al. Erythritol availability in bovine, murine and human models highlights a potential role for the host aldose reductase during Brucella infection. Front. Microbiol. 2017, 8, 1088. [Google Scholar] [CrossRef] [PubMed]
- MacHelart, A.; Willemart, K.; Zúñiga-Ripa, A.; Godard, T.; Plovier, H.; Wittmann, C.; Moriyón, I.; De Bolle, X.; Van Schaftingen, E.; Letesson, J.J.; et al. Convergent evolution of zoonotic Brucella species toward the selective use of the pentose phosphate pathway. Proc. Natl. Acad. Sci. USA 2020, 117, 26374–26381. [Google Scholar] [CrossRef]
- Letesson, J.J.; Barbier, T.; Zúñiga-Ripa, A.; Godfroid, J.; De Bolle, X.; Moriyón, I. Brucella genital tropism: What’s on the menu. Front. Microbiol. 2017, 8, 506. [Google Scholar] [CrossRef] [Green Version]
- Grilló, M.J.; Blasco, J.M.; Gorvel, J.P.; Moriyán, I.; Moreno, E. What have we learned from brucellosis in the mouse model? Vet. Res. 2012, 43, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Lalsiamthara, J.; Lee, J.H. Development and trial of vaccines against Brucella. J. Vet. Sci. 2017, 18, 281–290. [Google Scholar] [CrossRef]
- Lavazza, A.; Capucci, L. OIE Manual of diagnostic tests and vaccines for terrestrial animals. World Organ. Anim. Health 2018, 3, 355–398. [Google Scholar]
- Silva, T.M.A.; Costa, E.A.; Paixão, T.A.; Tsolis, R.M.; Santos, R.L. Laboratory Animal Models for Brucellosis Research. J. Biomed. Biotechnol. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Dong, S.L.; Watanabe, K.; Furuoka, H.; Suzuki, H.; Watarai, M. Interferon-γ promotes abortion due to Brucella infection in pregnant mice. BMC Microbiol. 2005, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Iwai, N.; Tachibana, M.; Furuoka, H.; Suzuki, H.; Watarai, M. Regulated upon activation normal T-cell expressed and secreted (RANTES) contributes to abortion caused by Brucella abortus infection in pregnant mice. J. Vet. Med. Sci. 2008, 70, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Hasino, M.; Kim, S.; Tachibana, M.; Shumizu, T.; Watari, M. Vertical Transmission of Brucella abortus causes sterility in pregnant mice. J. Vet. Med. Sci. 2012, 74, 1075–1077. [Google Scholar] [CrossRef] [Green Version]
- Keestra-Gounder, A.M.; Byndloss, X.; Seyffert, N.; Young, B.M.; Chávez-Arroyo, A.; Tsai, A.; Cevallos, S.A. NOD1/NOD2 signalig links ER stress with inflamation. Nature 2016, 532, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Byndloss, M.X.; Tsai, A.Y.; Walker, G.T.; Miller, C.N.; Young, B.M.; English, B.C.; Seyffert, N.; Kerrinnes, T.; de Jong, M.F.; Atluri, V.L.; et al. Brucella abortus infection of placental trophoblasts triggers endoplasmic reticulum stress-mediated cell death and fetal loss via type IV secretion system-dependent activation of CHOP. MBio 2019, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, S.P.; Chevrier, N.; Lacerda, T.L.S.; Ben Amara, A.; Gerart, S.; Gorvel, V.A.; De Chastellier, C.; Blasco, J.M.; Mege, J.L.; Gorvel, J.P. Pathogenic brucellae replicate in human trophoblasts. J. Infect. Dis. 2013, 207, 1075–1083. [Google Scholar] [CrossRef]
