
First Isolation and Characteristics of Bovine Parainfluenza Virus Type 3 from Yaks


Abstract
:1. Introduction
2. Results
2.1. Pathogens Detection and Isolation
2.2. Indirect Immunofluorescence Assay (IIFA), Transmission Electron Microscopy, and Hemagglutination (HA) Test
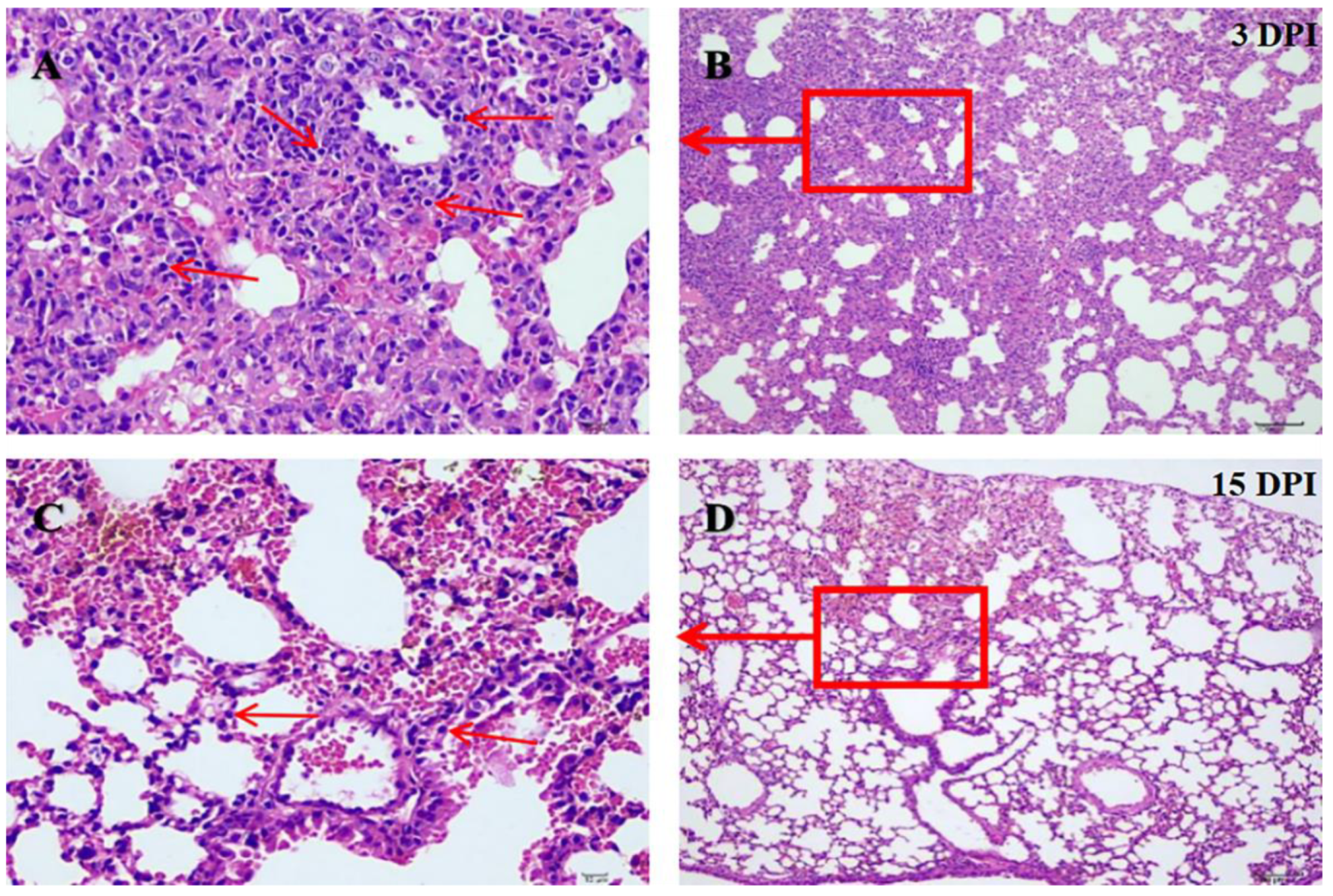
2.3. Experimental Infection of Yak/GZ01/20/CH to BALB/c Mice
2.4. Genomic Characteristics of BPIV3 from Yak
3. Discussion
4. Materials and Methods
4.1. Clinical Samples and Cell Lines
4.2. Sample Processing and Nucleic Acid Extraction
4.3. Pathogens Detection
4.4. Virus Isolation, Purification, and Titer Determination
4.5. IIFA
4.6. Transmission Electron Microscopy
4.7. HA Test
4.8. Experimental Infection of BALB/c Mice with the BPIV3 Isolate from Yak
4.9. Complete Genome Amplification
4.10. Sequence and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Miles, D.G. Overview of the North American beef cattle industry and the incidence of bovine respiratory disease (BRD). Anim. Health Res. Rev. 2009, 10, 101–103. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.A. Bovine parainfluenza-3 virus. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 575–593. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; Janke, B.H.; Elankumaran, S. Molecular characterization of glycoprotein genes and phylogenetic analysis of two swine paramyxoviruses isolated from United States. Virus Genes 2009, 39, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.; Wernery, U.; Lee, H.; Fung, J.; Joseph, S.; Li, K.; Elizabeth, S.; Fong, J.; Chan, K.; Chen, H.; et al. Co-circulation of a Novel Dromedary Camel Parainfluenza Virus 3 and Middle East Respiratory Syndrome Coronavirus in a Dromedary Herd With Respiratory Tract Infections. Front. Microbiol. 2021, 12, 739779. [Google Scholar] [CrossRef]
- Eberle, K.C.; Neill, J.D.; Venn-Watson, S.K.; McGill, J.L.; Sacco, R.E. Novel Atlantic bottlenose dolphin parainfluenza virus TtPIV-1 clusters with bovine PIV-3 genotype B strains. Virus Genes 2015, 51, 198–208. [Google Scholar] [CrossRef]
- Baghezza, S.; Mamache, B.; Bennoune, O.; Ghougal, K. Pathological study and detection of Bovine parainfluenza 3 virus in pneumonic sheep lungs using direct immunofluorescence antibody technique. Comp. Clin. Pathol. 2021, 30, 301–310. [Google Scholar] [CrossRef]
- Buchmeier, M.; Clegg, J.; Franze-Fernandez, M. Virus taxonomy. Sixth Report of the International Committee on Taxonomy of Viruses. Arch. Virol. 1995, 10, 350–354. [Google Scholar]
- Tashiro, M.; Homma, M. Protection of mice from wild-type Sendai virus infection by a trypsin-resistant mutant, TR-2. J. Virol. 1985, 53, 228. [Google Scholar] [CrossRef]
