
Prospective Longitudinal Study of Putative Agents Involved in Complex Gill Disorder in Atlantic salmon (Salmo salar)

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
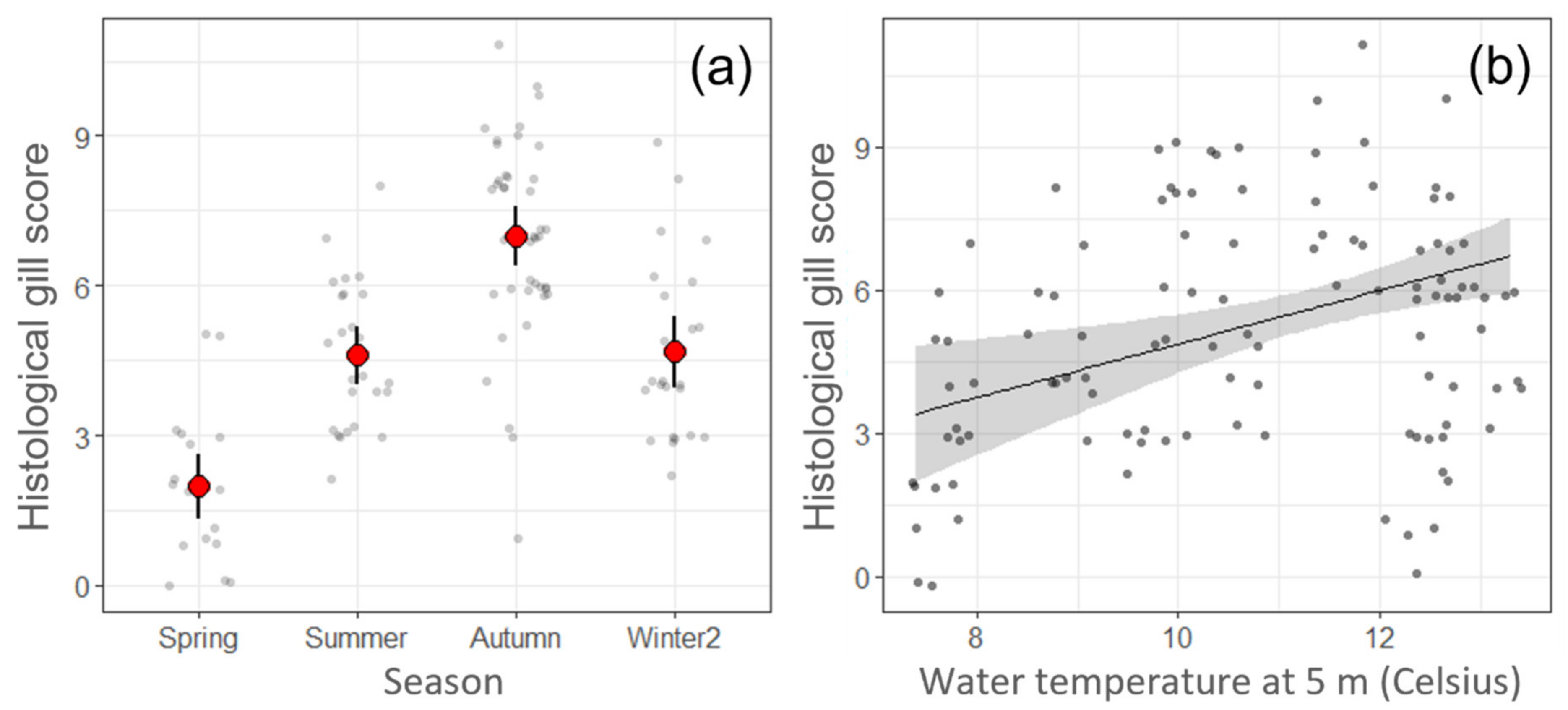
2.1. Environmental Data
2.2. Descriptive Epidemiology
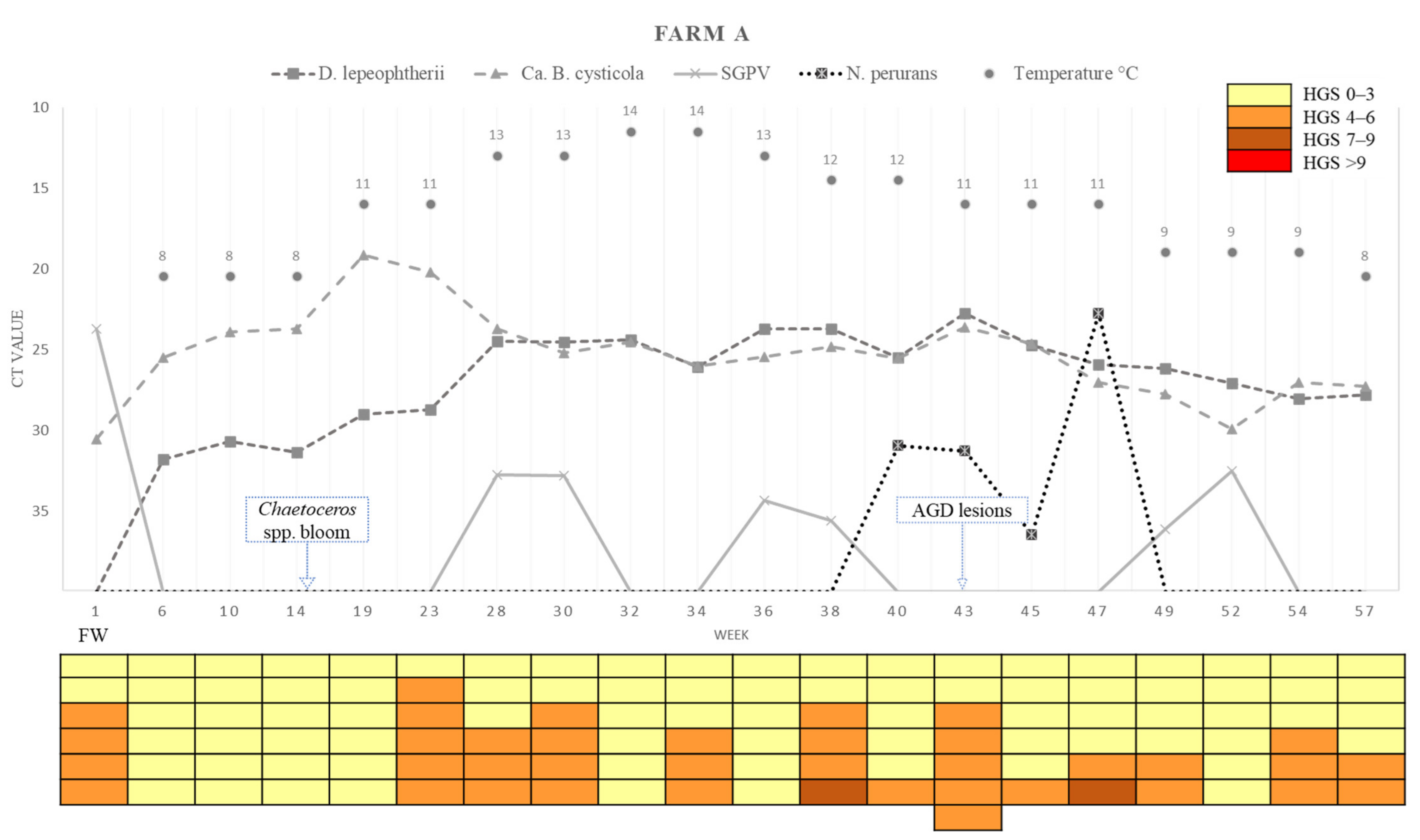
2.2.1. Farm A
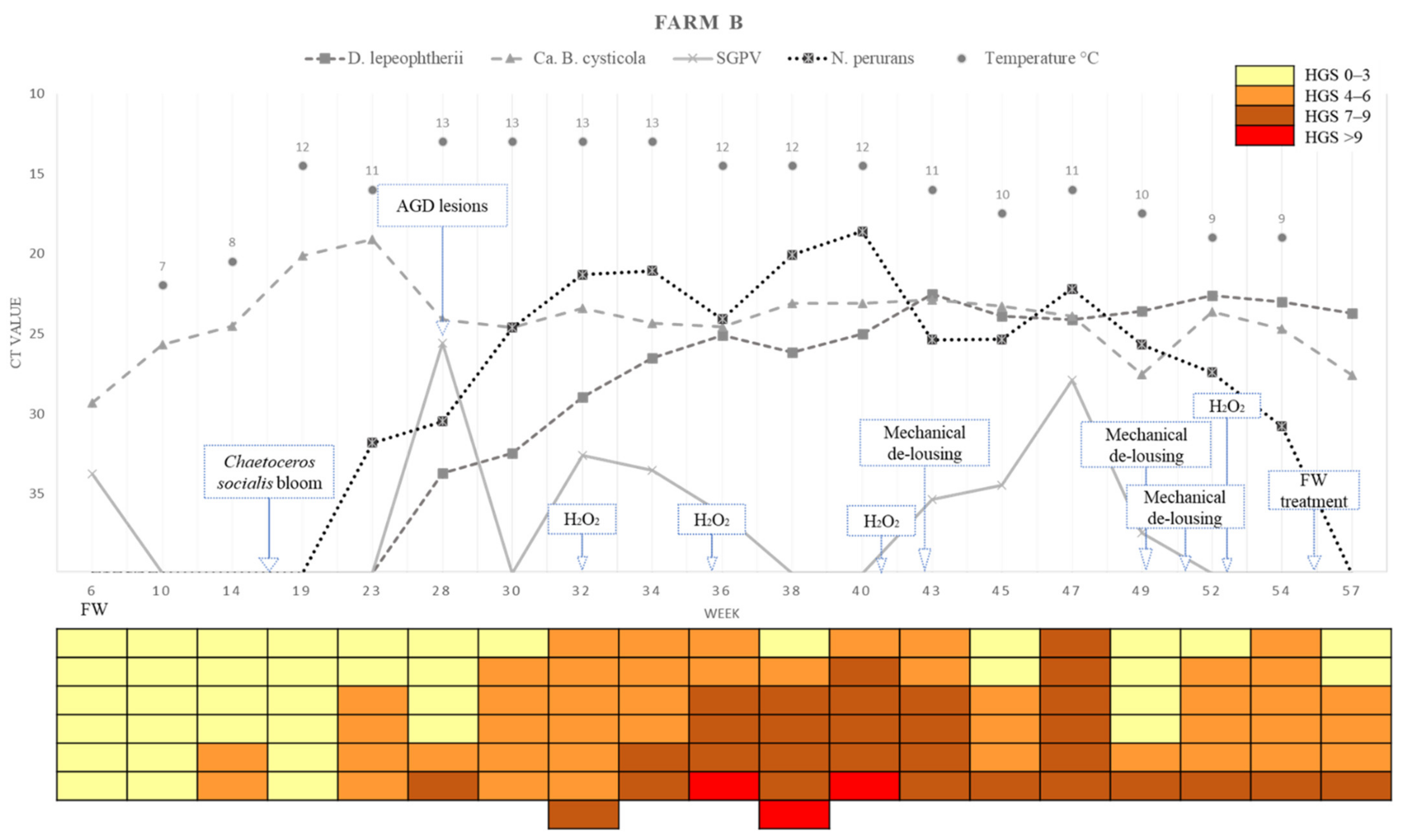
2.2.2. Farm B