- Gazzinelli, R.T.; Amichay, D.; Sharton-Kersten, T.; Grunwald, E.; Farber, J.M.; Sher, A. Role of macrophage-derived cytokines in the induction and regulation of cell-mediated immunity to Toxoplasma gondii. Curr. Top. Microbiol. Immunol. 1996, 219, 127–139. [Google Scholar] [CrossRef]
- Quinn, H.E.; Ellis, J.T.; Smith, N.C. Neospora caninum: A cause of immunemediated failure of pregnancy? Trends Parasitol. 2002, 18, 391–394. [Google Scholar] [CrossRef]
- Barber, E.M.; Fazzari, M.; Pollard, J.W. Th1 cytokines are essential for placental immunity to Listeria monocytogenes. Infect. Immun. 2005, 73, 6322–6331. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Cheers, C. Endogenous interleukin-12 is involved in resistance to Brucella abortus infection. Infect. Immun. 1995, 63, 1387–1390. [Google Scholar] [CrossRef] [Green Version]
- Dorneles, E.M.S.; Teixeira-Carvalho, A.; Araújo, M.S.S.; Sriranganathan, N.; Lage, A.P. Immune response triggered by Brucella abortus following infection or vaccination. Vaccine 2015, 33, 3659–3666. [Google Scholar] [CrossRef]
- Raghupathy, R. Th1-type immunity is incompatible with successful pregnancy. Immunol. Today 1997, 18, 478–482. [Google Scholar] [CrossRef]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in pregnancy and recurrent pregnancy losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef]
- Tobias, L.; Cordes, D.O.; Schurig, G.G. Placental pathology of the pregnant mouse inoculated with Brucella abortus strain 2308. Vet. Pathol. 1993, 30, 119–129. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.S.; Wang, G.L.; Wu, T.L.; Lv, Y.L.; Wu, Q.M. A pregnant mouse model for the vertical transmission of Brucella melitensis. Vet. J. 2014, 200, 116–121. [Google Scholar] [CrossRef]
- Blasco, J.M. A review of the use of B. melitensis Rev1 vaccine in adult sheep and goats. Prev. Vet. Med. 1997, 31, 275–283. [Google Scholar] [CrossRef]
- Olsen, S.C.; Tatum, F.M. Swine brucellosis: Current perspectives. Vet. Med. Res. Reports 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegmann, T.G.; Lin, H.; Guilbert, L.; Mosmann, T.R. Bidirectional cytokine interactions in the maternal-fetal relationship: Is successful pregnancy a Th2 phenomenon? Immunol. Today 1993, 14, 353–356. [Google Scholar] [CrossRef]
- Polese, B.; Gridelet, V.; Araklioti, E.; Martens, H.; D’Hauterive, S.P.; Geenen, V. The endocrine milieu and CD4 T-lymphocyte polarization during pregnancy. Front. Endocrinol. 2014, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athanassakis, I.; Iconomidou, B. Cytokine Production in the serum and spleen of mice from day 6 to 14 of gestation: Cytokines/Placenta/Spleen/Serum. Dev. Immunol. 1995, 4, 247–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghupathy, R. Pregnancy: Success and failure within the Th1/Th2/Th3 paradigm. Semin. Immunol. 2001, 13, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Cardoso, P.; Costa-Macedo, G.; Azevedo, V.; Costa-Oliveira, S. Brucella spp noncanonical LPS: Structure, biosynthesis, and interaction with host immune system. Microb. Cell Fact. 2006, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Il-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Guimarães, E.S.; Martins, J.M.; Gomes, M.T.R.; Cerqueira, D.M.; Oliveira, S.C. Lack of interleukin-6 affects ifn-γ and tnf-α production and early in vivo control of Brucella abortus infection. Pathogens 2020, 9, 1040. [Google Scholar] [CrossRef]