- Suzu, S.; Sakai, Y.; Shioda, T.; Shibuta, H. Nucleotide sequence of the bovine parainfluenza 3 virus genome: The genes of the F and HN glycoproteins. Nucleic Acids Res. 1987, 15, 2945–2958. [Google Scholar] [CrossRef]
- Yuko, S.; Shinya, S.; Tatsuo, S.; Hiroshi, S. Nucleotide sequence of the bovine parainfluenza 3 virus genome: Its 3′ end and the genes of NP, P., C and M proteins. Nucleic Acids Res. 1987, 15, 2927. [Google Scholar]
- Pelet, T.; Curran, J.; Kolakofsky, D. The P gene of bovine parainfluenza virus 3 expresses all three reading frames from a single mRNA editing site. EMBO J. 1991, 10, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Bailly, J.E.; Mcauliffe, J.M.; Skiadopoulos, M.H.; Collins, P.L.; Murphy, B.R. Sequence Determination and Molecular Analysis of Two Strains of Bovine Parainfluenza Virus Type 3 that are Attenuated for Primates. Virus Genes 2000, 20, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Horwood, P.F.; Gravel, J.L.; Mahony, T.J. Identification of two distinct bovine parainfluenza virus type 3 genotypes. J. Gen. Virol 2008, 89, 1643–1648. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Shi, H.F.; Gao, Y.R.; Xin, J.Q.; Liu, N.H.; Xiang, W.H.; Ren, X.G.; Feng, J.K.; Zhao, L.P.; Xue, F. Isolation and genetic characterization of bovine parainfluenza virus type 3 from cattle in China. Vet. Microbiol. 2011, 149, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Maidana, S.; Lomonaco, M.; Odeon, A.; Combessies, G.; Graig, M.I.; Rodriguez, D.; Parenno, V.; Zabal, O.; Konrad, J.L.; Crudelli, G. Isolation and characterization of bovine parainfluenza virus type 3 from water buffaloes (Bubalus bulalis) in Argentina. Rev. Colomb. Psiquiatr. 2012, 8, 249–260. [Google Scholar]
- Ohkura, T.; Kokuho, T.; Konishi, M.; Kameyama, K.I.; Takeuchi, K. Complete Genome Sequences of Bovine Parainfluenza Virus Type 3 Strain BN-1 and Vaccine Strain BN-CE. Genome Announc. 2013, 1, e00247-12. [Google Scholar] [CrossRef]
- Oem, J.K.; Lee, E.Y.; Lee, K.K.; Kim, S.H.; Hyun, B.H. Molecular characterization of a Korean bovine parainfluenza virus type 3 isolate. Vet. Microbiol. 2012, 162, 224–227. [Google Scholar] [CrossRef]
- Wen, Y.J.; Shi, X.C.; Wang, F.X.; Wang, W.; Zhang, S.Q.; Li, G.; Song, N.; Chen, L.Z.; Cheng, S.P.; Wu, H. Phylogenetic analysis of the bovine parainfluenza virus type 3 from cattle herds revealing the existence of a genotype A strain in China. Virus Genes 2012, 45, 542–547. [Google Scholar] [CrossRef]
- Neill, J.D.; Ridpath, J.F.; Valayudhan, B.T. Identification and genome characterization of genotype B and genotype C bovine parainfluenza type 3 viruses isolated in the United States. BMC Vet. Res. 2015, 11, 112. [Google Scholar] [CrossRef]
