
Biological Control of Verticillium Wilt and Growth Promotion in Tomato by Rhizospheric Soil-Derived Bacillus amyloliquefaciens Oj-2.16
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation and Purification of Rhizospheric Soil Bacteria of O. japonicus
2.3. Determination of Bacterial Antagonistic Activity
2.4. Scanning Electron Microscopy Observations of the Inhibited Hyphae of Pathogenic Fungi
2.5. Morphological, Physiological, and Biochemical Characteristics of Strain Oj-2.16
2.6. 16S rDNA Sequence Identification and Phylogenetic Analyses
2.7. PCR Amplification of Lipopeptide and Dipeptide Biosynthetic Genes
2.8. Biocontrol and Growth-Promoting Effects of the Strain Oj-2.16 on Tomato in Pot Experiments
2.9. Antioxidant Enzyme Activity Analysis
3. Results
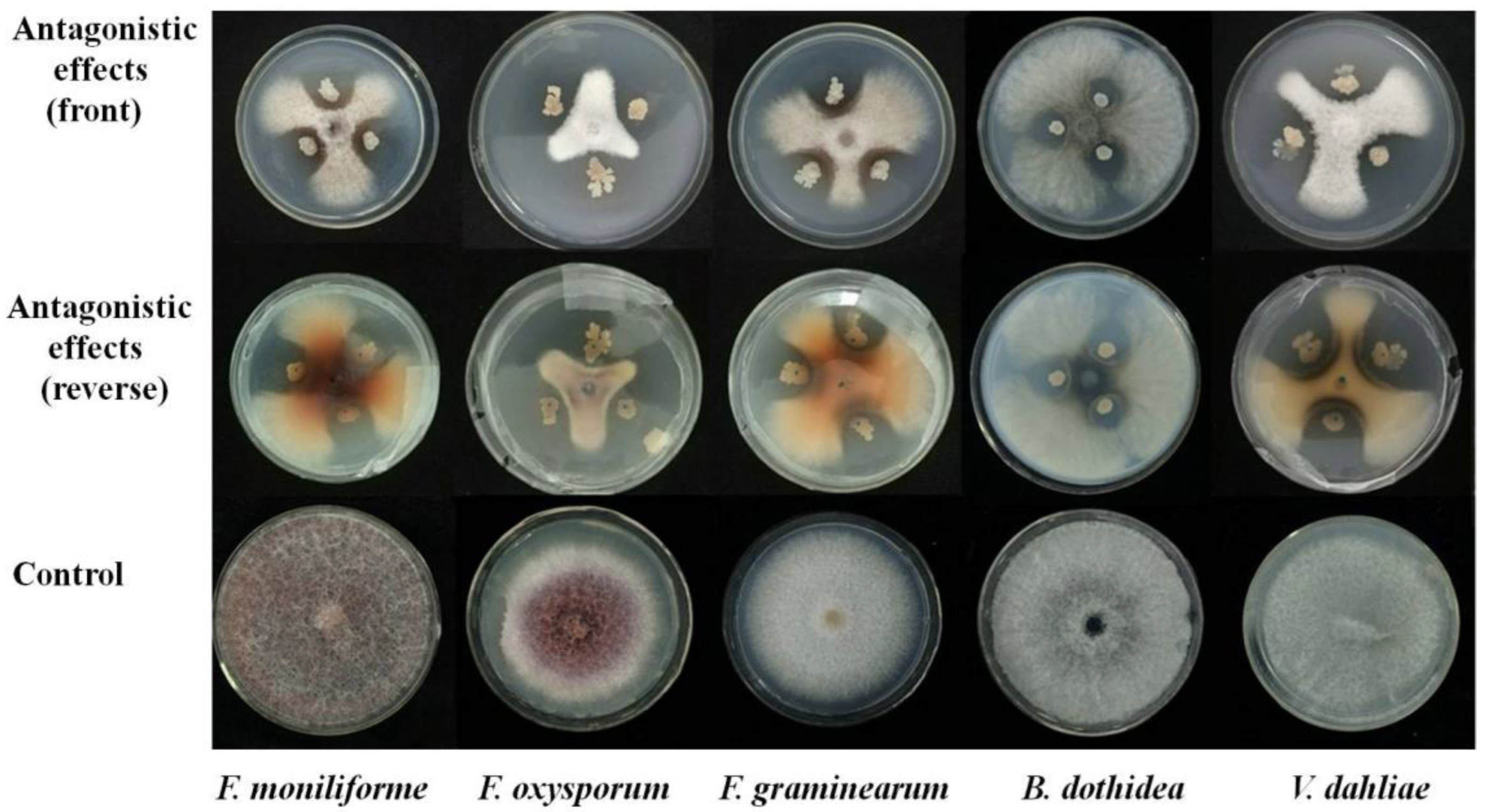
3.1. Isolation of Bacteria and Screening of Antagonistic Strains against Pathogenic Fungi
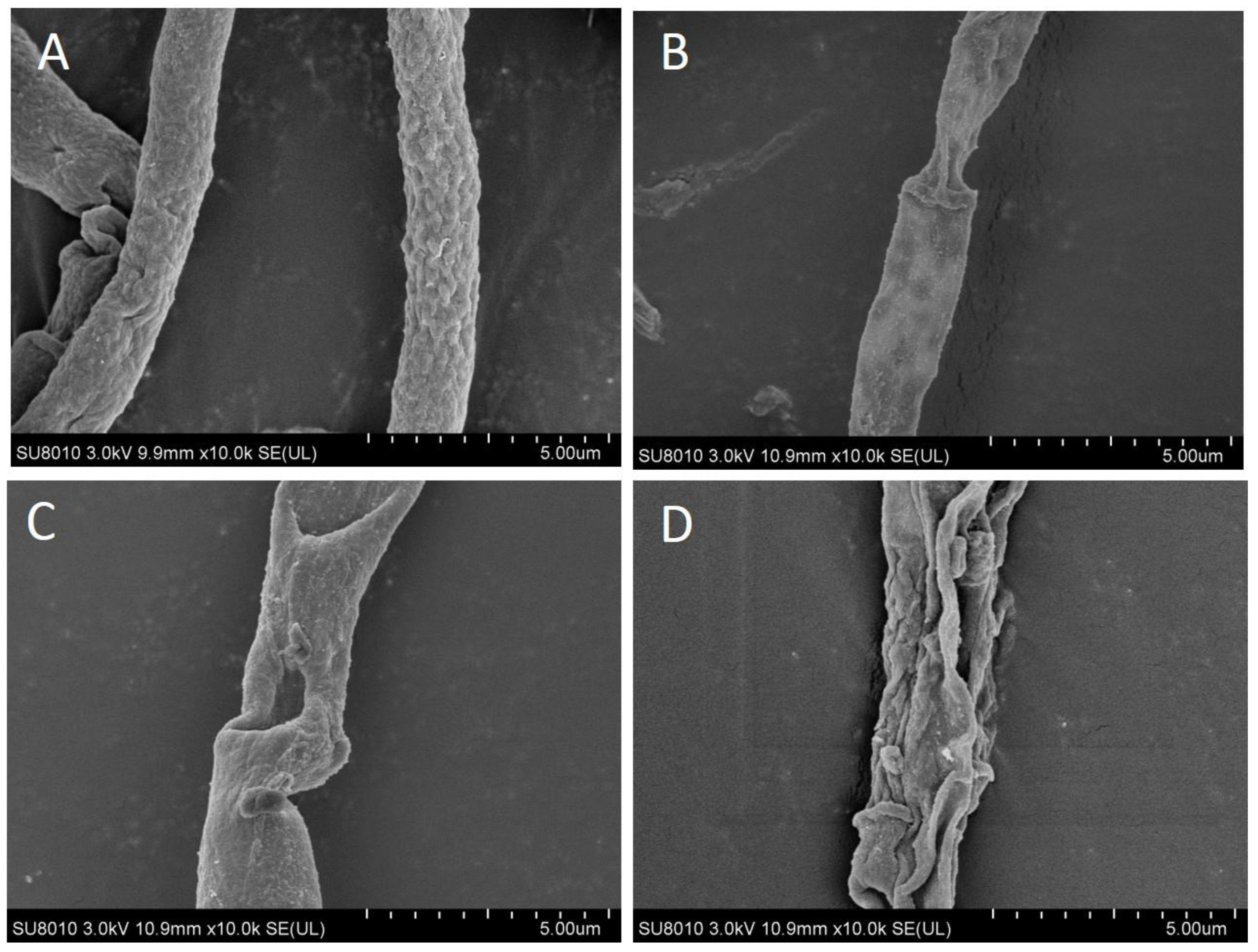
3.2. Scanning Electron Microscopy Observation of Inhibited Hyphae of Pathogen
3.3. Physiological and Biochemical Characteristics of Strain Oj-2.16
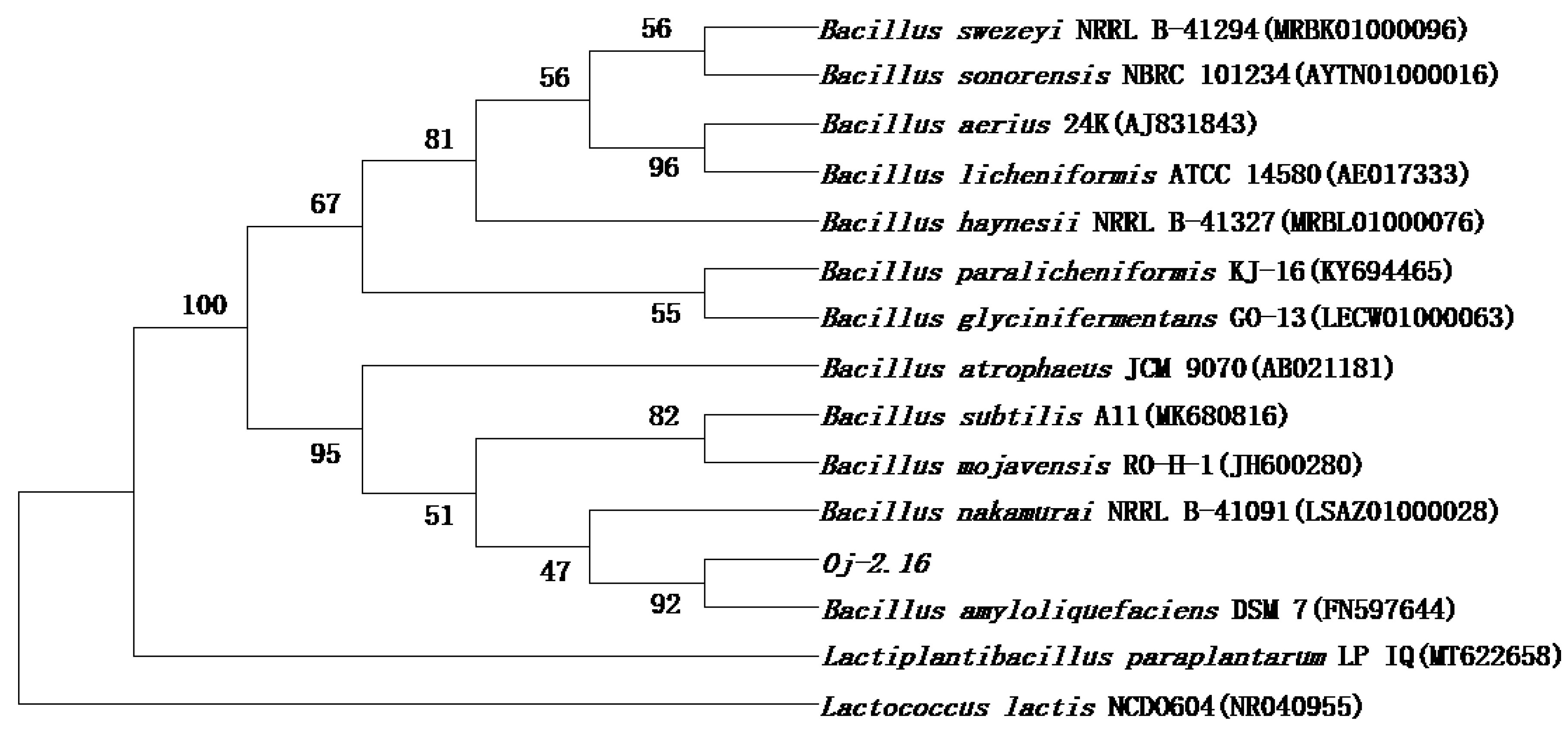
3.4. 16S rDNA Sequencing and Phylogenetic Analysis of Strain Oj-2.16
3.5. PCR Amplification of the Lipopeptide and Dipeptide Biosynthetic Genes from Strain Oj-2.16
3.6. Biological Efficacy of B. amyloliquefaciens Oj-2.16 on Verticillium wilt in Pot Experiments
3.7. Growth-Promoting Effect of B. amyloliquefaciens Oj-2.16 on Tomato Seedlings
3.8. Effects of B. amyloliquefaciens Oj-2.16 on Antioxidant Enzyme Activities and MDA Content of Tomato Seedlings
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fradin, E.F.; Zhang, Z.; Juarez Ayala, J.C.; Castroverde, C.D.; Nazar, R.N.; Robb, J.; Liu, C.M.; Thomma, B.P. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Goicoechea, N. To what extent are soil amendments useful to control Verticillium wilt? Pest Manag. Sci. 2009, 65, 831–839. [Google Scholar] [CrossRef]