- Fernández, A.G.; Ferrero, M.C.; Hielpos, M.S.; Fossati, C.A.; Baldi, P.C. Proinflammatory response of human trophoblastic cells to Brucella abortus infection and upon interactions with infected phagocytes. Biol. Reprod. 2016, 94, 48–49. [Google Scholar] [CrossRef]
- García-Méndez, K.B.; Hielpos, S.M.; Soler-Llorens, P.F.; Arce-Gorvel, V.; Hale, C.; Gorvel, J.P.; O’Callaghan, D.; Keriel, A. Infection by Brucella melitensis or Brucella papionis modifies essential physiological functions of human trophoblasts. Cell. Microbiol. 2019, 21, e13019. [Google Scholar] [CrossRef]
- Detilleux, P.G.; Deyoe, B.L.; Cheville, N.F. Penetration and intracellular growth of Brucella abortus in nonphagocytic cells in vitro. Infect. Immun. 1990, 58, 2320–2328. [Google Scholar] [CrossRef] [Green Version]
- Hensel, M.E.; Garcia-Gonzalez, D.G.; Chaki, S.P.; Hartwig, A.; Gordy, P.W.; Bowen, R.; Ficht, T.A.; Arenas-Gamboa, A.M. Vaccine candidate Brucella melitensis 16MΔvjbR is safe in a pregnant sheep model and confers protection. Ther. Prev. 2020, 5, e00120-20. [Google Scholar] [CrossRef]
- Manterola, L.; Guzmán-Verri, C.; Chaves-Olarte, E.; Barquero-Calvo, E.; De Miguel, M.J.; Moriyón, I.; Grilló, M.J.; López-Goñi, I.; Moreno, E. BvrR/BvrS-controlled outer membrane proteins Omp3a and Omp3b are not essential for Brucella abortus virulence. Infect. Immun. 2007, 75, 4867–4874. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Roldán, E.I.; Avelino-Flores, F.; Dall’Agnol, M.; Freer, E.; Cedillo, L.; Dornand, J.; Girón, J.A. Adherence of Brucella to human epithelial cells and macrophages is mediated by sialic acid residues. Cell. Microbiol. 2004, 6, 435–445. [Google Scholar] [CrossRef]
- Hernández-Castro, R.; Verdugo-Rodríguez, A.; Puente, J.L.; Suárez-Güemes, F. The BMEI0216 gene of Brucella melitensis is required for internalization in HeLa cells. Microb. Pathog. 2007, 44, 28–33. [Google Scholar] [CrossRef]
- Bialer, M.G.; Ferrero, M.C.; Delpino, M.V.; Ruiz-Ranwez, V.; Posadas, D.M.; Baldi, P.C.; Zorreguieta, A. Adhesive functions or pseudogenization of type Va autotransporters in Brucella species. Front. Cell. Infect. Microbiol. 2021, 11, 607610. [Google Scholar] [CrossRef] [PubMed]
- Whatmore, A.M. Current understanding of the genetic diversity of Brucella, an expanding genus of zoonotic pathogens. Infect. Genet. Evol. 2009, 9, 1168–1184. [Google Scholar] [CrossRef] [PubMed]
- Bialer, M.G.; Sycz, G.; González, F.M.; Ferrero, M.C.; Baldi, P.C.; Zorreguieta, A. Adhesins of Brucella: Their roles in the interaction with the host. Pathogens 2020, 9, 942. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.; Williams, A.E.; Pearce, J.H.; Keppie, J.; Harris-Smith, P.W.; Fitz-George, R.B.; Witt, K. Foetal Erythritol: A cause of the localización of Brucella abortus in bovine contagious abortion. Nature 1962, 193, 47–49. [Google Scholar] [CrossRef]
