
First Molecular Identification of Fasciola gigantica in Slaughtered Cattle in Cape Verde: Prevalence, Gross Pathological Lesions, Genetic Identification and Coprological Analysis



Abstract
:1. Introduction
2. Materials and Methods
3. Results
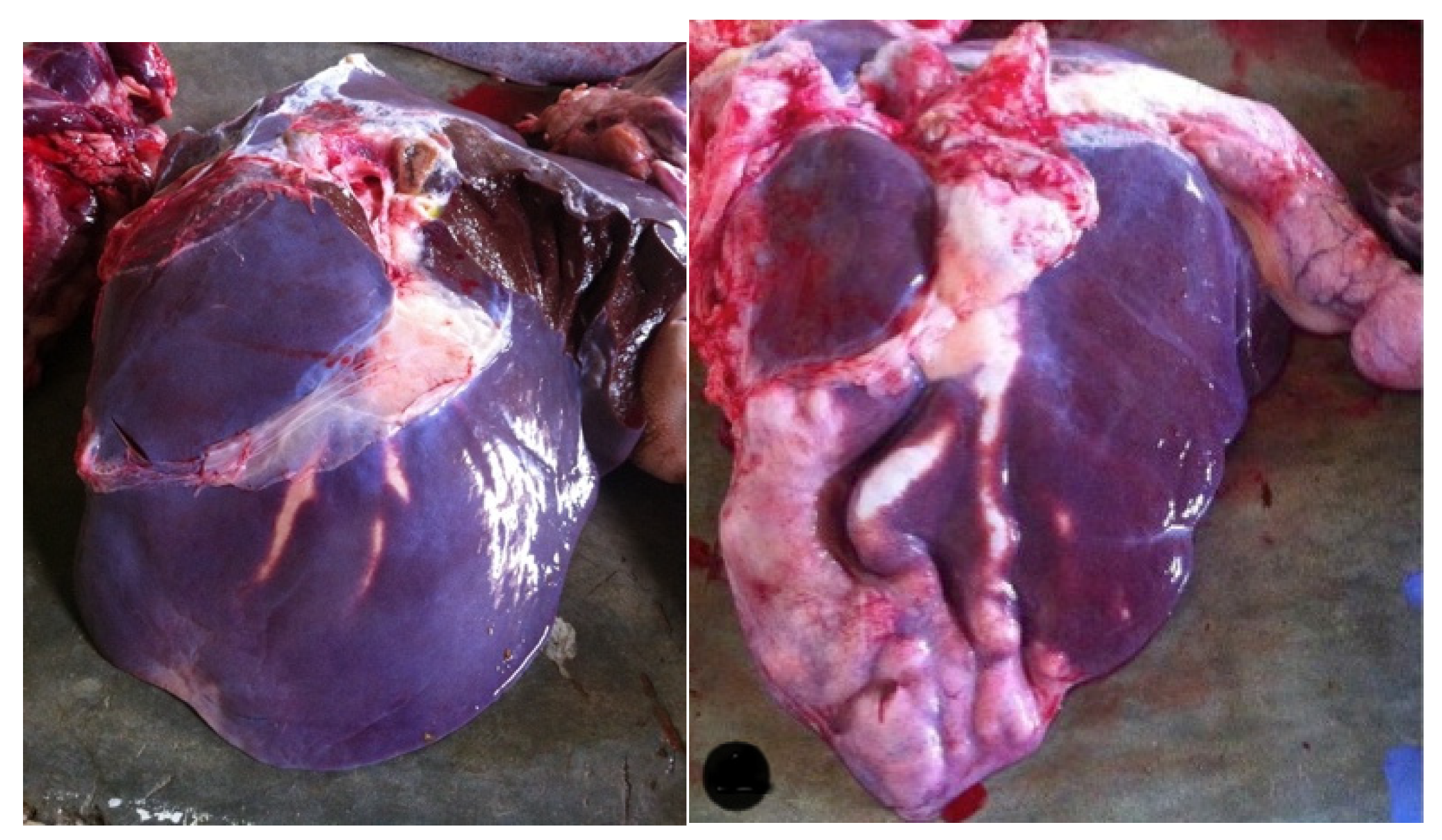
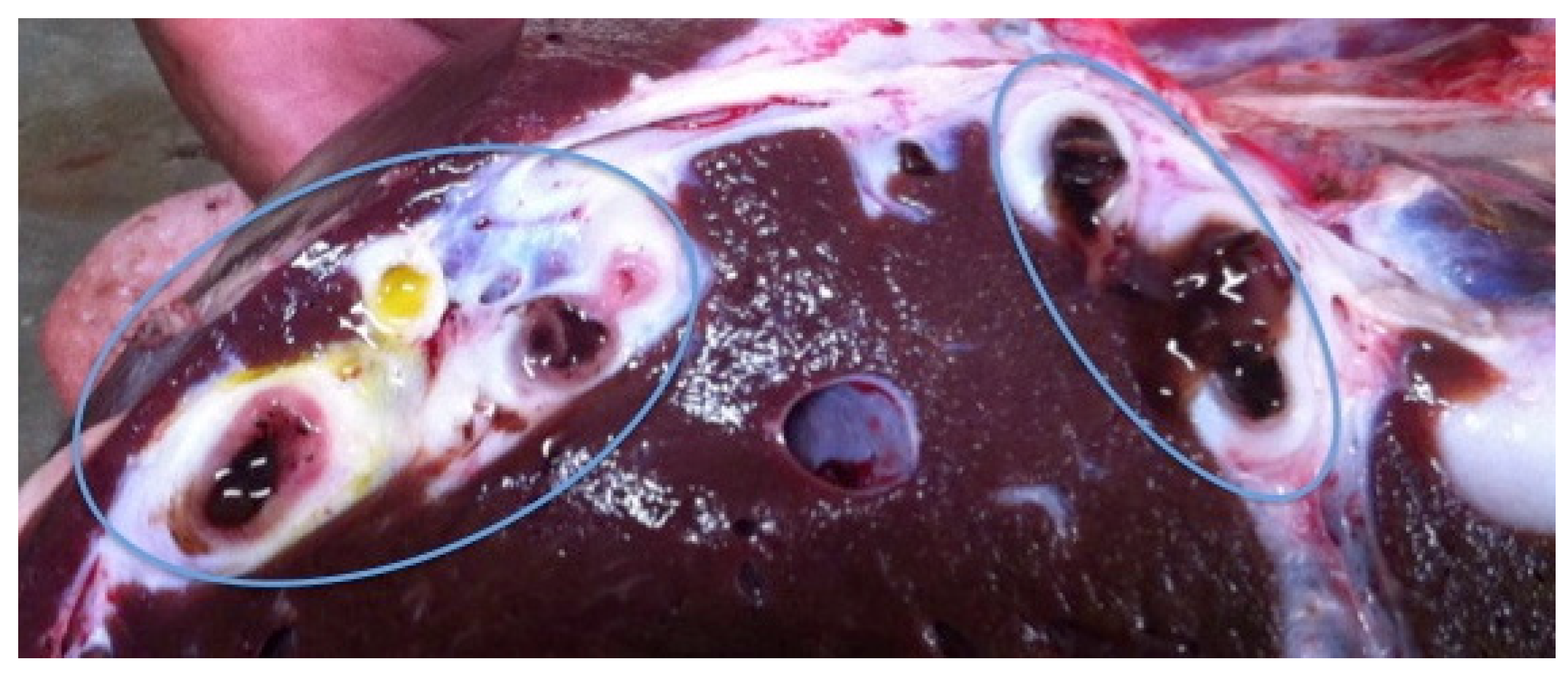
3.1. Prevalence, Gross Pathological Lesions, and Presence of Flukes
3.2. Coprological Analysis
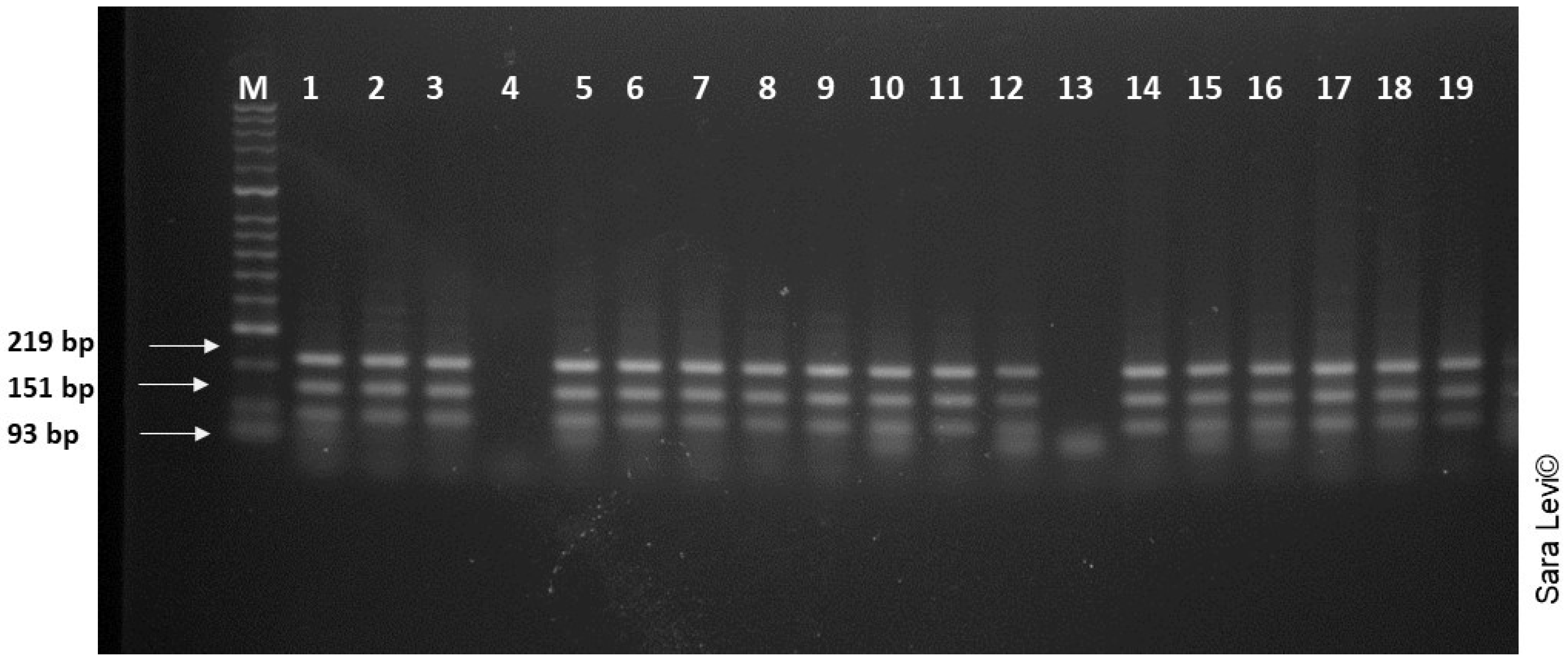
3.3. Genetic Analysis and Species Identification
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Taylor, S.M.; Andrews, A.H. Endoparasites. In Bovine Medicine Diseases and Husbandry of Cattle, 2nd ed.; Andrews, A.H., Ed.; Blackwell Science Ltd.: Oxford, UK, 2004; pp. 267–286. [Google Scholar]