- Luo, L.; Zhao, C.; Wang, E.; Raza, A.; Yin, C. Bacillus amyloliquefaciens as an excellent agent for biofertilizer and biocontrol in agriculture: An overview for its mechanisms. Microbiol. Res. 2022, 259, 127016. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.D.; Jeon, B.J.; Han, J.W.; Park, M.Y.; Kang, S.A.; Kim, B.S. Evaluation of the endophytic nature of Bacillus amyloliquefaciens strain GYL4 and its efficacy in the control of anthracnose. Pest Manag. Sci. 2016, 72, 1529–1536. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Asaf, S.; Lee, I.J. Plant growth-promoting endophytic bacteria versus pathogenic infections: An example of Bacillus amyloliquefaciens RWL-1 and Fusarium oxysporum f. sp. Lycopersici in tomato. Peer J. 2017, 5, e3107. [Google Scholar] [CrossRef] [Green Version]
- Ansary, W.R.; Prince, F.R.K.; Haque, E.; Sultana, F.; West, H.M.; Rahman, M.; Mondol, A.M.; Akanda, A.M.; Rahman, M.; Clarke, M.L.; et al. Endophytic Bacillus spp. from medicinal plants inhibit mycelial growth of Sclerotinia sclerotiorum and promote plant growth. Z. Naturforsch. C. J. Biosci. 2018, 73, 247–256. [Google Scholar] [CrossRef]
- Daungfu, O.; Youpensuk, S.; Lumyong, S. Endophytic bacteria isolated from citrus plants for biological control of citrus canker in lime plants. Trop. Life Sci. Res. 2019, 30, 73–88. [Google Scholar] [CrossRef]
- Cui, W.; He, P.; Munir, S.; He, P.; He, Y.; Li, X.; Yang, L.; Wang, B.; Wu, Y.; He, P. Biocontrol of soft rot of Chinese cabbage using an endophytic bacterial strain. Front. Microbiol. 2019, 10, 1471. [Google Scholar] [CrossRef] [Green Version]
- Samaras, A.; Hadjipetrou, C.; Karaoglanidis, G. Bacillus amyloliquefaciens strain QST713 may contribute to the management of SDHI resistance in Botrytis cinerea. Pest Manag. Sci. 2021, 77, 1316–1327. [Google Scholar] [CrossRef]
- Pan, Z.; Munir, S.; Li, Y.; He, P.; He, P.; Wu, Y.; Xie, Y.; Fu, Z.; Cai, Y.; He, Y. Deciphering the Bacillus amyloliquefaciens B9601-Y2 as a potential antagonist of tobacco leaf mildew pathogen during flue-curing. Front. Microbiol. 2021, 12, 683365. [Google Scholar] [CrossRef]
