
Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
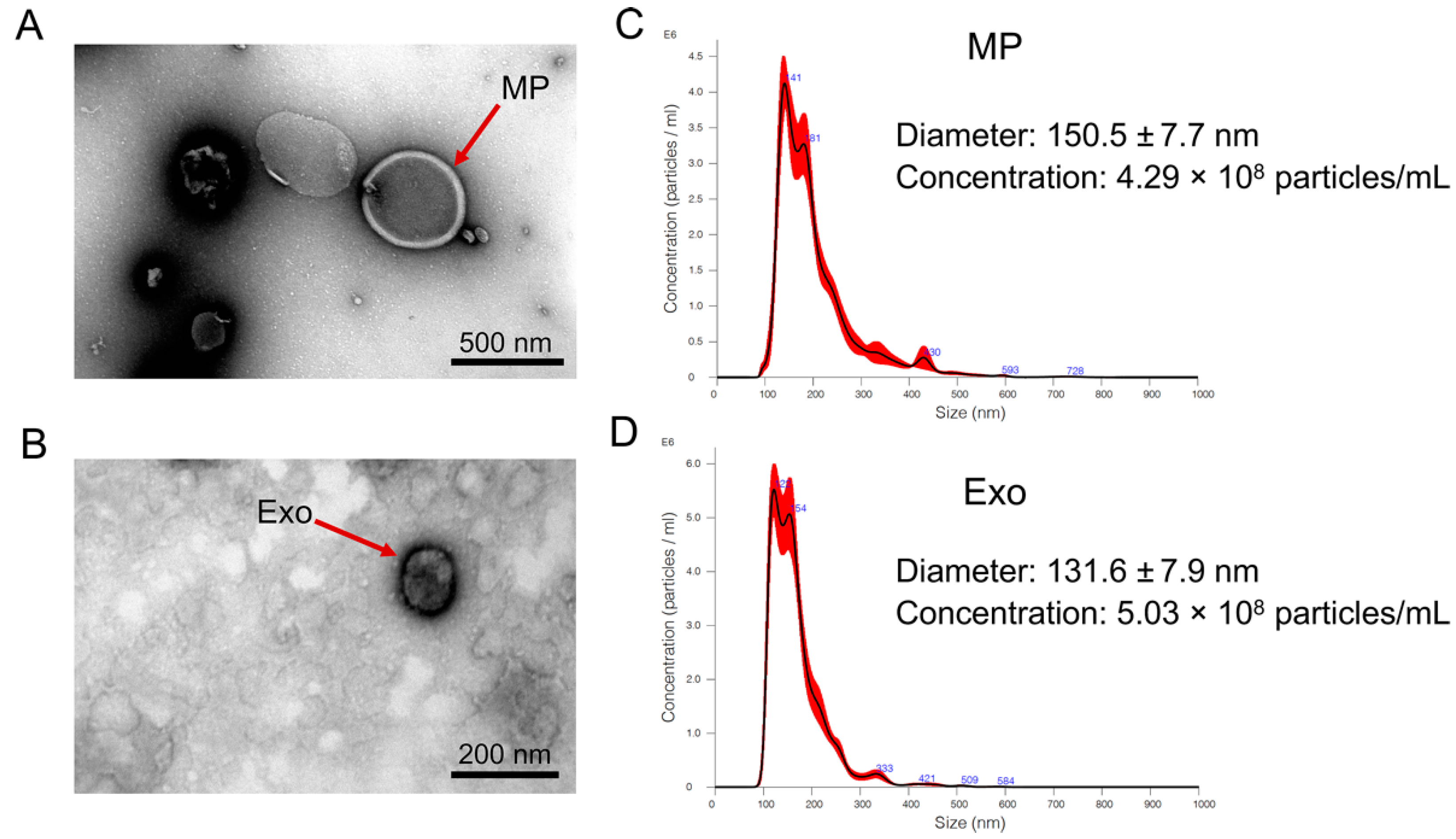
2.1. Isolation and Characterization of N. fowleri-Derived EVs
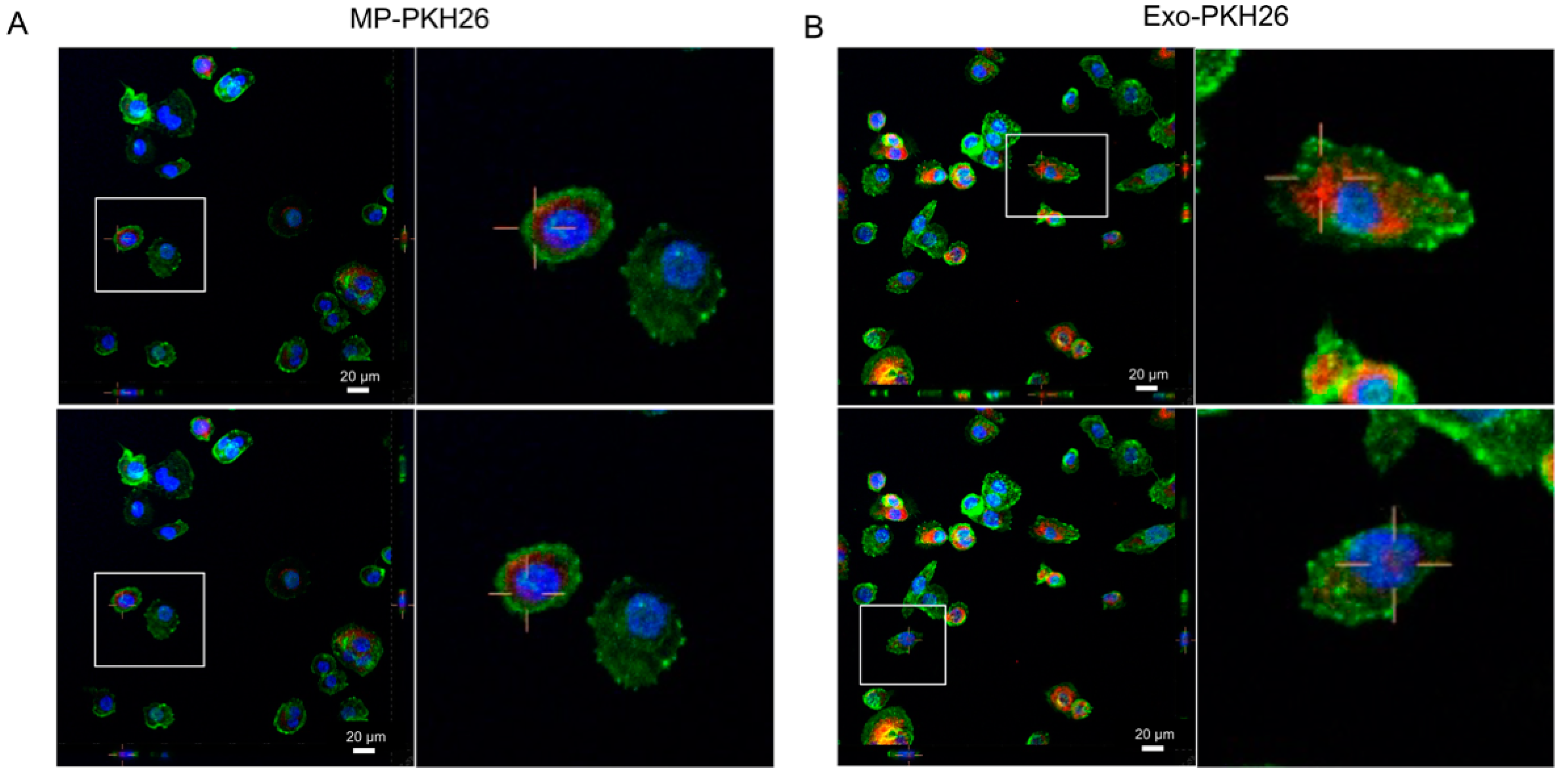
2.2. Internalization of Nf-EVs by THP-1 Macrophages
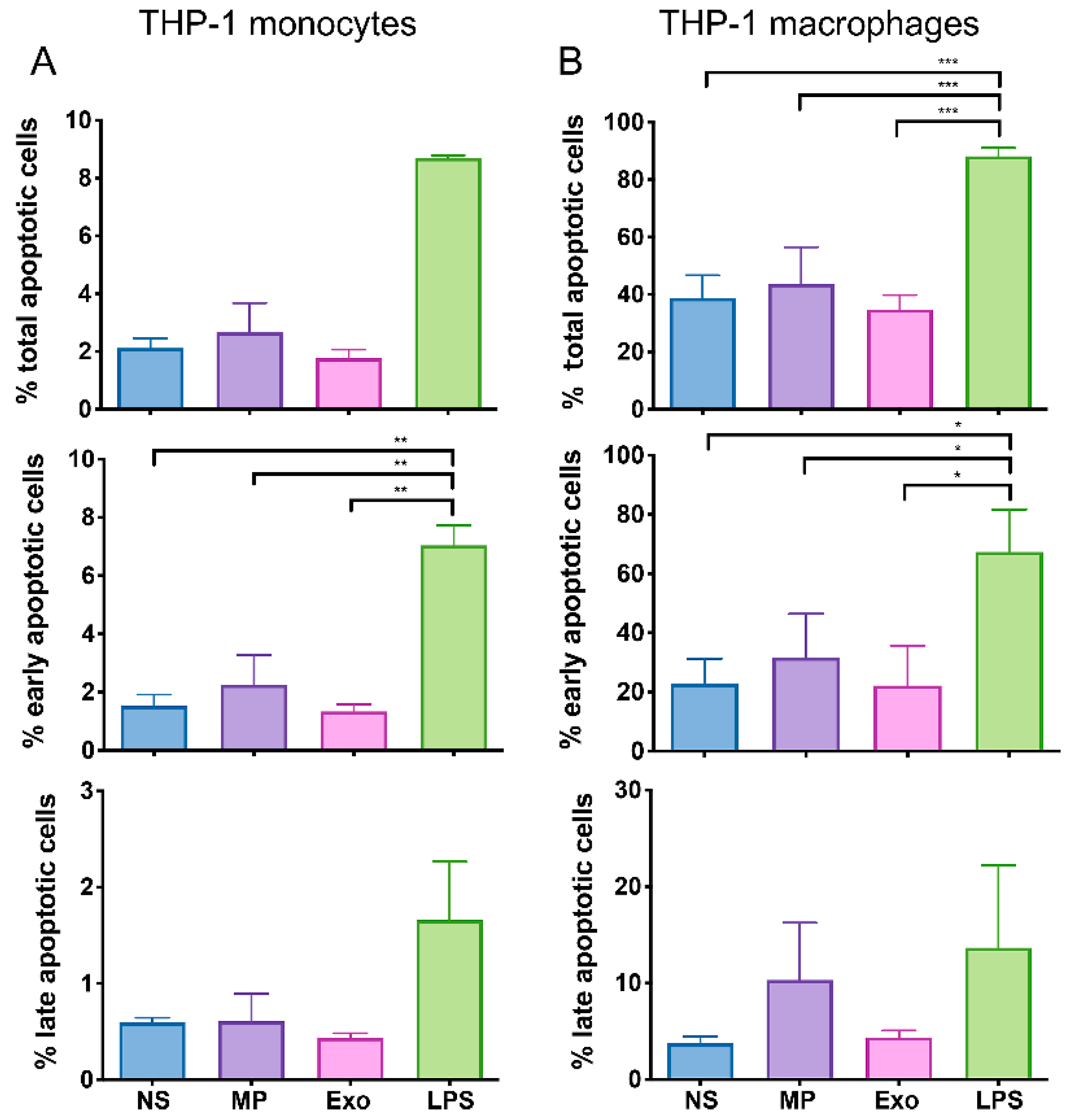
2.3. Nf-EVs Did Not Induce Apoptosis in THP-1 Monocytes and Macrophages
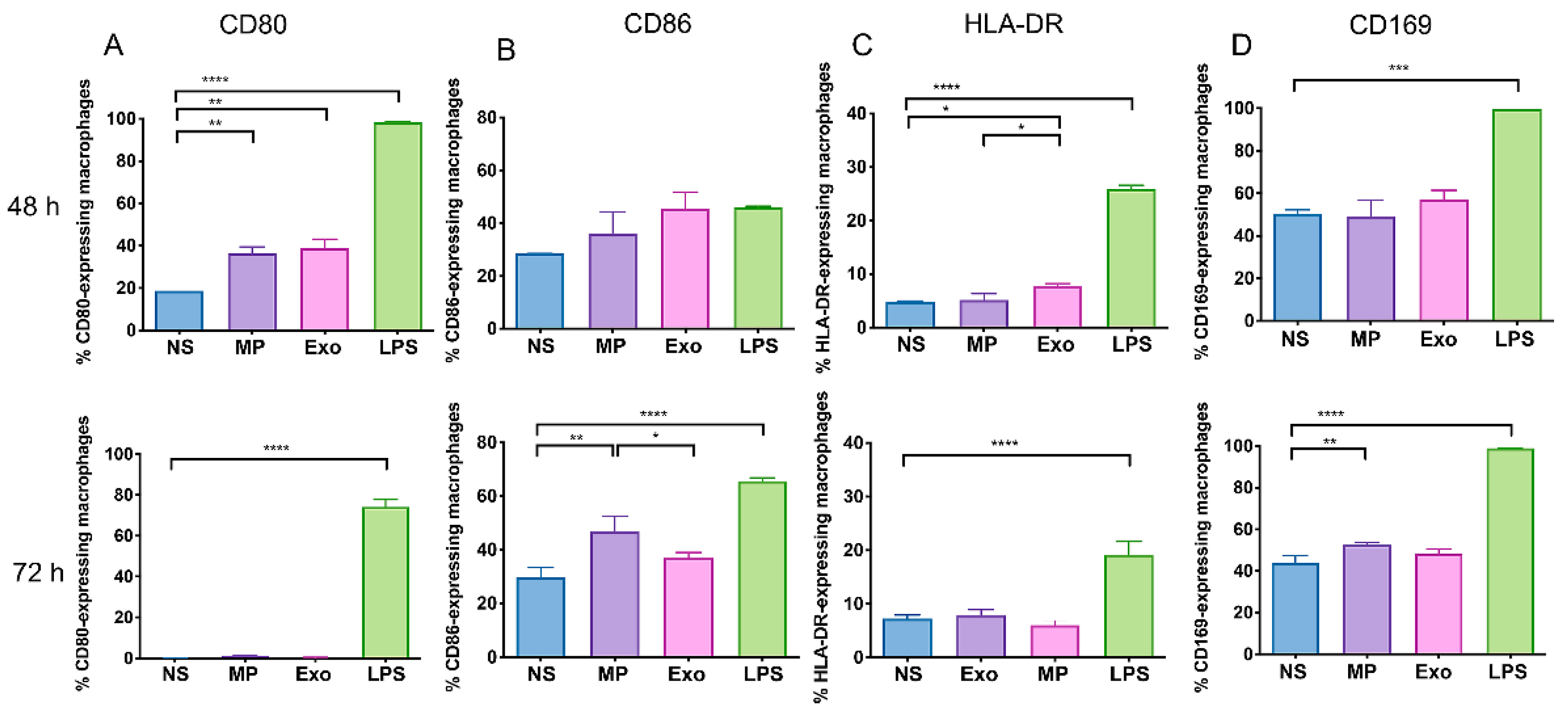
2.4. Nf-EVs Alter the Expression Markers on THP-1 Macrophages
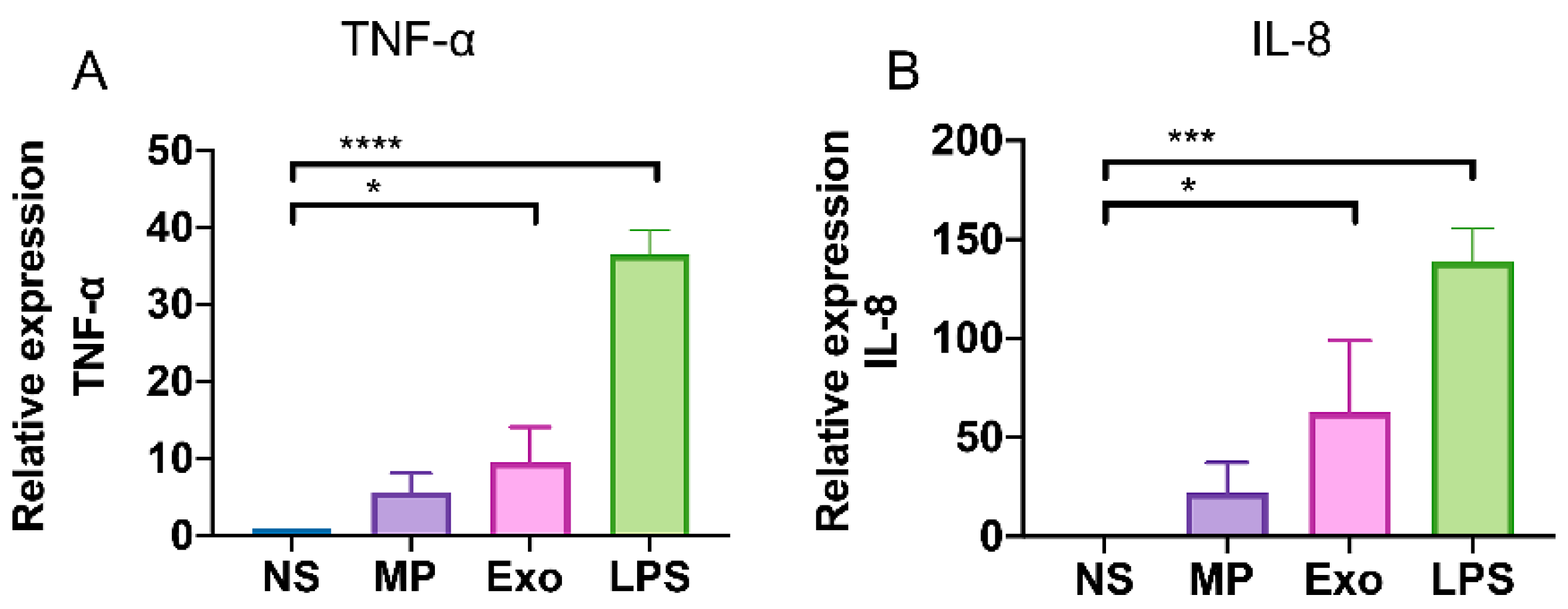
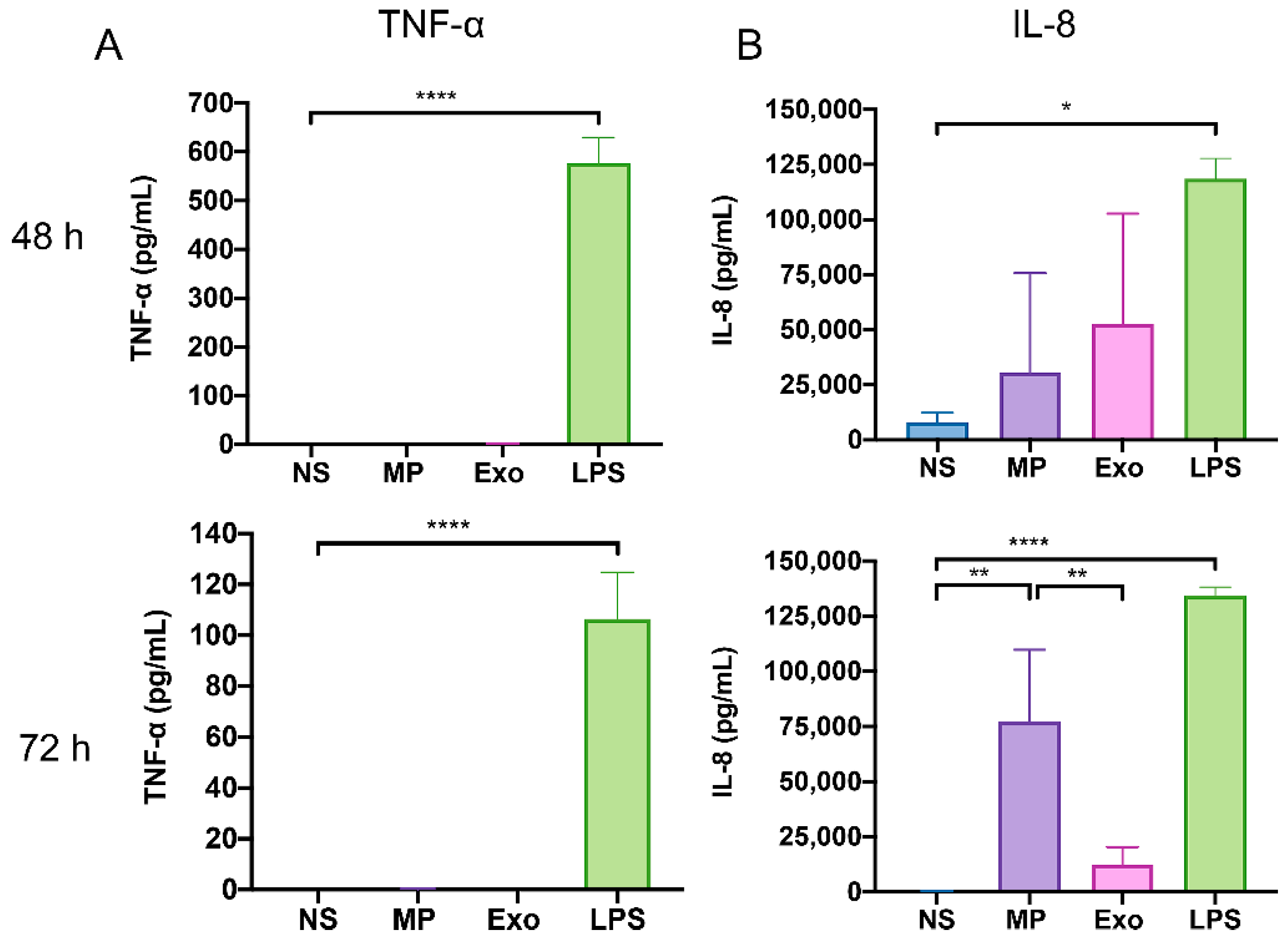
2.5. Nf-EVs Induce the Production of Pro-Inflammatory Cytokines by THP-1 Macrophages
3. Discussion
4. Materials and Methods
4.1. N. fowleri Culture
4.2. Cell Culture
4.3. Nf-EV Isolation
4.4. Transmission Electron Microscopy
4.5. Nanoparticle Tracking Analysis
4.6. Nf-EVs Labeling
4.7. Internalization of Nf-EVs with Macrophages
4.8. Cell Immunophenotyping and Apoptosis Assay by Flow Cytometry
4.9. Analysis of Cytokine Production by RT-qPCR
4.10. Measurement of Cytokine Concentration by ELISA
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butt, C.G. Primary amebic meningoencephalitis. N. Engl. J. Med. 1966, 274, 1473–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, R.F. Description of a Naegleria sp. isolated from two cases of primary amoebic meningo-encephalitis, and of the experimental pathological changes induced by it. J. Pathol. 1970, 100, 217–244. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyndall, R.L.; Ironside, K.S.; Metler, P.L.; Tan, E.L.; Hazen, T.C.; Fliermans, C.B. Effect of thermal additions on the density and distribution of thermophilic amoebae and pathogenic Naegleria fowleri in a newly created cooling lake. Appl. Environ. Microbiol. 1989, 55, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleary, S.F.; Marciano-Cabral, F. Activated macrophages demonstrate direct cytotoxicity, antibody-dependent cellular cytotoxicity, and enhanced binding of Naegleria fowleri amoebae. Cell. Immunol. 