
Integrated Analysis of the miRNA-mRNA Regulatory Network Involved in HIV-Associated Neurocognitive Disorder


Abstract
:1. Introduction
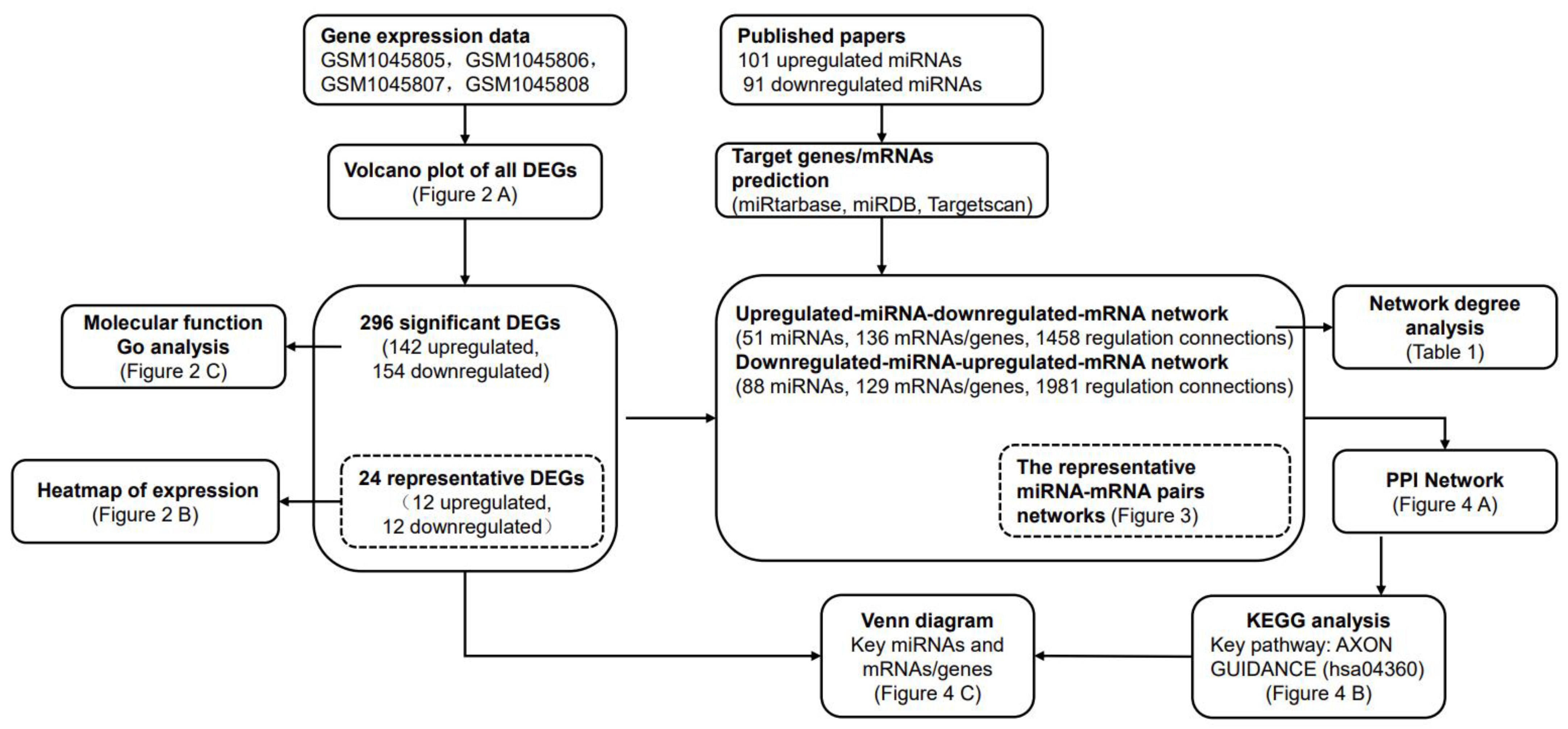
2. Results
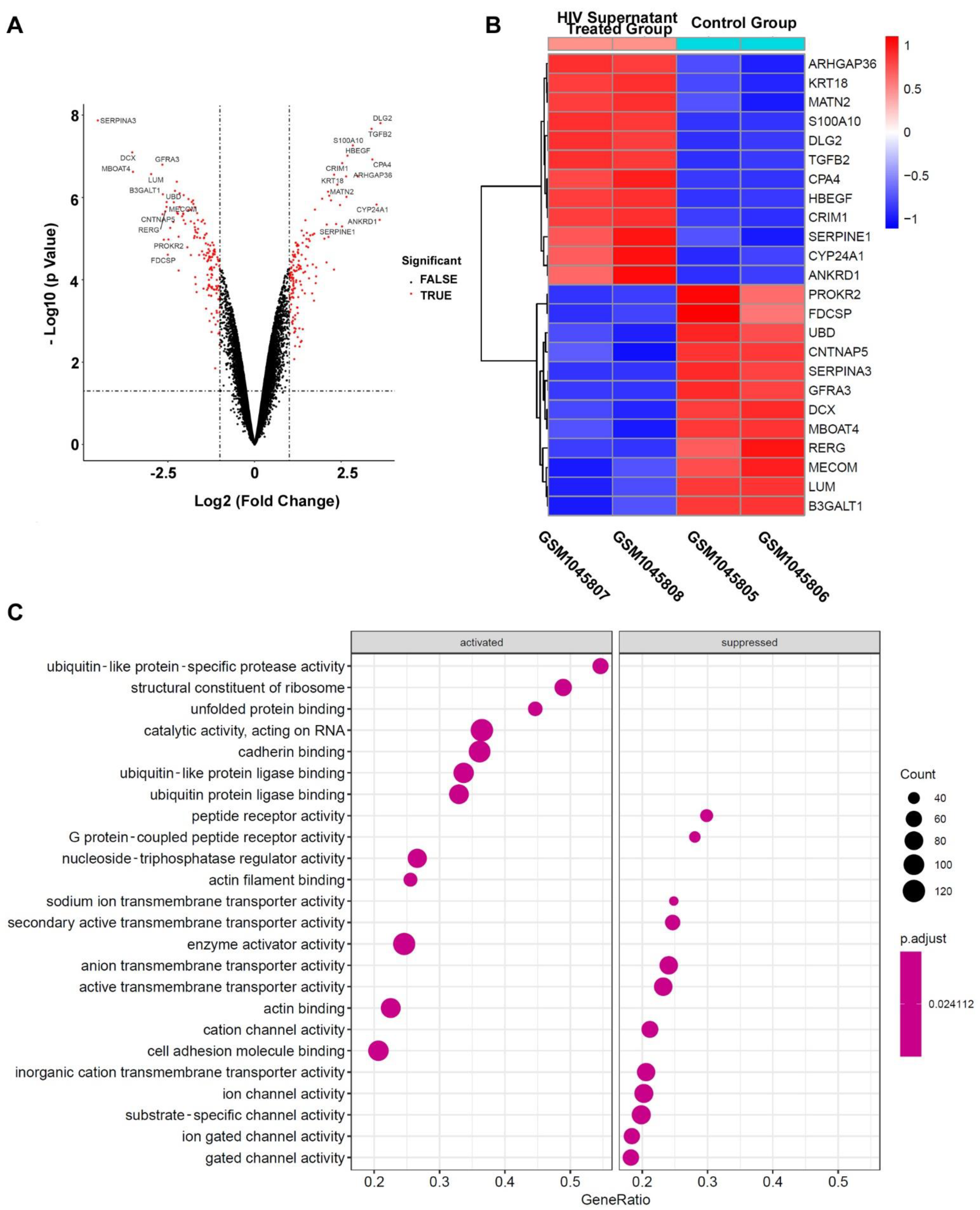