- Yang, L.; Quan, X.; Xue, B.; Goodwin, P.H.; Lu, S.; Wang, J.; Du, W.; Wu, C. Isolation and identification of Bacillus subtilis strain YB-05 and its antifungal substances showing antagonism against Gaeumannomyces graminis var. tritici. Biol. Control 2015, 85, 52–58. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X. Bacillomycin D produced by Bacillus amyloliquefaciens is involved in the antagonistic interaction with the plant-pathogenic fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, e01075-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.; Gibbons, N. Bergeys Manual of Systematic Bacteriology, 9th ed.; Williams and Wilkins Company: Baltimore, ML, USA, 1994; pp. 729–758. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- BLAST Tool. Available online: https://www.ncbi.nlm.nih.gov/BLAST (accessed on 10 December 2022).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis V ersion 7.0 for bigger datasets. Mol Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzand, A.; Moosa, A.; Zubair, M.; Khan, A.R.; Ayaz, M.; Massawe, V.C.; Gao, X. Transcriptional profiling of diffusible lipopeptides and fungal virulence genes during Bacillus amyloliquefaciens EZ1509-mediated suppression of Sclerotinia sclerotiorum. Phytopathology 2020, 110, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, X.; Chen, Y.; Qin, H.; Huang, L.; Han, Q. Inhibitory efficacy of endophytic Bacillus subtilis EDR4 against Sclerotinia sclerotiorum on rapeseed. Biol. Control 2014, 78, 67–76. [Google Scholar] [CrossRef]
- Wu, Z.B.; Li, J.; Feng, C.D.; Zhang, J.F. Research on identification technique of Verticillium wilt resistance in cotton. Hubei Agr. Sci. 1999, 5, 16–19. [Google Scholar] [CrossRef]
- Gao, J.F. Experimental Guidance of Plant Physiology; Higher Education Press: Beijing, China, 2006; pp. 123–130. [Google Scholar]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic acid seed treatment stimulates insecticide detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef]