- Leal, E.; Liu, C.; Zhao, Z.; Deng, Y.; Villanova, F.; Liang, L.; Li, J.; Cui, S. Isolation of a Divergent Strain of Bovine Parainfluenza Virus Type 3 (BPIV3) Infecting Cattle in China. Viruses 2019, 11, 489. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, J.; Chen, X.; Wei, X.; Wu, C.; Cui, Q.; Hao, Y. Investigation of viral pathogens in cattle with bovine respiratory disease complex in Inner Mongolia, China. Microb. Pathog. 2021, 153, 104594. [Google Scholar] [CrossRef] [PubMed]
- Mi, R.; Wang, X.; Li, C.; Huang, Y.; Zhou, P.; Li, Z.; Lei, M.; Cai, J.; Chen, Z. Prevalence and genetic characterization of Cryptosporidium in yaks in Qinghai Province of China. PLoS ONE 2013, 8, e74985. [Google Scholar]
- Chen, Z.; Wang, J.; Ma, J.; Li, S.; Huo, S.; Yang, Y.; Zhaxi, Y.; Zhao, Y.; Zhang, D. Transcriptome and proteome analysis of pregnancy and postpartum anoestrus ovaries in yak. J. Vet. Sci 2022, 23, e3. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Wang, D.; Yan, M.; Chang, Z.; Xu, Y.; Sizhu, S.; Li, Z.; Hu, S.; Bi, D. Isolation, identification and biological characteristics of Mycoplasma bovis in yaks. Microb. Pathog. 2021, 150, 104691. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Liu, L.; Zheng, F.; Chen, Q.; Li, Z.; Cao, X.; Yin, H.; Zhou, J.; Cai, X. Molecular investigation of bovine viral diarrhea virus infection in yaks (Bos gruniens) from Qinghai, China. Virol. J. 2014, 11, 29. [Google Scholar] [CrossRef]
- He, Q.; Guo, Z.; Zhang, B.; Yue, H.; Tang, C. First detection of bovine coronavirus in Yak (Bos grunniens) and a bovine coronavirus genome with a recombinant HE gene. J. Gen. Virol. 2019, 100, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Shibuta, H.; Kanda, T.; Adachi, A.; Yogo, Y. Characterization of bovine parainfluenza virus type 3. Microbiol. Immunol. 1979, 23, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Moloney, M.; Pye, D.; Smith, H.; Scott, P. Isolation of parainfluenza virus from dogs. Aust. Vet. J. 1985, 62, 285–286. [Google Scholar] [CrossRef]
- Henrickson, K.J. Parainfluenza viruse.es. Clin. Microbiol. Rev. 2003, 16, 242–264. [Google Scholar] [CrossRef]
- Dong, X.M.; Zhu, Y.M.; Cai, H.; Lv, C.; Gao, Y.R.; Yu, Z.; Xue, F. Studies on the pathogenesis of a Chinese strain of bovine parainfluenza virus type 3 infection in Balb/c mice. Vet. Microbiol. 2012, 158, 199–204. [Google Scholar] [CrossRef]
- Bryson, D.; Adair, B.; McNulty, M.; McAliskey, M.; Bradford, H.; Allan, G.; Evans, R.; Forster, F. Studies on the efficacy of intranasal vaccination for the prevention of experimentally induced parainfluenza type 3 virus pneumonia in calves. Vet. Rec. 1999, 145, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Bergman, R.; Moreno-Lopez, J.; Möllerberg, L.; Morein, B. Parainfluenza-3 virus: Difference in capacity of neuraminidase-weak and strong strains to infect young calves and to elicit cellular immune response. Res. Vet. Sci. 1978, 25, 193–199. [Google Scholar] [CrossRef]