- Bosseray, N. Colonization of mouse placentas by Brucella abortus inoculated during pregnancy. Br. J. Exp. Pathol. 1980, 61, 361–368. [Google Scholar]
- Tachibana, M.; Watanabe, K.; Yamasaki, Y.; Suzuki, H.; Watarai, M. Expression of heme oxygenase-1 is associated with abortion caused by Brucella abortus infection in pregnant mice. Microb. Pathog. 2008, 45, 105–109. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion of the the panel on the animal helath and welfare. Porcine brucellosis (Brucella suis). EFSA J. 2009, 7, 1144. [Google Scholar] [CrossRef] [Green Version]
- Samartino, L.E.; Truax, R.E.; Enright, F.M. Invasion and replication of Brucella abortus in three. J. Vet. Med. Ser. B 1994, 41, 229–236. [Google Scholar] [CrossRef]
- Tibor, A.; Jacques, I.; Guilloteau, L.; Verger, J.M.; Grayon, M.; Wansard, V.; Letesson, J.J. Effect of P39 gene deletion in live Brucella vaccine strains on residual virulence and protective activity in mice. Infect. Immun. 1998, 66, 5561–5564. [Google Scholar] [CrossRef] [Green Version]
- González, D.; Grilló, M.J.; De Miguel, M.J.; Ali, T.; Arce-Gorvel, V.; Delrue, R.M.; Conde-Álvarez, R.; Muñoz, P.; López-Goñi, I.; Iriarte, M.; et al. Brucellosis vaccines: Assessment of Brucella melitensis lipopolysaccharide rough mutants defective in core and O-polysaccharide synthesis and export. PLoS ONE 2008, 3, e2760. [Google Scholar] [CrossRef]
- Sancho, P.; Tejedor, C.; Sidhu-Muñoz, R.S.; Fernández-Lago, L.; Vizcaíno, N. Evaluation in mice of Brucella ovis attenuated mutants for use as live vaccines against B. ovis infection. Vet. Res. 2014, 45, 61. [Google Scholar] [CrossRef] [Green Version]
- Aragón-Aranda, B.; De Miguel, M.J.; Lázaro-Antón, L.; Salvador-Bescós, M.; Zúñiga-Ripa, A.; Moriyón, I.; Iriarte, M.; Muñoz, P.M.; Conde-Álvarez, R. Development of attenuated live vaccine candidates against swine brucellosis in a non-zoonotic B. suis biovar 2 background. Vet. Res. 2020, 51, 92. [Google Scholar] [CrossRef]
- Bustamante, J.J.; Dai, G.; Soares, M.J. Pregnancy and lactation modulate maternal splenic growth and development of the erythroid lineage in the rat and mouse. Reprod. Fertil. Dev. 2008, 20, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Pang, S.C.; Janzen-Pang, J.; Tse, M.Y.; Croy, B.A.; Tse, D. Implant Site Dissections; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780123944450. [Google Scholar]
- Elmore, S.A.; Cochran, R.Z.; Bolon, B.; Lubeck, B.; Mahler, B.; Sabio, D.; Ward, J.M. Histology Atlas of the Developing Mouse Placenta. Toxicol. Pathol. 2022, 50, 60–117. [Google Scholar] [CrossRef]
- Hu, D.; Cross, J.C. Development and function of trophoblast giant cells in the rodent placenta. Int. J. Dev. Biol. 2010, 54, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Cross, J.C. Genetic insights into trophoblast differentiation and placental morphogenesis. Semin. Cell Dev. Biol. 2000, 11, 105–113. [Google Scholar] [CrossRef]
- Durán-Ferrer, M.; Léon, L.; Nielsen, K.; Caporale, V.; Mendoza, J.; Osuna, A.; Perales, A.; Smith, P.; De-Frutos, C.; Gómez-Martín, B.; et al. Antibody response and antigen-specific gamma-interferon profiles of vaccinated and unvaccinated pregnant sheep experimentally infected with Brucella melitensis. Vet. Microbiol. 2004, 100, 219–231. [Google Scholar] [CrossRef]