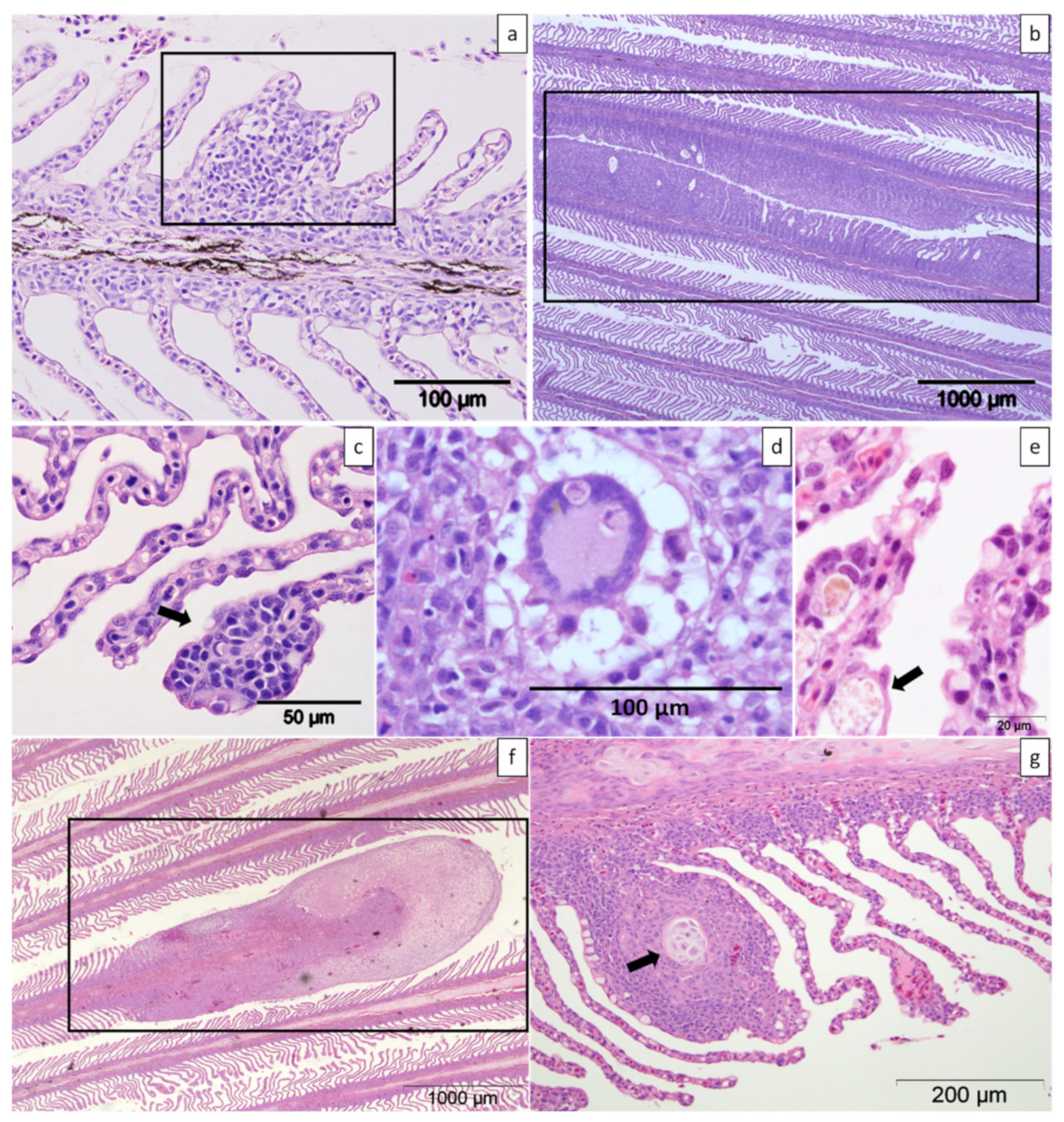
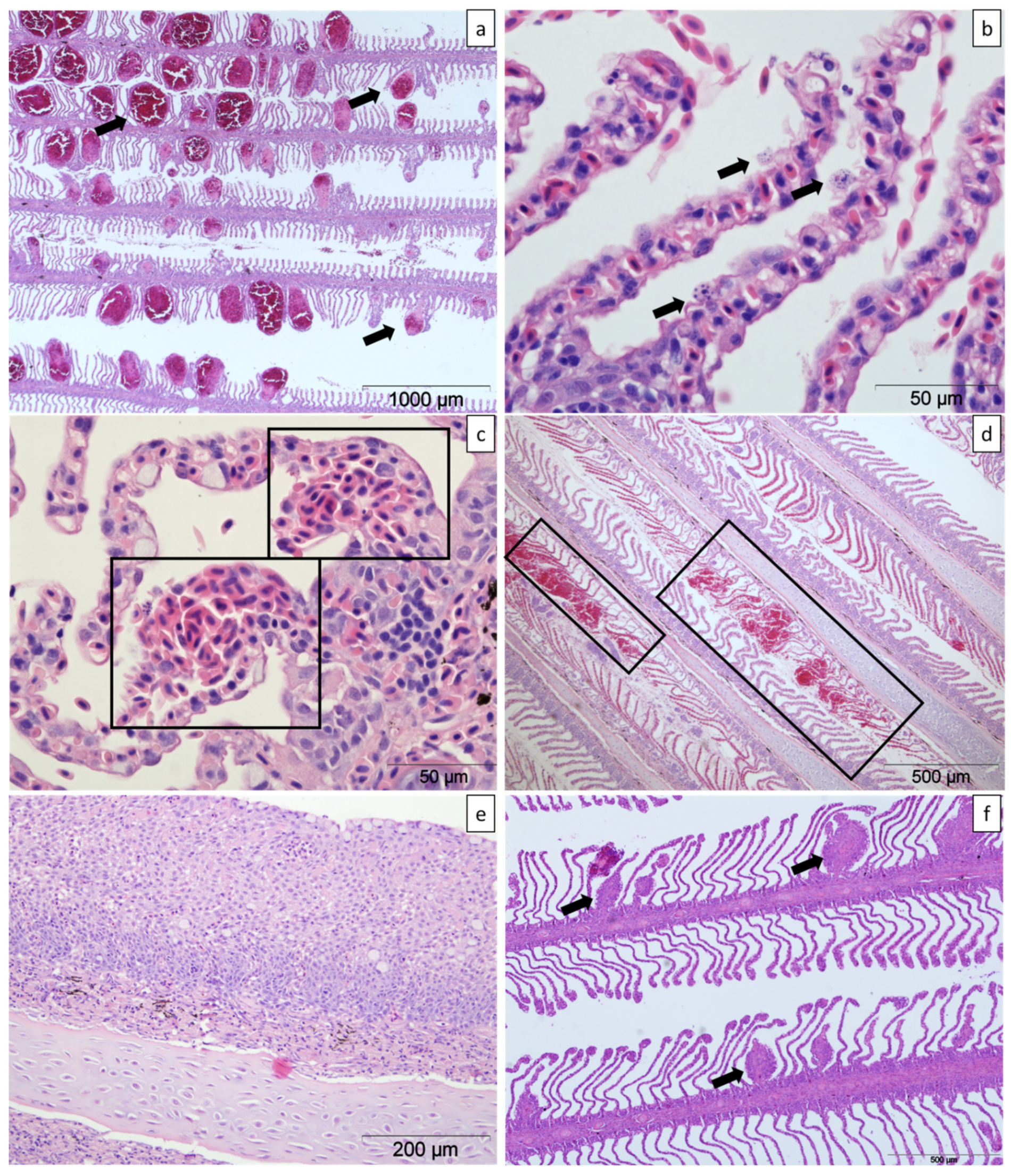
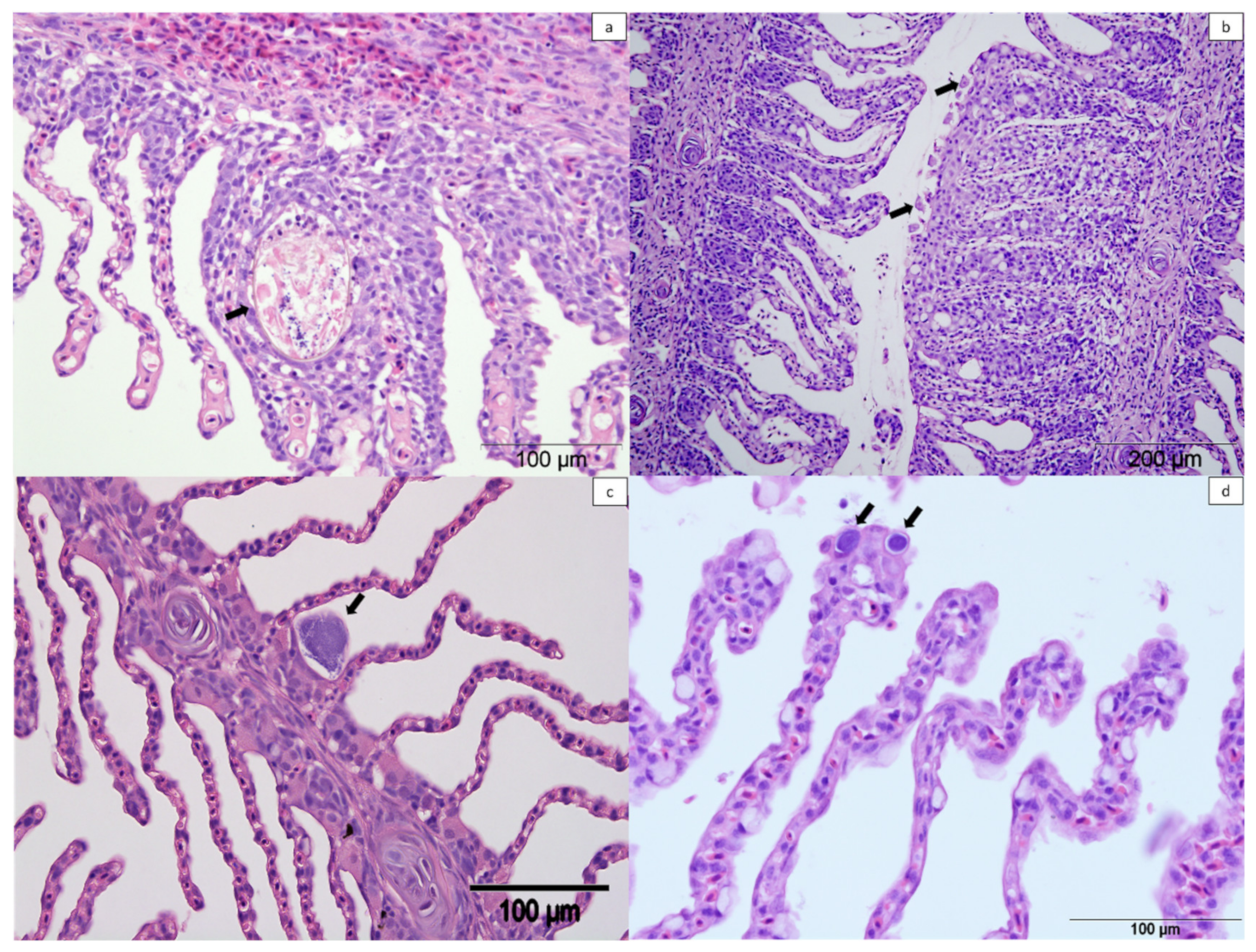
2.3. Histology
2.3.1. Farm A
2.3.2. Farm B
2.4. Changes in the Levels of the Different Pathogens across Time
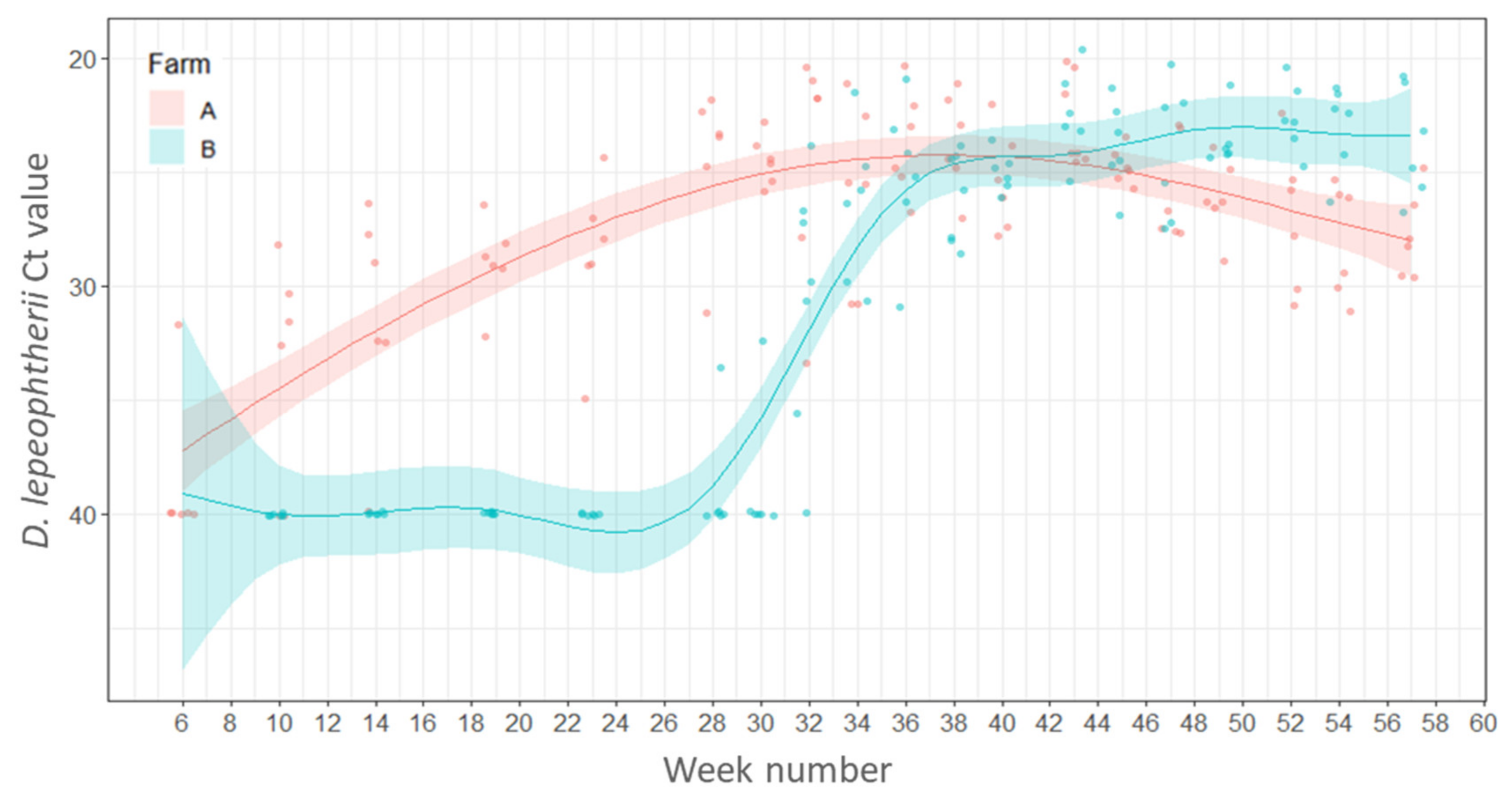
2.4.1. Variations in Load (Ct Values) of Desmozoon lepeophtherii
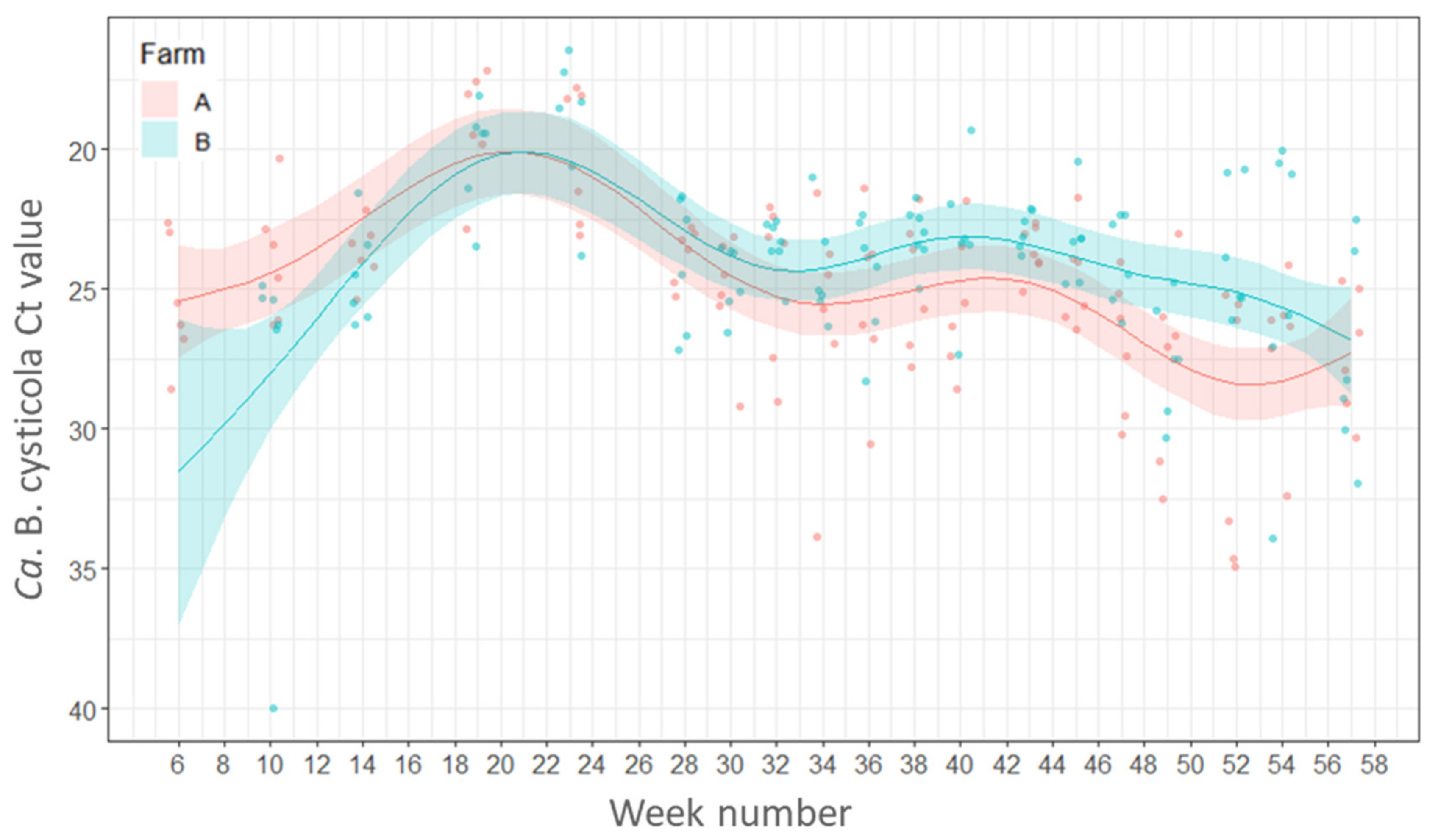