2.1. Identification and Functional Analysis of DEGs Associated with HAND
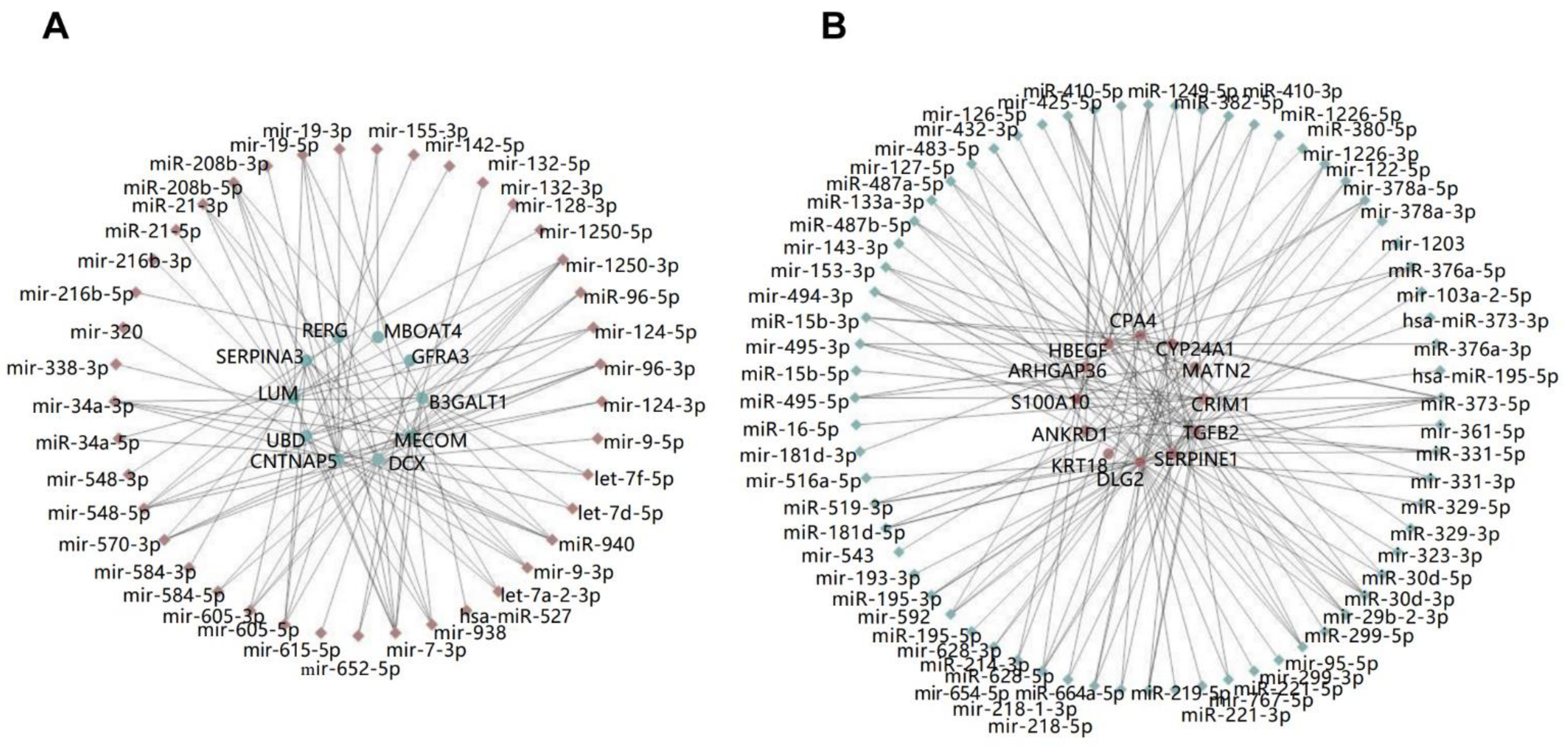
2.2. Construction of the miRNA-mRNA Net Involved in HAND
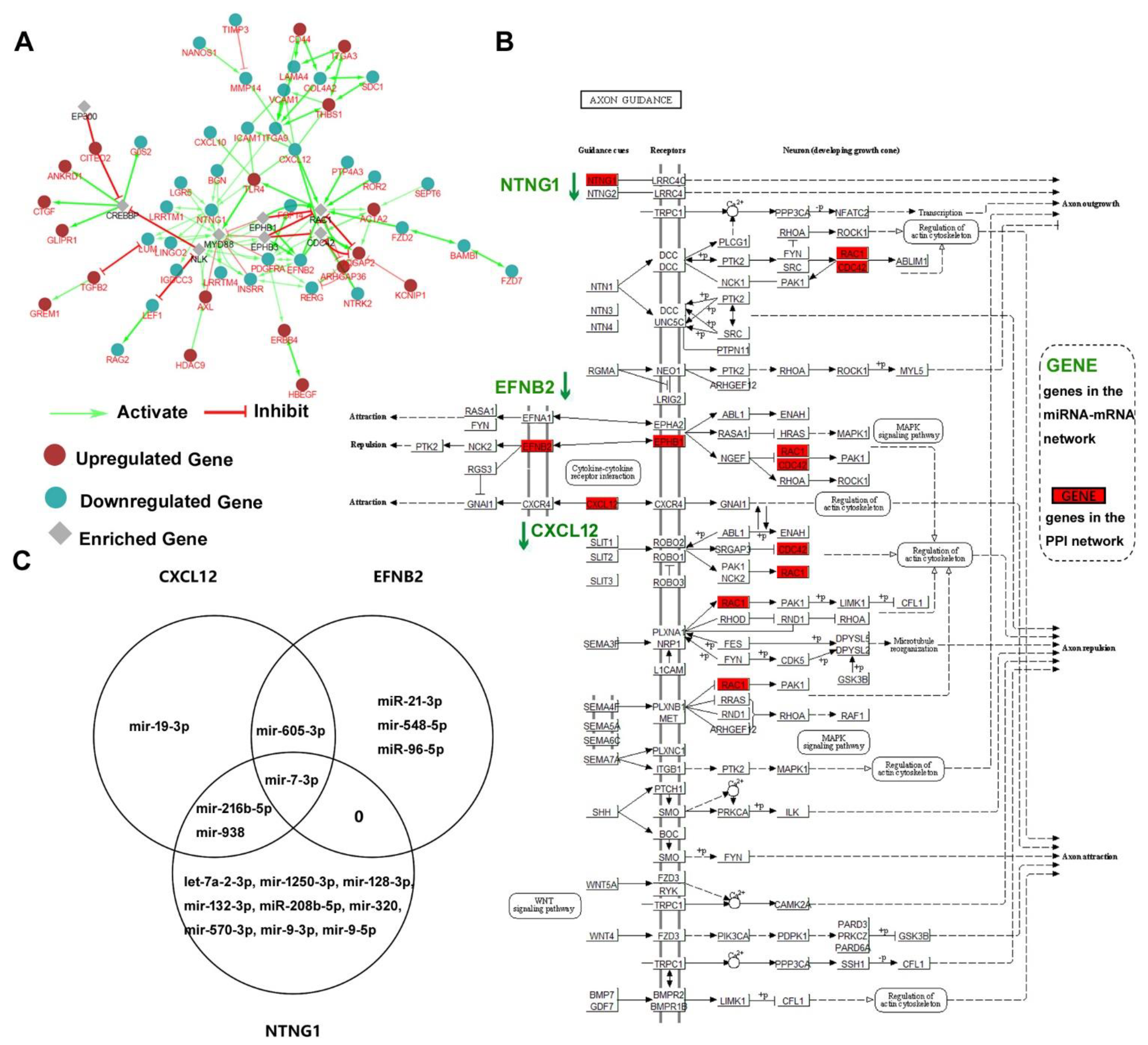
2.3. Functional Exploration of the miRNA-mRNA Net Involved in HAND
3. Discussion
4. Materials and Methods
4.1. GEO Data Processing
4.2. MiRNA Selection and miRNA-Target Gene Prediction
4.3. Construction of miRNA-mRNA Regulatory Network
4.4. Identification of Key miRNAs and mRNAs/Genes in miRNA-mRNA Regulatory Network
4.5. PPI Network Construction
4.6. KEGG Analysis and the Pathway Visualization
4.7. Venn Diagram
5. Conclusions
- 1
- The integrated miRNA-mRNA regulatory network involved in HAND was constructed.