- Borriss, R.; Chen, X.H.; Rueckert, C.; Blom, J.; Becker, A.; Baumgarth, B.; Fan, B.; Pukall, R.; Schumann, P.; Spröer, C.; et al. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7T and FZB42T: A proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int. J. Syst. Evol. Microbiol. 2011, 61, 1786–1801. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Yang, C.; Wang, Y.; Ma, T.; Cai, F.; Wei, L.; Jin, M.; Osei, R.; Zhang, J.; Tang, M. Potential of an endophytic bacteria Bacillus amyloliquefaciens 3-5 as biocontrol agent against potato scab. Microb. Pathog. 2022, 163, 105382. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Shan, Y.; Liu, S.; Yang, Y.; Liu, Y.; Yin, Y.; Hou, Z.; Luan, P.; Li, R. Antagonistic strain Bacillus amyloliquefaciens XZ34-1 for controlling Bipolaris sorokiniana and promoting growth in wheat. Pathogens 2021, 10, 1526. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, C.; Zuo, B.; Liang, X.; Zhang, D.; Liu, R.; Yang, L.N.; Lu, B.H.; Wang, X.; Gao, J. A novel biocontrol strain Bacillus amyloliquefaciens FS6 for excellent control of gray mold and seedling diseases of ginseng. Plant Dis. 2021, 105, 1926–1935. [Google Scholar] [CrossRef]
- Hu, J.; Zheng, M.; Dang, S.; Shi, M.; Zhang, J.; Li, Y. Biocontrol potential of Bacillus amyloliquefaciens LYZ69 against anthracnose of alfalfa (Medicago sativa). Phytopathology 2021, 111, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.H.; Chuang, C.Y.; Zheng, J.L.; Chen, H.H.; Liang, Y.S.; Huang, T.P.; Lin, Y.H. Bacillus amyloliquefaciens strain PMB05 intensifies plant immune responses to confer resistance against bacterial wilt of tomato. Phytopathology 2020, 110, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Chauhan, A.; Sharma, R.; Sehgal, R.; Shirkot, C.K. Potential of Bacillus amyloliquefaciens for biocontrol of bacterial canker of tomato incited by Clavibacter michiganensis ssp. michiganensis. Microb. Pathog. 