2.4.2. Variations in Load (Ct Values) of Candidatus Branchiomonas cysticola
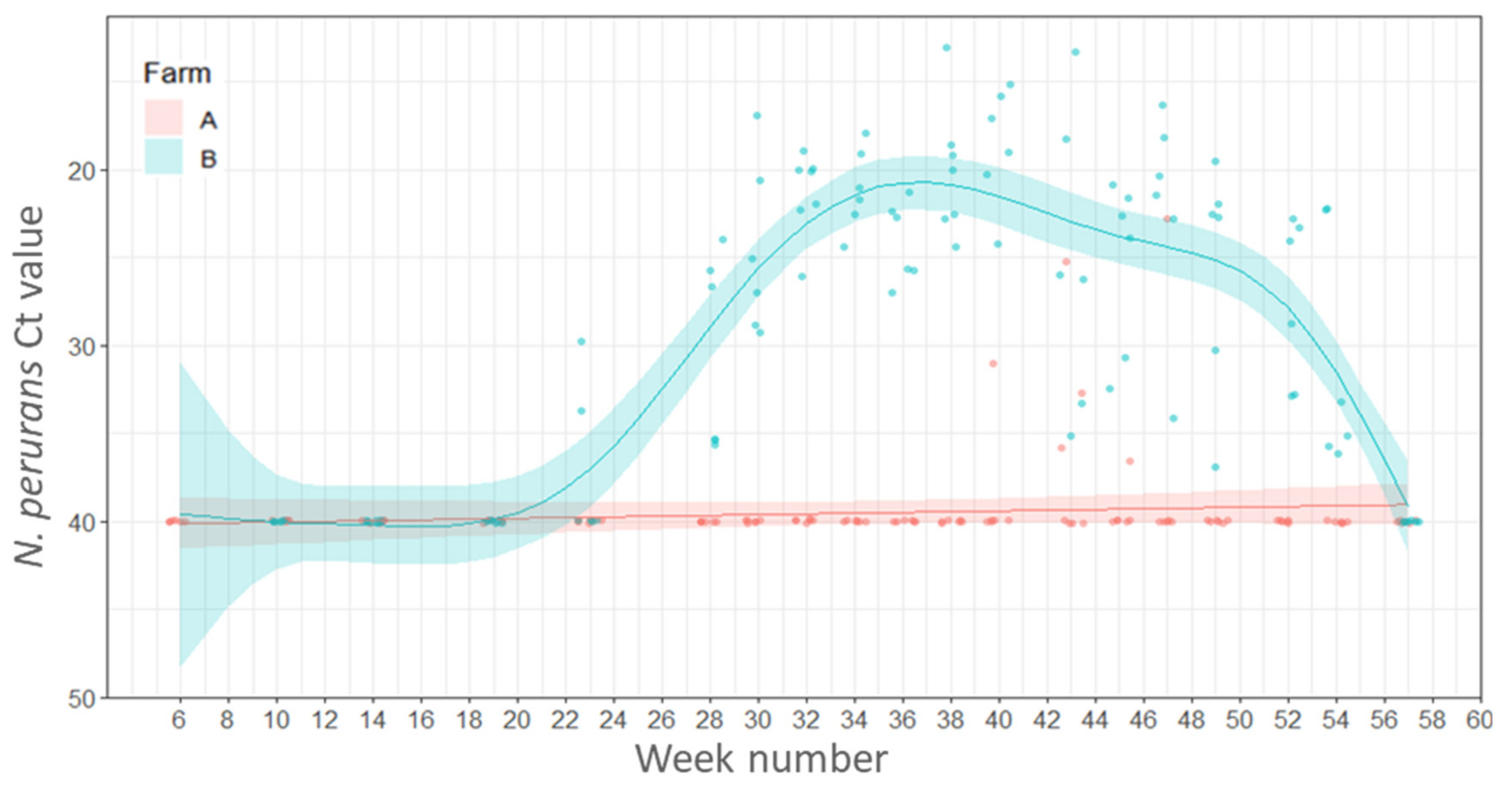
2.4.3. Variations in Load (Ct Values) of N. perurans
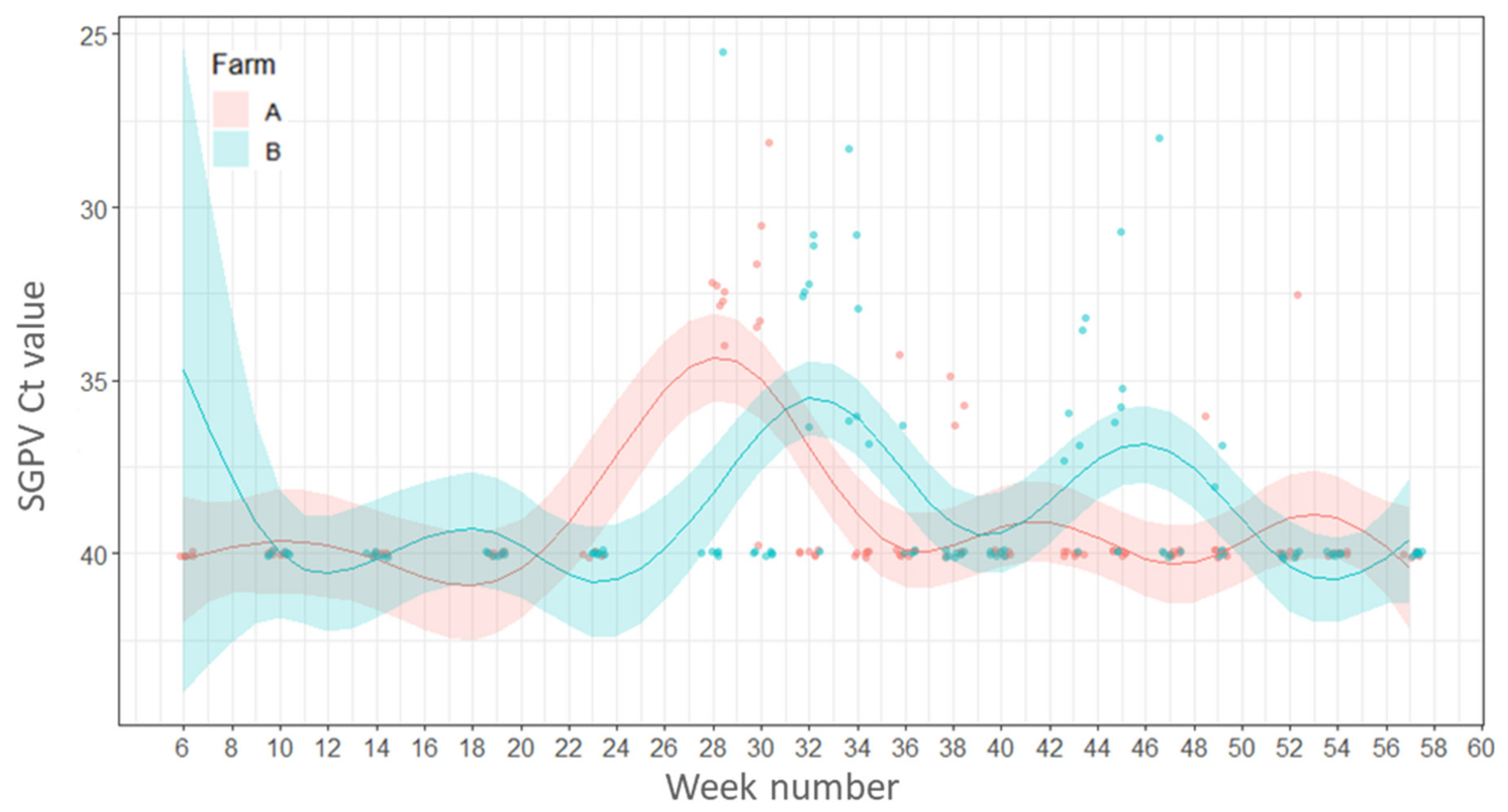
2.4.4. Variations in Load (Ct Values) of Salmon Gill Poxvirus (SGPV)
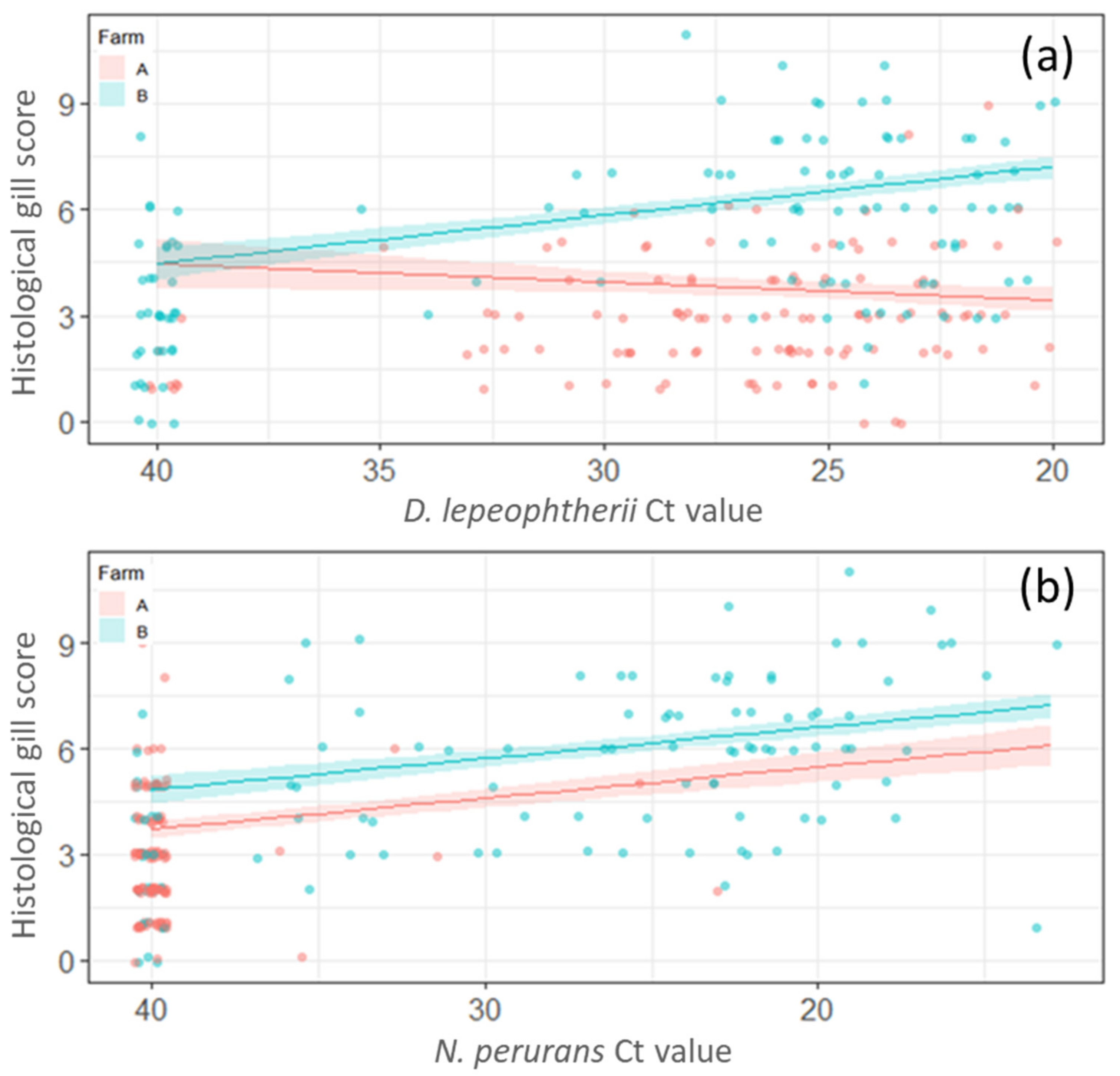
2.5. Linear Regression Models of the Histological Gill Score
2.6. Summary of the Variation in Pathogen Loads (Ct Values), Epidemiology in the Farms, HGS and Temperatures
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Sample and Data Collection from Farms
4.3. Histopathology
4.4. Molecular Analyses
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boerlage, A.S.; Ashby, A.; Herrero, A.; Reeves, A.; Gunn, G.J.; Rodger, H.D. Epidemiology of marine gill diseases in Atlantic salmon (Salmo salar) aquaculture: A review. Rev. Aquac. 2020, 12, 2140–2159. [Google Scholar] [CrossRef]
- Sommerset, I.; Bang Jensen, B.; Bornø, B.; Haukaas, A.; Brun, E. The Health Situation in Norwegian Aquaculture 2020. Available online: https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2021/fish-health-report-2020 (accessed on 9 July 2022).