- 2
- Based on the node degree analysis, 20 potential key miRNAs and 20 potential key genes were provided to HAND investigators.
- 3
- The axon guidance pathway (hsa04360), three genes (NTNG1, EFNB2, and CXCL12) and 17 miRNAs were highlighted participating in HAND progression.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moranguinho, I.; Valente, S.T. Block-and-Lock: New Horizons for a Cure for HIV-1. Viruses 2020, 12, 1443. [Google Scholar] [CrossRef] [PubMed]
- Battivelli, E.; Dahabieh, M.S.; Abdel-Mohsen, M.; Svensson, J.P.; Da Silva, I.T.; Cohn, L.B.; Gramatica, A.; Deeks, S.; Greene, W.C.; Pillai, S.K.; et al. Distinct chromatin functional states correlate with HIV latency reactivation in infected primary CD4+ T cells. eLife 2018, 7, e34655. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Hou, J.; Su, B.; Jiang, T.; Guo, C.; Wang, W.; Zhang, Y.; Chang, B.; Wu, H.; Zhang, T. The Prevalence of Frascati-Criteria-Based HIV-Associated Neurocognitive Disorder (HAND) in HIV-Infected Adults: A Systematic Review and Meta-Analysis. Front. Neurol. 2020, 11, 581346. [Google Scholar] [CrossRef] [PubMed]
- Achim, C.L.; Wang, R.; Miners, D.K.; Wiley, C.A. Brain Viral Burden in HIV Infection. J. Neuropathol. Exp. Neurol. 1994, 53, 284–294. [Google Scholar] [CrossRef]
- Figarola-Centurión, I.; Escoto-Delgadillo, M.; González-Enríquez, G.V.; Gutiérrez-Sevilla, J.E.; Vázquez-Valls, E.; Torres-Mendoza, B.M. Sirtuins Modulation: A Promising Strategy for HIV-Associated Neurocognitive Impairments. Int. J. Mol. Sci. 2022, 23, 643. [Google Scholar] [CrossRef]
- Ru, W.; Liu, X.; Bae, C.; Shi, Y.; Walikonis, R.; Chung, J.M.; Tang, S.-J. Microglia Mediate HIV-1 gp120-Induced Synaptic Degeneration in Spinal Pain Neural Circuits. J. Neurosci. 2019, 39, 8408–8421. [Google Scholar] [CrossRef] [Green Version]
- Sachan, V.; Lodge, R.; Mihara, K.; Hamelin, J.; Power, C.; Gelman, B.B.; Hollenberg, M.D.; Cohen, E.A.; Seidah, N.G. HIV-induced neuroinflammation: Impact of PAR1 and PAR2 processing by Furin. Cell Death Differ. 2019, 26, 1942–1954. [Google Scholar] [CrossRef]
- Churchill, M.; Nath, A. Where does HIV hide? A focus on the central nervous system. Curr. Opin. HIV AIDS 2013, 8, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Clifford, D.B. HIV-associated neurocognitive disorder. Curr. Opin. Infect. Dis. 2017, 30, 117–122. [Google Scholar] [CrossRef]
- Omeragic, A.; Kara-Yacoubian, N.; Kelschenbach, J.; Sahin, C.; Cummins, C.L.; Volsky, D.J.; Bendayan, R. Peroxisome Proliferator-Activated Receptor-gamma agonists exhibit anti-inflammatory and antiviral effects in an EcoHIV mouse model. Sci. Rep. 2019, 9, 9428. [Google Scholar] [CrossRef]
- Carvallo, L.; Lopez, L.; Fajardo, J.E.; Jaureguiberry-Bravo, M.; Fiser, A.; Berman, J.W. HIV-Tat regulates macrophage gene expression in the context of neuroAIDS. PLoS ONE 2017, 12, e0179882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, D.; Nagilla, P.; Redinger, C.; Srinivasan, A.; Schatten, G.P.; Ayyavoo, V. Neuronal apoptosis by HIV-1 Vpr: Contribution of proinflammatory molecular networks from infected target cells. J. Neuroinflamm. 2012, 9, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Lu, H.; Ramratnam, B. Neurotoxicity of HIV-1 Tat is attributed to its penetrating property. Sci. Rep. 2020, 10, 14002. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Smith, A.J.; Tan, J.; Shytle, R.D.; Giunta, B. MSM ameliorates HIV-1 Tat induced neuronal oxidative stress via rebalance of the glutathione cycle. Am. J. Transl. Res. 2015, 7, 328–338. [Google Scholar] [PubMed]