1986, 98, 125–136. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, D.; Shin, H.J. Contact-independent cell death of human microglial cells due to pathogenic Naegleria fowleri trophozoites. Korean J. Parasitol. 2008, 46, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Tiewcharoen, S.; Malainual, N.; Junnu, V.; Chetanachan, P.; Rabablert, J. Cytopathogenesis of Naegleria fowleri Thai strains for cultured human neuroblastoma cells. Parasitol. Res. 2008, 102, 997–1000. [Google Scholar] [CrossRef]
- Sohn, H.J.; Song, K.J.; Kang, H.; Ham, A.J.; Lee, J.H.; Chwae, Y.J.; Kim, K.; Park, S.; Kim, J.H.; Shin, H.J. Cellular characterization of actin gene concerned with contact-dependent mechanisms in Naegleria fowleri. Parasite Immunol. 2019, 41, e12631. [Google Scholar] [CrossRef]
- Ferrante, A.; Mocatta, T.J. Human neutrophils require activation by mononuclear leucocyte conditioned medium to kill the pathogenic free-living amoeba, Naegleria fowleri. Clin. Exp. Immunol. 1984, 56, 559–566. [Google Scholar]
- Holbrook, T.W.; Boackle, R.J.; Parker, B.W.; Vesely, J. Activation of the alternative complement pathway by Naegleria fowleri. Infect. Immun. 1980, 30, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Sandoval, I.; Serrano-Luna, J.J.; Garcia-Latorre, E.; Tsutsumi, V.; Shibayama, M. Characterization of brain inflammation during primary amoebic meningoencephalitis. Parasitol. Int. 2008, 57, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Boing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [Green Version]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, H.; Diaz-Varela, M.; Segui-Barber, J.; Roobsoong, W.; Baro, B.; Garcia-Silva, S.; Galiano, A.; Gualdron-Lopez, M.; Almeida, A.C.G.; Brito, M.A.M.; et al. Plasma-derived extracellular vesicles from Plasmodium vivax patients signal spleen fibroblasts via NF-kB facilitating parasite cytoadherence. Nat. Commun. 2020, 11, 2761. [Google Scholar] [CrossRef]