2019, 130, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J.; et al. Fengycin produced by Bacillus amyloliquefaciens FZB42 inhibits Fusarium graminearum growth and mycotoxins biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, I.; Berić, T.; Dimkić, I.; Popović, T.; Lozo, J.; Fira, D.; Stanković, S. Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 2019, 126, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.L.; Zhang, M.L.; Kong, Z.R.; Chen, X.; Wang, X.; Ding, W.; Lai, H.X.; Guo, Q. Genomic analysis reveals potential mechanisms underlying promotion of tomato plant growth and antagonism of soilborne pathogens by Bacillus amyloliquefaciens Ba13. Microbiol. Spectr. 2021, 9, e01615. [Google Scholar] [CrossRef]
- Shahid, I.; Han, J.; Hanooq, S.; Malik, K.A.; Borchers, C.H.; Mehnaz, S. Profiling of metabolites of Bacillus spp. and their application in sustainable plant growth promotion and biocontrol. Front. Sust. Food 2021, 5, 37. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, M.; Hamoud, Y.A.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, O.A.A.; Li, L.; Ma, J.B.; Hatab, S.; Xu, L.; Guo, J.W.; Rasulov, B.A.; Liu, Y.H.; Hedlund, B.P.; Li, W.J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, D.; Torres, M.; Sampedro, I.; Martínez-Checa, F.; Torres, B.; Béjar, V. Biological control of Verticillium wilt on olive trees by the salt-tolerant strain Bacillus velezensis XT1. Microorganisms 2020, 8, 1080. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alqueres, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic Lipopeptides of Bacillus amyloliquefaciens subsp. plantarum colonizing the lettuce rhizosphere enhance plant defense responses toward the bottom rot pathogen Rhizoctonia solani. Mol. Plant Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Su, L.; Fang, Z.Y.; Yuan, S.F.; Wang, L.L.; Shen, B.; Shen, Q.R. Competitive use of root exudates by Bacillus amyloliquefaciens with Ralstonia solanacearum decreases the pathogenic population density and effectively controls tomato bacterial wilt. Sci. Hortic. 2017, 218, 132–138. [Google Scholar] [CrossRef]