- Komatsu, T.; Takeuchi, K.; Gotoh, B. Bovine parainfluenza virus type 3 accessory proteins that suppress beta interferon production. Microbes Infect. 2007, 9, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chang, X.; Yao, W.; Wei, N.; Huo, N.; Wang, Y.; Wei, Q.; Liu, H.; Wang, X.; Zhang, S.; et al. Host CARD11 Inhibits Newcastle Disease Virus Replication by Suppressing Viral Polymerase Activity in Neurons. J. Virol. 2019, 93, e01499-19. [Google Scholar] [CrossRef]
- Chen, X.; Jia, Y.; Wei, N.; Ye, C.; Hao, H.; Xiao, S.; Wang, X.; Liu, H.; Yang, Z. Identification of a new amino acid mutation in the HN protein of NDV involved in pathogenicity. Vet. Res. 2021, 52, 147. [Google Scholar] [CrossRef]
- Haller, A.A.; Macphail, M.; Mitiku, M.; Tang, R.S. A Single Amino Acid Substitution in the Viral Polymerase Creates a Temperature-Sensitive and Attenuated Recombinant Bovine Parainfluenza Virus Type 3. Virology 2001, 288, 342–350. [Google Scholar] [CrossRef]
- Itoh, M.; Isegawa, Y.; Hotta, H.; Homma, M. Isolation of an avirulent mutant of Sendai virus with two amino acid mutations from a highly virulent field strain through adaptation to LLC-MK2 cells. J. Gen. Virol. 1997, 78, 3207–3215. [Google Scholar] [CrossRef]
- Lan, D.; Xiong, X.; Huang, C.; Mipam, T.D.; Li, J. Toward Understanding the Genetic Basis of Yak Ovary Reproduction: A Characterization and Comparative Analyses of Estrus Ovary Transcriptiome in Yak and Cattle. PLoS ONE 2016, 11, e0152675. [Google Scholar] [CrossRef]
- Alexander, T.W.; Cook, S.R.; Yanke, L.J.; Booker, C.W.; Morley, P.S.; Read, R.R.; Gow, S.P.; McAllister, T.A. A multiplex polymerase chain reaction assay for the identification of Mannheimia haemolytica, Mannheimia glucosida and Mannheimia ruminalis. Vet. Microbiol. 2008, 130, 165–175. [Google Scholar] [CrossRef]
- Gome, D.E.; Arroyo, L.G.; Poljak, Z.; Viel, L.; Weese, J.S. Detection of Bovine Coronavirus in Healthy and Diarrheic Dairy Calves. J. Vet. Intern. Med. 2017, 31, 1884–1891. [Google Scholar] [CrossRef]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Kisch, A.L.; Johnson, K.M. A Plaque Assay for Respiratory Syncytial Virus. Exp. Biol. Med. 1963, 112, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per cent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Horwood, P.F.; Mahony, T.J. Multiplex real-time RT-PCR detection of three viruses associated with the bovine respiratory disease complex. J. Virol. Methods 2011, 171, 360–363. [Google Scholar]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organs | Day 1 | Day 3 | Day 6 | Day 9 | Day 12 | Day 15 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | |
| Lung | − | + | + | + | + | + | + | + | − | + | + | + | − | + | − | + | + | − |