- Sampaio, N.G.; Emery, S.J.; Garnham, A.L.; Tan, Q.Y.; Sisquella, X.; Pimentel, M.A.; Jex, A.R.; Regev-Rudzki, N.; Schofield, L.; Eriksson, E.M. Extracellular vesicles from early stage Plasmodium falciparum-infected red blood cells contain PfEMP1 and induce transcriptional changes in human monocytes. Cell. Microbiol. 2018, 20, e12822. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania exosomes modulate innate and adaptive immune responses through effects on monocytes and dendritic cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, M.F.; Umezawa, E.S.; Katzin, A.M.; de Souza, W.; Alves, M.J.; Zingales, B.; Colli, W. Trypanosoma cruzi: Shedding of surface antigens as membrane vesicles. Exp. Parasitol. 1991, 72, 43–53. [Google Scholar] [CrossRef]
- Goncalves, D.S.; Ferreira, M.D.S.; Liedke, S.C.; Gomes, K.X.; de Oliveira, G.A.; Leao, P.E.L.; Cesar, G.V.; Seabra, S.H.; Cortines, J.R.; Casadevall, A.; et al. Extracellular vesicles and vesicle-free secretome of the protozoa Acanthamoeba castellanii under homeostasis and nutritional stress and their damaging potential to host cells. Virulence 2018, 9, 818–836. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.C.; Tsai, C.Y.; Huang, J.M.; Wu, S.R.; Chu, L.J.; Huang, K.Y. Quantitative proteomic analysis and functional characterization of Acanthamoeba castellanii exosome-like vesicles. Parasit. Vectors 2019, 12, 467. [Google Scholar] [CrossRef]