- Prasannath, K. Plant defense-related enzymes against pathogens: A review. AGRIEAST J. Agricult. Sci. 2017, 11, 38–48. [Google Scholar] [CrossRef]
- Xu, W.; Xu, L.; Deng, X.; Goodwin, P.H.; Xia, M.; Zhang, J.; Wang, Q.; Sun, R.; Pan, Y.; Wu, C.; et al. Biological control of take-all and growth promotion in wheat by Pseudomonas chlororaphis YB-10. Pathogens 2021, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.D.; Dutta, S.; Ryu, H.; Yoo, S.J.; Suh, D.S.; Park, K. Induction of systemic resistance in Panax ginseng against Phytophthora cactorum by native Bacillus amyloliquefaciens HK34. J. Ginseng Res. 2015, 39, 213–220. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipopeptides and Dipeptides | Annealing Temperature | Size (Base Paires) | Primers |
|---|---|---|---|
| Surfacin | 56.3 | 1298 | 5′ CGGTGATCTTGCGAAGCTTTAT 3′ 5′ CGCTTTCGTTCTGCCATTCT 3′ |
| Fengycin | 56.3 | 1447 | 5′ CGGCCATTCGCTCATCTTTTAT 3′ 5′ GTTTCCGCTTCATCAGTCTCTTC 3′ |
| Iturin | 55.9 | 1244 | 5′ ACCTCACCTTGATCGGCTATAC 3′ 5′ TGGTGGGCGAAAGAAGTTTATG 3′ |
| Bacilysin | 52.8 | 657 | 5′ TCATGACTCTTTCGCCTCT 3′ 5′ GAATGGGATAACGGAGTAAGAC3′ |
| Physiological and Biochemical Characteristics | Oj-2.16 |
|---|---|
| gelatin liquefaction | − |
| methyl red reaction | − |
| Voges-Proskauer reaction | + |
| nitrate reduction | + |
| D-glucose utilization | + |
| fructose utilization | + |
| sucrose utilization | − |
| mannitol utilization | + |
| starch hydrolysis | + |
| Gram staining | + |
| IAA production | + |
| siderophore production | + |
| Treatment | Disease Incidence (%) | Disease Index | Biocontrol Efficacy (%) |
|---|---|---|---|
| Vd + Oj-2.16 | 30.00 ± 5.00 c | 7.50 ± 1.25 c | 89.26 ± 2.05 a |
| Vd + Carbendazim | 41.67 ± 7.64 b | 11.67 ± 1.91 b | 83.27 ± 3.21 a |
| Vd | 96.67 ± 2.89 a | 70.00 ± 2.50 a | — — |
| Treatments | Plant Height (cm) | Root Length (cm) | Stem Width (mm) | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|---|
| Oj-2.16 | 16.64 ± 1.26 a | 8.60 ± 0.70 a | 5.73 ± 0.60 a | 4.02 ± 0.55 a | 0.41 ± 0.06 a |