| Trachea | + | + | + | + | + | + | + | + | + | − | + | − | + | + | + | − | − | + |
| Heart | − | − | − | − | + | − | − | − | + | + | − | + | + | − | − | − | − | − |
| Liver | − | − | − | + | + | + | − | + | + | + | + | − | − | − | − | + | − | − |
| Spleen | − | − | − | − | + | − | + | + | − | + | + | − | − | − | + | − | − | − |
| Kidney | − | − | − | + | − | + | + | − | − | − | + | + | + | − | − | − | − | − |
| Blood | − | − | − | + | + | − | + | − | − | + | + | − | − | + | − | + | − | − |
| Gene Name | Nucleotide Differences | Amino Acid Differences | ||||
|---|---|---|---|---|---|---|
| 13 BPIV3 Genotype C Strains | Position | Three Yak BPIV3 Strains | 13 BPIV3 Genotype C Strains | Position | Three Yak BPIV3 Strains | |
| N | T | 1401 | C | - | - | - |
| P | T/A | 781 | C | Y/N | 261 | H |
| M | T | 321 | C | - | - | - |
| F | A; C; T; C | 81; 714; 1035; 1380 | G; T; G; T | - | - | - |
| HN | G; G | 145; 1650 | A; A | I; I | 49; 550 | V; M |
| L | G; T | 3293; 4389 | A; C | -; S | -; 1098 | -; N |
| Pathogens | Primer Name | Sequence (5′–3′) | Size (bp) | Target Gene |
|---|---|---|---|---|
| BPIV3 | BPIV3-F | GAATGACTCATGATAGAGGTAT | 647 | HN |
| BPIV3-R | AGGACAACCAGTTGTATTACAT | |||
| BVDV | BVDV-F | ATGCCCTTAGTAGGACTAG | 287 | 5′ UTR |
| BVDV-R | TCAACTCCATGTGCCATGT | |||
| BHV-1 | BHV-F | ATGCCGCGATACAACTACACTGAAC | 921 | gD |
| BHV-R | TTATTCGAGGCTCGGCCAGCCTT | |||
| BRSV | BRSV-F | ATGGCTCTTAGCAAGGTC | 459 | N |
| BRSV-R | AGAGTCATGTCTGTATTC | |||
| BCoV | BCoV-F | CTAGTAACCAGGCTGATGTCAATACC | 81 | N |
| BCoV-R | GGCGGAAACCTAGTCGGAATA | |||
| M. bovis | M.bovis-F | TAATTTAGAAGCTTTAAATGAGCGC | 238 | uvrC |
| M.bovis-R | CATATCTAGGTCAATTAAGGCTTTG | |||
| P. multocida | P-F | ATCCGCTATTTACCCAGTGG | 460 | KMT1 |
| P-R | GCTGTAAACGAACTCGCCAC | |||
| M. haemolytica | M-F | GCAGGAGGTGATTATTAAAGTGG | 206 | lktD |
| M-R | CAGCAGTTATTGTCATACCTGAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Chen, X.; Tang, C.; Yue, H. First Isolation and Characteristics of Bovine Parainfluenza Virus Type 3 from Yaks. Pathogens 2022, 11, 962. https://doi.org/10.3390/pathogens11090962
Ren Y, Chen X, Tang C, Yue H. First Isolation and Characteristics of Bovine Parainfluenza Virus Type 3 from Yaks. Pathogens. 2022; 11(9):962. https://doi.org/10.3390/pathogens11090962
Chicago/Turabian StyleRen, Yunxin, Xi Chen, Cheng Tang, and Hua Yue. 2022. "First Isolation and Characteristics of Bovine Parainfluenza Virus Type 3 from Yaks" Pathogens 11, no. 9: 962. https://doi.org/10.3390/pathogens11090962
APA StyleRen, Y., Chen, X., Tang, C., & Yue, H. (2022). First Isolation and Characteristics of Bovine Parainfluenza Virus Type 3 from Yaks. Pathogens, 11(9), 962. https://doi.org/10.3390/pathogens11090962