- Nievas, Y.R.; Coceres, V.M.; Midlej, V.; de Souza, W.; Benchimol, M.; Pereira-Neves, A.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J.; de Miguel, N. Membrane-shed vesicles from the parasite Trichomonas vaginalis: Characterization and their association with cell interaction. Cell. Mol. Life Sci. 2018, 75, 2211–2226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cao, L.; Wang, X.; Dong, J.; Zhang, N.; Li, X.; Li, J.; Zhang, X.; Gong, P. Extracellular vesicles secreted by Giardia duodenalis regulate host cell innate immunity via TLR2 and NLRP3 inflammasome signaling pathways. PLoS Negl. Trop. Dis. 2021, 15, e0009304. [Google Scholar] [CrossRef] [PubMed]
- Nicolao, M.C.; Rodriguez Rodrigues, C.; Cumino, A.C. Extracellular vesicles from Echinococcus granulosus larval stage: Isolation, characterization and uptake by dendritic cells. PLoS Negl. Trop. Dis. 2019, 13, e0007032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, A.; Cwiklinski, K.; Lalor, R.; O’Connell, B.; Robinson, M.W.; Gerlach, J.; Joshi, L.; Kilcoyne, M.; Dalton, J.P.; O’Neill, S.M. Fasciola hepatica Extracellular Vesicles isolated from excretory-secretory products using a gravity flow method modulate dendritic cell phenotype and activity. PLoS Negl. Trop. Dis. 2020, 14, e0008626. [Google Scholar] [CrossRef] [PubMed]
- Czernek, L.; Chworos, A.; Duechler, M. The Uptake of Extracellular Vesicles is Affected by the Differentiation Status of Myeloid Cells. Scand. J. Immunol. 2015, 82, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, Y.H.; Jeong, S.R.; Kim, J.H.; Song, K.J.; Kim, K.; Park, S.; Sohn, S.; Shin, H.J. Cytopathic changes and pro-inflammatory cytokines induced by Naegleria fowleri trophozoites in rat microglial cells and protective effects of an anti-Nfa1 antibody. Parasite Immunol. 2005, 27, 453–459. [Google Scholar] [CrossRef]
- Mondino, A.; Jenkins, M.K. Surface proteins involved in T cell costimulation. J. Leukoc. Biol. 1994, 55, 805–815. [Google Scholar] [CrossRef]