- Wallace, D.R. HIV-associated neurotoxicity and cognitive decline: Therapeutic implications. Pharmacol. Ther. 2021, 108047. [Google Scholar] [CrossRef] [PubMed]
- Cull-Candy, S.; Brickley, S.; Farrant, M. NMDA receptor subunits: Diversity, development and disease. Curr. Opin. Neurobiol. 2001, 11, 327–335. [Google Scholar] [CrossRef]
- Hayashi, H.; Campenot, R.B.; Vance, D.E.; Vance, J.E. Apolipoprotein E-Containing Lipoproteins Protect Neurons from Apoptosis via a Signaling Pathway Involving Low-Density Lipoprotein Receptor-Related Protein-1. J. Neurosci. 2007, 27, 1933–1941. [Google Scholar] [CrossRef]
- Chang, L.; Wang, G.-J.; Volkow, N.D.; Ernst, T.; Telang, F.; Logan, J.; Fowler, J.S. Decreased brain dopamine transporters are related to cognitive deficits in HIV patients with or without cocaine abuse. NeuroImage 2008, 42, 869–878. [Google Scholar] [CrossRef] [Green Version]
- Rao, V.R.; Ruiz, A.P.; Prasad, V.R. Viral and cellular factors underlying neuropathogenesis in HIV associated neurocognitive disorders (HAND). AIDS Res. Ther. 2014, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Catani, M.V.; Corasaniti, M.T.; Navarra, M.; Nisticò, G.; Finazzi-Agrò, A.; Melino, G. gp120 Induces Cell Death in Human Neuroblastoma Cells Through the CXCR4 and CCR5 Chemokine Receptors. J. Neurochem. 2002, 74, 2373–2379. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jones, M.; Hingtgen, C.M.; Bu, G.; Laribee, R.N.; Tanzi, R.E.; Moir, R.; Nath, A.; He, J.J. Uptake of HIV-1 Tat protein mediated by low-density lipoprotein receptor-related protein disrupts the neuronal metabolic balance of the receptor ligands. Nat. Med. 2000, 6, 1380–1387. [Google Scholar] [CrossRef]
- Eugenin, E.A.; King, J.E.; Nath, A.; Calderon, T.M.; Zukin, R.S.; Bennett, M.V.L.; Berman, J.W. HIV-tat induces formation of an LRP–PSD-95– NMDAR–nNOS complex that promotes apoptosis in neurons and astrocytes. Proc. Natl. Acad. Sci. USA 2007, 104, 3438–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.A.; Worrad, A.H.; Singh, M.V.; Maggirwar, S.B.; Singh, V.B. Human immunodeficiency virus-1 Tat exerts its neurotoxic effects by downregulating Sonic hedgehog signaling. J. NeuroVirology 2022. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy, P.; Thangaraj, A.; Guo, M.-L.; Hu, G.; Callen, S.; Buch, S. Epigenetic Promoter DNA Methylation of miR-124 Promotes HIV-1 Tat-Mediated Microglial Activation via MECP2-STAT3 Axis. J. Neurosci. 2018, 38, 5367–5383. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Niu, F.; Liao, K.; Periyasamy, P.; Sil, S.; Liu, J.; Dravid, S.M.; Buch, S. HIV-1 Tat-Induced Astrocytic Extracellular Vesicle miR-7 Impairs Synaptic Architecture. J. Neuroimmune Pharmacol. 2019, 15, 538–553. [Google Scholar] [CrossRef]
- Zhou, L.; Pupo, G.M.; Gupta, P.; Liu, B.; Tran, S.L.; Rahme, R.; Wang, B.; Rua, R.; Rizos, H.; Carroll, A.; et al. A parallel genome-wide mRNA and microRNA profiling of the frontal cortex of HIV patients with and without HIV-associated dementia shows the role of axon guidance and downstream pathways in HIV-mediated neurodegeneration. BMC Genom. 2012, 13, 677. [Google Scholar] [CrossRef] [Green Version]