- Pérez-Sancho, M.; Durán-Ferrer, M.; García-Seco, T.; Macías, P.; García, N.; Martínez, I.; Ruiz, E.; Legaz, E.; Diez-Guerrier, A.; González, S.; et al. Interferon-gamma responses in sheep exposed to virulent and attenuated Brucella melitensis strains. Vet. Immunol. Immunopathol. 2014, 160, 123–128. [Google Scholar] [CrossRef]
- Baldwin, C.L.; Goenka, R. Host immune responses to the intracellular bacteria Brucella: Does the bacteria instruct the host to facilitate chronic infection? Crit. Rev. Immunol. 2006, 26, 407–442. [Google Scholar] [CrossRef]
- Caron, E.; Peyrard, T.; Kohler, S.; Cabane, S.; Liautard, J.P.; Dornand, J. Live Brucella spp. fail to induce tumor necrosis factor alpha excretion upon infection of U937-derived phagocytes. Infect. Immun. 1994, 62, 5267–5274. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Zhang, X.; Wu, X.; Yang, X.; Han, C.; Wang, Z.; Du, Q.; Zhao, X.; Liu, S.L.; Tong, D.; et al. Brucella downregulates tumor necrosis factor-α to promote intracellular survival via Omp25 regulation of different microRNAs in porcine and murine macrophages. Front. Immunol. 2018, 8, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, Y.; Li, Z.; Liu, J.; Shao, X.; Wu, C.; Wang, Y.; Wang, K.; Li, T.; Liu, L.; et al. Outer membrane protein 25 of Brucella activates mitogen-activated protein kinase signal pathway in human trophoblast cells. Front. Vet. Sci. 2017, 4, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteside, E.J.; Boucaut, K.J.; Teh, A.; Garcia-Aragon, J.; Harvey, M.B.; Herington, A.C. Elevated concentration of TNF-α induces trophoblast differentiation in mouse blastocyst outgrowths. Cell Tissue Res. 2003, 314, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Chaouat, G.; Dubanchet, S.; Ledée, N. Cytokines: Important for implantation? J. Assist. Reprod. Genet. 2007, 24, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Robbins, J.R.; Bakardjiev, A.I. Pathogens and the placental fortress. Curr. Opin. Microbiol. 2012, 15, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Kuroda, Y.; Sugiyama, A. A comparison of the histological structure of the placenta in experimental animals. J. Toxicol. Pathol. 2014, 27, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, P.; Boadella, M.; Arnal, M.; de Miguel, M.J.; Revilla, M.; Martínez, D.; Vicente, J.; Acevedo, P.; Oleaga, Á.; Ruiz-Fons, F.; et al. Spatial distribution and risk factors of Brucellosis in berian wild ungulates. BMC Infect. Dis. 2010, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Grilló, M.J.; Manterola, L.; De Miguel, M.J.; Muñoz, P.M.; Blasco, J.M.; Moriyón, I.; López-Goñi, I. Increases of efficacy as vaccine against Brucella abortus infection in mice by simultaneous inoculation with avirulent smooth bvrS/bvrR and rough wbkA mutants. Vaccine 2006, 24, 2910–2916. [Google Scholar] [CrossRef]
- Pattillo, R.A.; Gey, G.O. The establishment of a cell line of human hormone-synthesizing trophoblastic cells in vitro. Cancer Res. 1968, 28, 1231–1236. [Google Scholar]