- Costabel, U.; Bross, K.J.; Andreesen, R.; Matthys, H. HLA-DR antigens on human macrophages from bronchoalveolar lavage fluid. Thorax 1986, 41, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; Chen, Q.; Soncin, I.; Ng, S.L.; Karjalainen, K.; Ruedl, C. A Discrete Subset of Monocyte-Derived Cells among Typical Conventional Type 2 Dendritic Cells Can Efficiently Cross-Present. Cell Rep. 2017, 21, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Kikuchi, K.; Tanaka, M. CD169 macrophages regulate immune responses toward particulate materials in the circulating fluid. J. Biochem. 2018, 164, 77–85. [Google Scholar] [CrossRef]
- Giovane, R.A.; Lavender, P.D. Central Nervous System Infections. Prim. Care 2018, 45, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; Grau, G.E.; Riley, E.M.; de Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atayde, V.D.; Hassani, K.; da Silva Lira Filho, A.; Borges, A.R.; Adhikari, A.; Martel, C.; Olivier, M. Leishmania exosomes and other virulence factors: Impact on innate immune response and macrophage functions. Cell. Immunol. 2016, 309, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Xiu, F.; Wang, J.; Cong, H.; He, S.; Shi, Y.; Wang, X.; Li, X.; Zhou, H. Characterization of exosomes derived from Toxoplasma gondii and their functions in modulating immune responses. Int. J. Nanomed. 2018, 13, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, Z.; Shen, J.; Liu, Z.; Liang, J.; Wu, X.; Sun, X.; Wu, Z. Exosome-like vesicles derived by Schistosoma japonicum adult worms mediates M1 type immune- activity of macrophage. Parasitol. Res. 2015, 114, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Martin-Jaular, L.; Nakayasu, E.S.; Ferrer, M.; Almeida, I.C.; Del Portillo, H.A. Exosomes from Plasmodium yoelii-infected reticulocytes protect mice from lethal infections. PLoS ONE 2011, 6, e26588. [Google Scholar] [CrossRef] [Green Version]
- Silverman, J.M.; Reiner, N.E. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Front. Cell. Infect. Microbiol. 2011, 1, 26. [Google Scholar] [CrossRef] [Green Version]