- Santerre, M.; Bagashev, A.; Gorecki, L.; Lysek, K.Z.; Wang, Y.; Shrestha, J.; Del Carpio-Cano, F.; Mukerjee, R.; Sawaya, B.E. HIV-1 Tat protein promotes neuronal dysregulation by inhibiting E2F transcription factor 3 (E2F3). J. Biol. Chem. 2019, 294, 3618–3633. [Google Scholar] [CrossRef] [Green Version]
- Pulliam, L.; Sun, B.; Mustapic, M.; Chawla, S.; Kapogiannis, D. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer’s disease. J. NeuroVirol. 2019, 25, 702–709. [Google Scholar] [CrossRef]
- Ke, X.-S.; Liu, C.-M.; Liu, D.-P.; Liang, C.-C. MicroRNAs: Key participants in gene regulatory networks. Curr. Opin. Chem. Biol. 2003, 7, 516–523. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podolska, A.; Kaczkowski, B.; Busk, P.K.; Søkilde, R.; Litman, T.; Fredholm, M.; Cirera, S. MicroRNA Expression Profiling of the Porcine Developing Brain. PLoS ONE 2011, 6, e14494. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.-B. Posttranscriptional control of neuronal development by microRNA networks. Trends Neurosci. 2008, 31, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fineberg, S.; Kosik, K.S.; Davidson, B. MicroRNAs Potentiate Neural Development. Neuron 2009, 64, 303–309. [Google Scholar] [CrossRef] [Green Version]
- McNeill, E.; Van Vactor, D. MicroRNAs Shape the Neuronal Landscape. Neuron 2012, 75, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Rahimian, P.; He, J.J. HIV-1 Tat-shortened neurite outgrowth through regulation of microRNA-132 and its target gene expression. J. Neuroinflamm. 2016, 13, 247. [Google Scholar] [CrossRef] [Green Version]
- Bagashev, A.; Mukerjee, R.; Santerre, M.; Del Carpio-Cano, F.E.; Shrestha, J.; Wang, Y.; He, J.J.; Sawaya, B.E. Involvement of miR-196a in HIV-associated neurocognitive disorders. Apoptosis Int. J. Program. Cell Death 2014, 19, 1202–1214. [Google Scholar] [CrossRef]
- Chaudhuri, A.D.; Yelamanchili, S.V.; Fox, H.S. MicroRNA-142 Reduces Monoamine Oxidase A Expression and Activity in Neuronal Cells by Downregulating SIRT1. PLoS ONE 2013, 8, e79579. [Google Scholar] [CrossRef] [Green Version]
- Yelamanchili, S.V.; Chaudhuri, A.D.; Chen, L.-N.; Xiong, H.; Fox, H.S. MicroRNA-21 dysregulates the expression of MEF2C in neurons in monkey and human SIV/HIV neurological disease. Cell Death Dis. 2010, 1, e77. [Google Scholar] [CrossRef]
- Tatro, E.T.; Hefler, S.; Shumaker-Armstrong, S.; Soontornniyomkij, B.; Yang, M.; Yermanos, A.; Wren, N.; Moore, D.J.; Achim, C.L. Modulation of BK Channel by MicroRNA-9 in Neurons After Exposure to HIV and Methamphetamine. J. Neuroimmune Pharmacol. 2013, 8, 1210–1223. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Cai, Y.; Lin, Y.; Guan, R.; Xiao, G.; Yang, J. MiR-133b-5p regulates the expression of the heat shock protein 70 during rat neuronal cell apoptosis induced by the gp120 V3 loop peptide. J. Med. Virol. 2015, 88, 437–447. [Google Scholar] [CrossRef]