- Vitry, M.A.; Mambres, D.H.; Deghelt, M.; Hack, K.; Machelart, A.; Lhomme, F.; Vanderwinden, J.M.; Vermeersch, M.; De Trez, C.; Pérez-Morga, D.; et al. Brucella melitensis invades murine erythrocytes during infection. Infect. Immun. 2014, 82, 3927–3938. [Google Scholar] [CrossRef] [Green Version]
- De Miguel, M.J.; Marín, C.M.; Muñoz, P.M.; Dieste, L.; Grilló, M.J.; Blasco, J.M. Development of a selective culture medium for primary isolation of the main Brucella species. J. Clin. Microbiol. 2011, 49, 1458–1463. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Spleen | Reproductive Tract | ||||
|---|---|---|---|---|---|---|
| Strain | Weight (g) | Infection (log10 CFU/g) | Placentas | Foetuses | ||
| Females Infected/Total | Infection (log10 CFU/g) | Females with Pups Infected/Total | Infection (log10 CFU/g) | |||
| B. melitensis 16M | 0.62 ± 0.18 a | 4.47 ± 0.38 a | 4/4 a | 7.71 ± 1.82 a | 4/4 a | 6.37 ± 0.86 a |
| B. melitensis Rev1 | 0.82 ± 0.44 a | 4.52 ± 1.01 a | 4/4 a | 8.17 ± 0.18 a | 4/4 a | 7.75 ± 0.54 a |
| B. suis bv2 | 0.30 ± 0.08 ab | 6.26 ± 0.22 ab | 5/5 a | 4.27 ± 2.06 ab | 1/5 b | 2.05 ± 1.18 b |
| Control | 0.18 ± 0.04 | 1.52 ± 0 | 0/4 | 1.52 ± 0 | 0/4 | 1.52 ± 0 |
| Group | Mouse Number | Litter | Pups Infection | |||
|---|---|---|---|---|---|---|
| Size (No. Pups/Mouse) | Weight (g) | Viability & (%) | Females with Infected Pups/Total | Log10 CFU/g # | ||
| B. melitensis 16M | 1 | Foetal reabsorption | / | 0 | / | 5.25 † |
| 2 | Foetal reabsorption | / | 0 | / | 4.46 † | |
| 3 | 18 | 6.7 | 0 | + | 5.34 | |
| 4 | 16 | 12.4 | 0 | + | 3.28 | |
| 5 | 11 | 12.1 | 0 | + | 3 | |
| mean ± SD | 15.0 ± 3.6 | 10.4 ± 3.2 | 0 a | 3/3 a | 3.87 ± 1.28 a | |
| B. melitensis Rev1 | 1 | 11 | 11.5 | 72.2 | - | 1.52 |
| 2 | 4 | 3.5 | 75 | - | 1.52 | |
| 3 | 11 | 3.1 | 0 | + | 3.45 | |
| 4 | 10 | 9.5 | 30 | + | 2.30 | |
| 5 | 9 | 8.0 | 0 | + | 4.28 | |
| mean ± SD | 9.0 ± 2.6 a | 7.1 ± 3.3 | 35.4 ± 33.0 a | 3/5 a | 2.61 ± 1.09 | |
| B. suis bv2 | 1 | 11 | 6.2 | 81.8 | - | 1.52 |
| 2 | 10 | 5.4 | 50 | + | 5.16 | |
| 3 | 9 | 7.0 | 100 | - | 1.52 | |
| 4 | 14 | 5.2 | 100 | - | 1.52 | |
| 5 | 10 | 6.6 | 100 | - | 1.52 | |
| mean ± SD | 10.8 ± 1.7 | 6.1 ± 0.7 | 86.4 ± 19.5 b | 1/5 b | 2.25 ± 1.46 | |
| Control | 1 | 13 | 7.7 | 100 | - | 1.52 |
| 2 | 14 | 7.3 | 92.8 | - | 1.52 | |
| 3 | 13 | 7.1 | 100 | - | 1.52 | |
| mean ± SD | 13.3 ± 0.6 | 7.4 ± 0.3 | 97.6 ± 4.2 | 0/3 | 1.52 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poveda-Urkixo, I.; Ramírez, G.A.; Grilló, M.-J. Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens 2022, 11, 279. https://doi.org/10.3390/pathogens11030279
Poveda-Urkixo I, Ramírez GA, Grilló M-J. Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens. 2022; 11(3):279. https://doi.org/10.3390/pathogens11030279
Chicago/Turabian StylePoveda-Urkixo, Irati, Gustavo A. Ramírez, and María-Jesús Grilló. 2022. "Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice" Pathogens 11, no. 3: 279. https://doi.org/10.3390/pathogens11030279
APA StylePoveda-Urkixo, I., Ramírez, G. A., & Grilló, M. -J. (2022). Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens, 11(3), 279. https://doi.org/10.3390/pathogens11030279