- Twu, O.; de Miguel, N.; Lustig, G.; Stevens, G.C.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J. Trichomonas vaginalis exosomes deliver cargo to host cells and mediate hostratioparasite interactions. PLoS Pathog. 2013, 9, e1003482. [Google Scholar] [CrossRef] [Green Version]
- Combes, V.; Taylor, T.E.; Juhan-Vague, I.; Mege, J.L.; Mwenechanya, J.; Tembo, M.; Grau, G.E.; Molyneux, M.E. Circulating endothelial microparticles in malawian children with severe falciparum malaria complicated with coma. JAMA 2004, 291, 2542–2544. [Google Scholar] [CrossRef]
- Burri, D.C.; Gottstein, B.; Zumkehr, B.; Hemphill, A.; Schurch, N.; Wittwer, M.; Muller, N. Development of a high- versus low-pathogenicity model of the free-living amoeba Naegleria fowleri. Microbiology 2012, 158, 2652–2660. [Google Scholar] [CrossRef] [Green Version]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell. Biol. 2006, 3, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Balaj, L.; Alian, S.; Mantel, P.Y.; Halleck, A.E.; Trachtenberg, A.J.; Soria, C.E.; Oquin, S.; Bonebreak, C.M.; Saracoglu, E.; et al. Current methods for the isolation of extracellular vesicles. Biol. Chem. 2013, 394, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Zhao, W.L.; Ye, Y.Y.; Bai, X.C.; Liu, R.Q.; Chang, L.F.; Zhou, Q.; Sui, S.F. Cellular internalization of exosomes occurs through phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Park, C.E.; Kim, J.H.; Sohn, H.J.; Lee, J.; Jung, S.Y.; Shin, H.J. Naegleria fowleri lysate induces strong cytopathic effects and pro-inflammatory cytokine release in rat microglial cells. Korean J. Parasitol. 2011, 49, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Cho, M.S.; Jung, S.U.; Kim, H.I.; Park, S.; Kim, H.J.; Im, K.I. Molecular cloning and characterization of a gene encoding a 13.1 kDa antigenic protein of Naegleria fowleri. J. Eukaryot. Microbiol. 2001, 48, 713–717. [Google Scholar] [CrossRef]