| Vd + Oj-2.16 | 15.98 ± 1.51 a | 6.72 ± 0.71 b | 5.03 ± 0.54 b | 3.36 ± 0.35 b | 0.33 ± 0.04 b |
| Vd + Carbendazim | 12.43 ± 1.35 b | 4.55 ± 0.67 c | 4.09 ± 0.42 c | 2.58 ± 0.38 c | 0.27 ± 0.03 c |
| CK | 11.52 ± 1.13 b | 3.87 ± 0.52 c | 3.18 ± 0.45 d | 2.55 ± 0.40 c | 0.25 ± 0.03 c |
| Vd | 9.04 ± 0.79 c | 3.02 ± 0.38 d | 2.40 ± 0.49 e | 1.79 ± 0.40 d | 0.19 ± 0.04 d |
| Treatments | SOD (U/mg Protein) | POD (U/mg Protein) | CAT (U/mg Protein) | PAL (U/mg Protein) | MDA (µmol·g−1 FW) |
|---|---|---|---|---|---|
| Vd+Oj-2.16 | 7.24 ± 0.27 a | 17.86 ± 0.43 a | 28.58 ± 0.77 a | 85.76 ± 1.18 a | 8.16 ± 0.53 c |
| Vd | 6.28 ± 0.31 b | 15.77 ± 0.40 b | 18.79 ± 0.40 c | 60.77 ± 1.16 c | 14.74 ± 0.60 a |
| Vd+Carbendazim | 4.41 ± 0.37 d | 14.29 ± 0.40 c | 24.65 ± 0.65 b | 72.27 ± 0.97 b | 10.21 ± 0.49 b |
| Oj-2.16 | 5.39 ± 0.44 c | 12.08 ± 0.31 d | 15.89 ± 0.61 d | 43.46 ± 0.78 d | 5.25 ± 0.59 d |
| CK | 3.57 ± 0.29 e | 8.81 ± 0.30 e | 10.12 ± 0.43 e | 22.52 ± 0.58 e | 4.36 ± 0.46 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, D.; Zhang, Q.; Zhu, X.; Zhang, L. Biological Control of Verticillium Wilt and Growth Promotion in Tomato by Rhizospheric Soil-Derived Bacillus amyloliquefaciens Oj-2.16. Pathogens 2023, 12, 37. https://doi.org/10.3390/pathogens12010037
Pei D, Zhang Q, Zhu X, Zhang L. Biological Control of Verticillium Wilt and Growth Promotion in Tomato by Rhizospheric Soil-Derived Bacillus amyloliquefaciens Oj-2.16. Pathogens. 2023; 12(1):37. https://doi.org/10.3390/pathogens12010037
Chicago/Turabian StylePei, Dongli, Qingchen Zhang, Xiaoqin Zhu, and Lei Zhang. 2023. "Biological Control of Verticillium Wilt and Growth Promotion in Tomato by Rhizospheric Soil-Derived Bacillus amyloliquefaciens Oj-2.16" Pathogens 12, no. 1: 37. https://doi.org/10.3390/pathogens12010037
APA StylePei, D., Zhang, Q., Zhu, X., & Zhang, L. (2023). Biological Control of Verticillium Wilt and Growth Promotion in Tomato by Rhizospheric Soil-Derived Bacillus amyloliquefaciens Oj-2.16. Pathogens, 12(1), 37. https://doi.org/10.3390/pathogens12010037