- Shinn, A.P.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.E.; Brooker, A.J. Economic costs of protistan and metazoan parasites to global mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.O.; Rodger, H.D. A review of infectious gill disease in marine salmonid fish. J. Fish Dis. 2011, 34, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Rodger, H.D. Amoebic gill disease (AGD) in farmed salmon (Salmo salar) in Europe. Fish Vet. J. 2014, 14, 16–27. [Google Scholar]
- Herrero, A.; Thompson, K.D.; Ashby, A.; Rodger, H.D.; Dagleish, M.P. Complex gill disease: An emerging syndrome in farmed Atlantic salmon (Salmo Salar L.). J. Comp. Pathol. 2018, 163, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Noguera, P.; Olsen, A.B.; Hoare, J.; Lie, K.I.; Marcos-López, M.; Poppe, T.T.; Rodger, H. Complex gill disorder (CGD): A histopathology workshop report. Bull. Eur. Assoc. Fish Pathol. 2019, 39, 172–176. [Google Scholar]
- Gjessing, M.C.; Spilsberg, B.; Steinum, T.M.; Amundsen, M.; Austbø, L.; Hansen, H.; Colquhoun, D.; Olsen, A.B. Multi-agent in situ hybridization confirms Ca. Branchiomonas cysticola as a major contributor in complex gill disease in Atlantic salmon. Fish Shellfish Immunol. Rep. 2021, 2, 100026. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Steinum, T.; Olsen, A.B.; Lie, K.I.; Tavornpanich, S.; Colquhoun, D.J.; Gjevre, A.-G. Histopathological investigation of complex gill disease in sea farmed Atlantic salmon. PLoS ONE 2019, 14, e0222926. [Google Scholar] [CrossRef] [PubMed]
- Gjessing, M.C.; Thoen, E.; Tengs, T.; Skotheim, S.A.; Dale, O.B. Salmon gill poxvirus, a recently characterized infectious agent of multifactorial gill disease in freshwater- and seawater-reared Atlantic salmon. J. Fish Dis. 2017, 40, 1253–1265. [Google Scholar] [CrossRef]
- Mitchell, S.O.; Steinum, T.M.; Toenshoff, E.R.; Kvellestad, A.; Falk, K.; Horn, M.; Colquhoun, D.J. “Candidatus Branchiomonas cysticola” is a common agent of epitheliocysts in seawater-farmed Atlantic salmon Salmo salar in Norway and Ireland. Dis. Aquat. Organ. 2013, 103, 35–43. [Google Scholar] [CrossRef]
- Weli, S.C.; Dale, O.B.; Hansen, H.; Gjessing, M.C.; Rønneberg, L.B.; Falk, K. A case study of Desmozoon lepeophtherii infection in farmed Atlantic salmon associated with gill disease, peritonitis, intestinal infection, stunted growth, and increased mortality. Parasit. Vectors 2017, 10, 370. [Google Scholar] [CrossRef] [PubMed]
- Steinum, T.; Kvellestad, A.; Colquhoun, D.J.; Heum, M.; Mohammad, S.; Grøntvedt, R.N.; Falk, K. Microbial and pathological findings in farmed Atlantic salmon Salmo salar with proliferative gill inflammation. Dis. Aquat. Organ. 2010, 91, 201–211. [Google Scholar] [CrossRef]
- Gunnarsson, G.S.; Blindheim, S.; Karlsbakk, E.; Plarre, H.; Imsland, A.K.; Handeland, S.; Sveier, H.; Nylund, A. Desmozoon lepeophtherii (microsporidian) infections and pancreas disease (PD) outbreaks in farmed Atlantic salmon (Salmo salar L.). Aquaculture 2017, 468, 141–148. [Google Scholar] [CrossRef]
- Thoen, E.; Tartor, H.; Amundsen, M.; Dale, O.B.; Sveinsson, K.; Rønning, H.P.; Grønneberg, E.; Dahle, M.K.; Gjessing, M.C. First record of experimentally induced salmon gill poxvirus disease (SGPVD) in Atlantic salmon (Salmo salar L.). Vet. Res. 2020, 51, 63. [Google Scholar] [CrossRef] [PubMed]
- Nylund, A.; Watanabe, K.; Nylund, S.; Karlsen, M.; Sæther, P.A.; Arnesen, C.E.; Karlsbakk, E. Morphogenesis of salmonid gill poxvirus associated with proliferative gill disease in farmed Atlantic salmon (Salmo salar) in Norway. Arch. Virol. 2008, 153, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.B.; Nowak, B.F. Amoebic gill disease: Sequential pathology in cultured Atlantic salmon, Salmo salar L. J. Fish Dis. 2003, 26, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, M.; Nylund, A.; Watanabe, K.; Helvik, J.V.; Nylund, S.; Plarre, H. Characterization of ‘Candidatus Clavochlamydia salmonicola’: An intracellular bacterium infecting salmonid fish. Environ. Microbiol. 2008, 10, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Toenshoff, E.R.; Kvellestad, A.; Mitchell, S.O.; Steinum, T.; Falk, K.; Colquhoun, D.J.; Horn, M. A novel betaproteobacterial agent of gill epitheliocystis in seawater farmed Atlantic salmon (Salmo salar). PLoS ONE 2012, 7, e32696. [Google Scholar] [CrossRef]
- Downes, J.K.; Yatabe, T.; Marcos-Lopez, M.; Rodger, H.D.; MacCarthy, E.; O’Connor, I.; Collins, E.; Ruane, N.M. Investigation of co-infections with pathogens associated with gill disease in Atlantic salmon during an amoebic gill disease outbreak. J. Fish Dis. 2018, 41, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, G.S.; Karlsbakk, E.; Blindheim, S.; Plarre, H.; Imsland, A.K.; Handeland, S.; Sveier, H.; Nylund, A. Temporal changes in infections with some pathogens associated with gill disease in farmed Atlantic salmon (Salmo salar L). Aquaculture 2017, 468, 126–134. [Google Scholar] [CrossRef]
- Laurin, E.; Jaramillo, D.; Vanderstichel, R.; Ferguson, H.; Kaukinen, K.H.; Schulze, A.D.; Keith, I.R.; Gardner, I.A.; Miller, K.M. Histopathological and novel high-throughput molecular monitoring data from farmed salmon (Salmo salar and Oncorhynchus spp.) in British Columbia, Canada, from 2011–2013. Aquaculture 2019, 499, 220–234. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Christensen, D.H.; Manji, F.; Mohammad, S.; Petersen, P.E.; Saure, B.; Skjengen, C.; Weli, S.C.; Dale, O.B. Salmon gill poxvirus disease in Atlantic salmon fry as recognized by improved immunohistochemistry also demonstrates infected cells in non-respiratory epithelial cells. J. Fish Dis. 2018, 41, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Wiik-Nielsen, J.; Gjessing, M.; Solheim, H.T.; Litlabø, A.; Gjevre, A.-G.; Kristoffersen, A.B.; Powell, M.D.; Colquhoun, D.J. Ca. Branchiomonas cysticola, Ca. Piscichlamydia salmonis and salmon gill pox virus transmit horizontally in Atlantic salmon held in fresh water. J. Fish Dis. 2017, 40, 1387–1394. [Google Scholar] [CrossRef]