- Jung, S.Y.; Kim, J.H.; Song, K.J.; Lee, Y.J.; Kwon, M.H.; Kim, K.; Park, S.; Im, K.I.; Shin, H.J. Gene silencing of nfa1 affects the in vitro cytotoxicity of Naegleria fowleri in murine macrophages. Mol. Biochem. Parasitol. 2009, 165, 87–93. [Google Scholar] [CrossRef]
- Chávez-Galán, L.; Olleros, M.L.; Vesin, D.; Garcia, I. Much More than M1 and M2 Macrophages, There are also CD169+ and TCR+ Macrophages. Front. Immunol. 2015, 6, 263. [Google Scholar]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Wang, J.; Yao, Y.; Xiong, J.; Wu, J.; Tang, X.; Li, G. Evaluation of the inflammatory response in macrophages stimulated with exosomes secreted by Mycobacterium avium-infected macrophages. Biomed. Res. Int. 2015, 2015, 658421. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, L.; Al-Enezi, F.; Al-Saif, M.; Warsy, A.; Khabar, K.S.; Hitti, E.G. Sustained stabilization of Interleukin-8 mRNA in human macrophages. RNA Biol. 2014, 11, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kang, J.M.; Kim, T.I.; Kim, J.H.; Sohn, H.J.; Na, B.K.; Shin, H.J. Excretory and Secretory Proteins of Naegleria fowleri Induce Inflammatory Responses in BV-2 Microglial Cells. J. Eukaryot. Microbiol. 2017, 64, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Tiewcharoen, S.; Junnu, V. Distribution of pathogenic Naegleria spp in Thailand. Southeast Asian J. Trop. Med. Public Health 2001, 32 (Suppl. S2), 172–178. [Google Scholar] [PubMed]
- Atipimonpat, A.; Siwaponanan, P.; Khuhapinant, A.; Svasti, S.; Sukapirom, K.; Khowawisetsut, L.; Pattanapanyasat, K. Extracellular vesicles from thalassemia patients carry iron-containing ferritin and hemichrome that promote cardiac cell proliferation. Ann. Hematol. 2021, 100, 1929–1946. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Accession Number | |