- Asahchop, E.L.; Branton, W.G.; Krishnan, A.; Chen, P.A.; Yang, D.; Kong, L.; Zochodne, D.W.; Brew, B.J.; Gill, M.J.; Power, C. HIV-associated sensory polyneuropathy and neuronal injury are associated with miRNA–455-3p induction. JCI Insight 2018, 3, e122450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eletto, D.; Russo, G.; Passiatore, G.; Del Valle, L.; Giordano, A.; Khalili, K.; Gualco, E.; Peruzzi, F. Inhibition of SNAP25 expression by HIV-1 Tat involves the activity of mir-128a. J. Cell. Physiol. 2008, 216, 764–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.R.; Mukerjee, R.; Bagashev, A.; Del Valle, L.; Chabrashvili, T.; Hawkins, B.J.; He, J.J.; Sawaya, B.E. HIV-1 Tat Protein Promotes Neuronal Dysfunction through Disruption of MicroRNAs. J. Biol. Chem. 2011, 286, 41125–41134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukerjee, R.; Chang, J.R.; Del Valle, L.; Bagashev, A.; Gayed, M.M.; Lyde, R.B.; Hawkins, B.J.; Brailoiu, E.; Cohen, E.; Power, C.; et al. Deregulation of microRNAs by HIV-1 Vpr Protein Leads to the Development of Neurocognitive Disorders. J. Biol. Chem. 2011, 286, 34976–34985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2017, 46, D296–D302. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 1–10. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA–target interactions—Beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ransohoff, R.M. Multiple roles of chemokine CXCL12 in the central nervous system: A migration from immunology to neurobiology. Prog. Neurobiol. 2008, 84, 116–131. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, T.; Kong, Y.; Chiarella, J.; Price, R.W.; Chaudhury, R.; Liu, X.; Spudich, S.; Robertson, K.; Emu, B.; Lu, L. Exosomal MicroRNAs Associate With Neuropsychological Performance in Individuals With HIV Infection on Antiretroviral Therapy. JAIDS J. Acquir. Immune Defic. Syndr. 2019, 82, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Ye, G. Microarray analysis for the identification of specific proteins and functional modules involved in the process of hepatocellular carcinoma originating from cirrhotic liver. Mol. Med. Rep. 2018, 17, 5619–5626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. 4), S11. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated miRNAs | Downregulated miRNAs | Upregulated mRNAs/Genes | Downregulated mRNAs/Genes |
|---|---|---|---|
| mir-570-3p | mir-95-5p | LPP | CNNM2 |
| mir-7-3p | miR-373-5p | STX7 | FUT9 |
| mir-1250-3p | mir-126-5p | DCLK1 | SLC8A1 |
| miR-940 | miR-30d-3p | CCDC80 | CLSTN2 |
| mir-548-5p | miR-195-3p | ERBB4 | IGSF3 |
| miR-208b-5p | miR-329-5p | CADM2 | NEBL |
| mir-9-3p | mir-29b-2-5p | PARVA | RAB27B |
| mir-19-5p | miR-519-3p | NRXN3 | CACNA1C |
| mir-548-3p | mir-103a-2-5p | PLA2G16 | DCX |
| mir-124-5p | miR-380-3p | DNAJB4 | BAMBI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Ding, Q.; Kong, X. Integrated Analysis of the miRNA-mRNA Regulatory Network Involved in HIV-Associated Neurocognitive Disorder. Pathogens 2022, 11, 407. https://doi.org/10.3390/pathogens11040407
Liu C, Ding Q, Kong X. Integrated Analysis of the miRNA-mRNA Regulatory Network Involved in HIV-Associated Neurocognitive Disorder. Pathogens. 2022; 11(4):407. https://doi.org/10.3390/pathogens11040407
Chicago/Turabian StyleLiu, Chang, Qian Ding, and Xiaohong Kong. 2022. "Integrated Analysis of the miRNA-mRNA Regulatory Network Involved in HIV-Associated Neurocognitive Disorder" Pathogens 11, no. 4: 407. https://doi.org/10.3390/pathogens11040407
APA StyleLiu, C., Ding, Q., & Kong, X. (2022). Integrated Analysis of the miRNA-mRNA Regulatory Network Involved in HIV-Associated Neurocognitive Disorder. Pathogens, 11(4), 407. https://doi.org/10.3390/pathogens11040407