- van West, P. Saprolegnia parasitica, an oomycete pathogen with a fishy appetite: New challenges for an old problem. Mycologist 2006, 20, 99–104. [Google Scholar] [CrossRef]
- Bustos, P.A.; Young, N.D.; Rozas, M.A.; Bohle, H.M.; Ildefonso, R.S.; Morrison, R.N.; Nowak, B.F. Amoebic gill disease (AGD) in Atlantic salmon (Salmo salar) farmed in Chile. Aquaculture 2011, 310, 281–288. [Google Scholar] [CrossRef]
- Matthews, C.G.G.; Richards, R.H.; Shinn, A.P.; Cox, D.I. Gill pathology in Scottish farmed Atlantic salmon, Salmo salar L., associated with the microsporidian Desmozoon lepeophtherii Freeman et Sommerville, 2009. J. Fish Dis. 2013, 36, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Steinum, T.; Kvellestad, A.; Rønneberg, L.B.; Nilsen, H.; Asheim, A.; Fjell, K.; Nygård, S.M.R.; Olsen, A.B.; Dale, O.B. First cases of amoebic gill disease (AGD) in Norwegian seawater farmed Atlantic salmon, Salmo salar L., and phylogeny of the causative amoeba using 18S cDNA Sequences. J. Fish Dis. 2008, 31, 205–214. [Google Scholar] [CrossRef]
- Rodger, H.D.; Murphy, K.; Mitchell, S.O.; Henry, L. Gill disease in marine farmed Atlantic salmon at four farms in Ireland. Vet. Rec. 2011, 168, 668. [Google Scholar] [CrossRef]
- Benedicenti, O.; Pottinger, T.G.; Collins, C.; Secombes, C.J. Effects of temperature on amoebic gill disease development: Does it play a role? J. Fish Dis. 2019, 42, 1241–1258. [Google Scholar] [CrossRef]
- Clinton, M.; Ferrier, D.E.K.; Martin, S.A.M.; Brierley, A.S. Impacts of jellyfish on marine cage aquaculture: An overview of existing knowledge and the challenges to finfish health. ICES J. Mar. Sci. 2021, 78, 1557–1573. [Google Scholar] [CrossRef]
- Rodger, H.D.; Henry, L.; Mitchell, S.O. Non-infectious gill disorders of marine salmonid fish. Rev. Fish Biol. Fish. 2011, 21, 423–440. [Google Scholar] [CrossRef]
- Malviya, S.; Scalco, E.; Audic, S.; Vincent, F.; Veluchamy, A.; Poulain, J.; Wincker, P.; Iudicone, D.; de Vargas, C.; Bittner, L.; et al. Insights into global diatom distribution and diversity in the world’s ocean. Proc. Natl. Acad. Sci. USA 2016, 113, E1516–E1525. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.W.; Dear, G.; Seaton, D.D. Mortality associated with phytoplankton blooms among farmed Atlantic salmon, Salmo salar L., in Scotland. Aquaculture 1989, 78, 217–222. [Google Scholar] [CrossRef]
- Burridge, L.E.; Martin, J.L.; Lyons, M.C.; LeGresley, M.M. Lethality of microalgae to farmed Atlantic salmon (Salmo salar). Aquaculture 2010, 308, 101–105. [Google Scholar] [CrossRef]
- Sveen, S.; Øverland, H.; Karlsbakk, E.; Nylund, A. Paranucleospora theridion (microsporidia) infection dynamics in farmed Atlantic salmon Salmo salar put to sea in spring and autumn. Dis. Aquat. Organ. 2012, 101, 43–49. [Google Scholar] [CrossRef]
- Herrero, A.; Palenzuela, O.; Rodger, H.; Matthews, C.; Marcos-López, M.; Bron, J.E.; Dagleish, M.P.; Thompson, K.D. Novel DNA-based in situ hybridization method to detect Desmozoon lepeophtherii in Atlantic salmon tissues. J. Fish Dis. 2022, 45, 871–882. [Google Scholar] [CrossRef]
- Mitchell, S.O.; Steinum, T.; Rodger, H.; Holland, C.; Falk, K.; Colquhoun, D.J. Epitheliocystis in Atlantic salmon, Salmo salar L., farmed in fresh water in Ireland is associated with “Candidatus Clavochlamydia salmonicola” infection. J. Fish Dis. 2010, 33, 665–673. [Google Scholar] [CrossRef]
- Baxter, E.J.; Sturt, M.M.; Ruane, N.M.; Doyle, T.K.; McAllen, R.; Rodger, H.D. Biofouling of the hydroid Ectopleura larynx on aquaculture nets in Ireland: Implications for finfish health. Fish Veterinary J. 2012, 13, 18–30. [Google Scholar]
- Østevik, L.; Stormoen, M.; Nødtvedt, A.; Alarcón, M.; Lie, K.-I.; Skagøy, A.; Rodger, H. Assessment of acute effects of in situ net cleaning on gill health of farmed Atlantic salmon (Salmo salar L). Aquaculture 2021, 545, 737203. [Google Scholar] [CrossRef]
- Østevik, L.; Stormoen, M.; Evensen, Ø.; Xu, C.; Lie, K.I.; Nødtvedt, A.; Rodger, H.; Skagøy, A.; Manji, F.; Alarcón, M. Effects of thermal and mechanical delousing on gill health of farmed Atlantic salmon (Salmo salar L.). Aquaculture 2022, 552, 738019. [Google Scholar] [CrossRef]
- Wolf, J.C.; Baumgartner, W.A.; Blazer, V.S.; Camus, A.C.; Engelhardt, J.A.; Fournie, J.W.; Frasca, S.; Groman, D.B.; Kent, M.L.; Khoo, L.H.; et al. Nonlesions, misdiagnoses, missed diagnoses, and other interpretive challenges in fish histopathology studies: A guide for investigators, authors, reviewers, and readers. Toxicol. Pathol. 2015, 43, 297–325. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.O.; Baxter, E.J.; Holland, C.; Rodger, H.D. Development of a novel histopathological gill scoring protocol for assessment of gill health during a longitudinal study in marine-farmed Atlantic salmon (Salmo salar). Aquac. Int. 2012, 20, 813–825. [Google Scholar] [CrossRef]
- Nylund, S.; Nylund, A.; Watanabe, K.; Arnesen, C.E.; Karlsbakk, E. Paranucleospora theridion n. Gen., n. Sp. (Microsporidia, Enterocytozoonidae) with a life cycle in the salmon louse (Lepeophtheirus salmonis, Copepoda) and Atlantic Salmon (Salmo Salar). J. Eukaryot. Microbiol. 2010, 57, 95–114. [Google Scholar] [CrossRef] [PubMed]
- Gjessing, M.C.; Yutin, N.; Tengs, T.; Senkevich, T.; Koonin, E.; Rønning, H.P.; Alarcon, M.; Ylving, S.; Lie, K.-I.; Saure, B.; et al. Salmon gill poxvirus, the deepest representative of the Chordopoxvirinae. J. Virol. 2015, 89, 9348–9367. [Google Scholar] [CrossRef] [PubMed]
- Fringuelli, E.; Gordon, A.W.; Rodger, H.; Welsh, M.D.; Graham, D.A. Detection of Neoparamoeba perurans by duplex quantitative taqman real-time PCR in formalin-fixed, paraffin-embedded Atlantic salmonid gill tissues. J. Fish Dis. 2012, 35, 711–724. [Google Scholar] [CrossRef]
- Bruno, D.; Collet, B.; Turnbull, A.; Kilburn, R.; Walker, A.; Pendrey, D.; McIntosh, A.; Urquhart, K.; Taylor, G. Evaluation and development of diagnostic methods for Renibacterium salmoninarum causing bacterial kidney disease (BKD) in the UK. Aquaculture 2007, 269, 114–122. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D. lepeophtherii | Ca. B. cysticola | N. perurans | SGPV | |||||