|---|---|---|---|
| TNF-α | Forward Reverse | CCTGTAGCCCATGTTGTAGCAAA TGAGGAGCACATGGGTGGAG | NM_000594 |
| IL-1α | Forward Reverse | GCGTTTGAGTCAGCAAAGAAGT CATGGAGTGGGCCATAGCTT | NM_000575 |
| IL-6 | Forward Reverse | AACCTGAACCTTCCAAAGATGG TCTGGCTTGTTCCTCACTACT | NM_001371096 |
| IL-8 | Forward Reverse | ACTGAGAGTGATTGAGAGTGGAC AACCCTCTGCACCCAGTTTTC | NM_001354840 |
| IL-10 | Forward Reverse | CCAAGCTGAGAACCAAGACC GGGAAGAAATCGATGACAGC | NM_000572 |
| CXCL10 | Forward Reverse | AAGTGGCATTCAAGGAGTACCT GGACAAATTGGCTTGCAGGA | NM_001565 |
| GAPDH | Forward Reverse | ACCCAGAAGACTGTGGATGG TTCAGCTCAGGGATGACCTT | NM_002046 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lertjuthaporn, S.; Somkird, J.; Lekmanee, K.; Atipimonpat, A.; Sukapirom, K.; Sawasdipokin, H.; Tiewcharoen, S.; Pattanapanyasat, K.; Khowawisetsut, L. Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage. Pathogens 2022, 11, 632. https://doi.org/10.3390/pathogens11060632
Lertjuthaporn S, Somkird J, Lekmanee K, Atipimonpat A, Sukapirom K, Sawasdipokin H, Tiewcharoen S, Pattanapanyasat K, Khowawisetsut L. Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage. Pathogens. 2022; 11(6):632. https://doi.org/10.3390/pathogens11060632
Chicago/Turabian StyleLertjuthaporn, Sakaorat, Jinjuta Somkird, Kittima Lekmanee, Anyapat Atipimonpat, Kasama Sukapirom, Hathai Sawasdipokin, Supathra Tiewcharoen, Kovit Pattanapanyasat, and Ladawan Khowawisetsut. 2022. "Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage" Pathogens 11, no. 6: 632. https://doi.org/10.3390/pathogens11060632
APA StyleLertjuthaporn, S., Somkird, J., Lekmanee, K., Atipimonpat, A., Sukapirom, K., Sawasdipokin, H., Tiewcharoen, S., Pattanapanyasat, K., & Khowawisetsut, L. (2022). Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage. Pathogens, 11(6), 632. https://doi.org/10.3390/pathogens11060632