|---|---|---|---|---|---|---|---|---|
| Week | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) |
| 1 | 0% | - | 100% | 30.5 (1.8) | 0% | - | 100% | 23.7 (3.2) |
| 6 | 17% | 31.8 (NA) | 100% | 25.5 (2.3) | 0% | - | 0% | - |
| 10 | 67% | 30.7 (1.9) | 100% | 23.9 (2.2) | 0% | - | 0% | - |
| 14 | 100% | 31.3 (4.9) | 100% | 23.7 (1.1) | 0% | - | 0% | - |
| 19 | 100% | 29.0 (1.9) | 100% | 19.2 (2.1) | 0% | - | 0% | - |
| 23 | 100% | 28.7 (3.5) | 100% | 20.2 (2.5) | 0% | - | 0% | - |
| 28 | 100% | 24.5 (3.4) | 100% | 23.7 (1.0) | 0% | - | 100% | 32.7 (0.7) |
| 30 | 100% | 24.5 (1.1) | 100% | 25.2 (2.2) | 0% | - | 100% | 32.8 (4.0) |
| 32 | 100% | 24.4 (5.2) | 100% | 24.5 (3.0) | 0% | - | 0% | - |
| 34 | 100% | 26.1 (4.0) | 100% | 26.0 (4.2) | 0% | - | 0% | - |
| 36 | 100% | 23.7 (2.4) | 100% | 25.5 (3.2) | 0% | - | 17% | 34.3 (NA) |
| 38 | 100% | 23.7 (2.2) | 100% | 24.8 (2.4) | 0% | - | 33% | 35.6 (0.7) |
| 40 | 100% | 25.5 (2.2) | 100% | 25.5 (2.5) | 17% | 30.9 (NA) | 0% | - |
| 43 | 100% | 22.8 (2.0) | 100% | 23.6 (0.9) | 42% | 31.3 (5.5) | 0% | - |
| 45 | 100% | 24.7 (0.8) | 100% | 24.6 (1.8) | 17% | 36.4 (NA) | 0% | - |
| 47 | 100% | 25.9 (2.3) | 100% | 27.1 (2.5) | 17% | 22.8 (NA) | 0% | - |
| 49 | 100% | 26.2 (1.7) | 100% | 27.7 (3.5) | 0% | - | 17% | 36.1 (NA) |
| 52 | 100% | 27.1 (3.2) | 100% | 29.9 (4.8) | 0% | - | 17% | 32.5 (NA) |
| 54 | 100% | 28.0 (2.5) | 100% | 27.0 (2.8) | 0% | - | 0% | - |
| 57 | 100% | 27.8 (1.9) | 100% | 27.3 (2.3) | 0% | - | 0% | - |
| D. lepeophtherii | Ca. B. cysticola | N. perurans | SGPV | |||||
|---|---|---|---|---|---|---|---|---|
| Week | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) | % Positive | Ct Mean (sd) |
| 6 | 0% | - | 83% | 29.3 (4.0) | 0% | - | 67% | 33.8 (1.7) |
| 10 | 0% | - | 83% | 25.7 (0.7) | 0% | - | 0% | - |
| 14 | 0% | - | 100% | 24.5 (1.8) | 0% | - | 0% | - |
| 19 | 0% | - | 100% | 20.1 (2.0) | 0% | - | 0% | - |
| 23 | 0% | - | 100% | 19.1 (2.7) | 33% | 31.8 (2.8) | 0% | - |
| 28 | 17% | 33.7 (NA) | 100% | 24.1 (2.4) | 100% | 30.5 (5.5) | 17% | 25.6 (NA) |
| 30 | 17% | 32.5 (NA) | 100% | 24.6 (1.2) | 100% | 24.6 (4.9) | 0% | - |
| 32 | 86% | 29.0 (4.1) | 100% | 23.4 (1.0) | 100% | 21.3 (2.4) | 86% | 32.6 (2.0) |
| 34 | 100% | 26.5 (3.4) | 100% | 24.3 (1.9) | 100% | 21.1 (2.3) | 100% | 33.5 (3.5) |
| 36 | 100% | 25.1 (3.4) | 100% | 24.5 (2.3) | 100% | 24.1 (2.3) | 17% | 36.3 (NA) |
| 38 | 100% | 26.1 (2.0) | 100% | 23.1 (1.1) | 100% | 20.1 (3.8) | 0% | - |
| 40 | 100% | 25.0 (0.9) | 100% | 23.1 (2.6) | 100% | 18.6 (3.4) | 0% | - |
| 43 | 100% | 22.5 (1.9) | 100% | 22.9 (0.7) | 100% | 25.4 (8.5) | 83% | 35.4 (1.9) |
| 45 | 100% | 23.9 (1.9) | 100% | 23.3 (1.6) | 100% | 25.4 (4.9) | 67% | 34.5 (2.6) |
| 47 | 100% | 24.1 (3.0) | 100% | 23.9 (1.7) | 100% | 22.2 (6.3) | 17% | 27.9 (NA) |
| 49 | 100% | 23.6 (1.2) | 100% | 27.5 (2.1) | 100% | 25.7 (6.6) | 33% | 37.4 (0.9) |
| 52 | 100% | 22.6 (1.5) | 100% | 23.6 (2.3) | 100% | 27.4 (4.7) | 0% | - |
| 54 | 100% | 23.0 (1.9) | 100% | 24.7 (5.4) | 100% | 30.8 (6.7) | 0% | - |
| 57 | 100% | 23.7 (2.5) | 100% | 27.6 (3.7) | 0% | - | 0% | - |
| Week Number | Farm A | Farm B | Season |
|---|---|---|---|
| 1 | 05/02/2016 | - | Winter-1 |
| 6 | 10/03/2016 | 10/03/2016 | Winter-1 |
| 10 | 05/04/2016 | 06/04/2016 | Spring |
| 14 | 03/05/2016 | 04/05/2016 | Spring |
| 19 | 06/06/2016 | 07/06/2016 | Spring |
| 23 | 07/07/2016 | 08/07/2016 | Summer |
| 28 | 10/08/2016 | 11/08/2016 | Summer |
| 30 | 25/08/2016 | 26/08/2016 | Summer |
| 32 | 06/09/2016 | 07/09/2016 | Summer |
| 34 | 20/09/2016 | 21/09/2016 | Autumn |
| 36 | 04/10/2016 | 05/10/2016 | Autumn |
| 38 | 18/10/2016 | 19/10/2016 | Autumn |
| 40 | 01/11/2016 | 02/11/2016 | Autumn |
| 43 | 22/11/2016 | 23/11/2016 | Autumn |
| 45 | 07/12/2016 | 06/12/2016 | Autumn |
| 47 | 20/12/2016 | 19/12/2016 | Autumn |
| 49 | 06/01/2017 | 05/01/2017 | Winter-2 |
| 52 | 25/01/2017 | 24/01/2017 | Winter-2 |
| 54 | 08/02/2017 | 09/02/2017 | Winter-2 |
| 57 | 28/02/2017 | 01/03/2017 | Winter-2 |
| Primers and Probes | Target Pathogen | Sequence | Reference |
| Fwd_Des | CGGACAGGGAGCATGGTATAG | [44] | |
| Rev_Des | Desmozoon lepeophtherii | GGTCCAGGTTGGGTCTTGAG | |
| Probe_Des | TTGGCGAAGAATGAAA | ||
| Fwd_sgpv | ATCCAAAATACGGAACATAAGCAAT | [45] | |
| Rev_sgpv | CAACGACAAGGAGATCAACGC | ||
| Probe_sgpv | Salmon gill poxvirus | CTCAGAAACTTCAAAGGA | |
| Fwd_Branch | Candidatus | AATACATCGGAACGTGTCTAGTG | [11] |
| Rev_Branch | Branchiomonas | GCCATCAGCCGCTCATGTG | |
| Probe_Branch | cysticola | CTCGGTCCCAGGCTTTCCTCTCCCA | |
| Fwd_Neop | GTTCTTTCGGGAGCTGGGAG | [46] | |
| Rev_Neop | GAACTATCGCCGGCACAAAAG | ||
| Probe_Neop | Neoparamoeba perurans | CAATGCCATTCTTTTCGGA | |
| Fwd_ELF | GGCCAGATCTCCCAGGGCTAT | [47] | |
| Rev_ELF | TGAACTTGCAGGCGATGTGA | ||
| Probe_ELF | Elongation factor 1 α | CCTGTGCTGGATTGCCATACTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrero, A.; Rodger, H.; Hayward, A.D.; Cousens, C.; Bron, J.E.; Dagleish, M.P.; Thompson, K.D. Prospective Longitudinal Study of Putative Agents Involved in Complex Gill Disorder in Atlantic salmon (Salmo salar). Pathogens 2022, 11, 878. https://doi.org/10.3390/pathogens11080878
Herrero A, Rodger H, Hayward AD, Cousens C, Bron JE, Dagleish MP, Thompson KD. Prospective Longitudinal Study of Putative Agents Involved in Complex Gill Disorder in Atlantic salmon (Salmo salar). Pathogens. 2022; 11(8):878. https://doi.org/10.3390/pathogens11080878
Chicago/Turabian StyleHerrero, Ana, Hamish Rodger, Adam D. Hayward, Chris Cousens, James E. Bron, Mark P. Dagleish, and Kim D. Thompson. 2022. "Prospective Longitudinal Study of Putative Agents Involved in Complex Gill Disorder in Atlantic salmon (Salmo salar)" Pathogens 11, no. 8: 878. https://doi.org/10.3390/pathogens11080878
APA StyleHerrero, A., Rodger, H., Hayward, A. D., Cousens, C., Bron, J. E., Dagleish, M. P., & Thompson, K. D. (2022). Prospective Longitudinal Study of Putative Agents Involved in Complex Gill Disorder in Atlantic salmon (Salmo salar). Pathogens, 11(8), 878. https://doi.org/10.3390/pathogens11080878