- Mas-Coma, M.S.; Rodriguez, A.; Bargues, M.D.; Valero, M.A.; Coelho, J.R.; Aangles, R. Secondary reservoir role of domestic animals other than sheep and cattle in fasciolosis transmission in the Northern Bolivian Altiplano. Res. Rev. Parasitol. 1997, 57, 39–46. [Google Scholar]
- Mas-Coma, S.; Angles, R.; Esteban, J.G.; Bargues, M.D.; Buchon, P.; Franken, M.; Strauss, W. The Northern Bolivian Altiplano: A region highly endemic for human fasciolosis. Trop. Med. Int. Health 1999, 4, 454–467. [Google Scholar] [CrossRef]
- Bowman, D.D. Georgi’s Parasitology for Veterinarians, 9th ed.; Saunders Elsevier: St. Louis, MO, USA, 2009. [Google Scholar]
- Torgerson, P.R.; Claxton, J. Epidemiology and control. In Fasciolosis, 1st ed.; Dalton, J.P., Ed.; CABI Publishing: Oxford, UK, 1999; pp. 113–149. [Google Scholar]
- Spithill, T.W.; Smooker, P.M.; Copeman, D.B. Fasciola gigantica: Epidemiology, control, immunology and molecular biology. In Fasciolosis, 1st ed.; Dalton, J.P., Ed.; CABI Publishing: Oxford, UK, 1999; pp. 465–525. [Google Scholar]
- Dermauw, V.; Muchai, J.; Al Kappany, Y.; Castaneda, A.L.F.; Dorny, P. Human Fascioliasis in Africa: A systematic review. PLoS ONE 2021, 16, e0261166. [Google Scholar] [CrossRef]
- Nozais, J.P.; Thomas, I.; Bricaire, F.; Danis, M.; Gentilini, M. Apropos of 1 case of hepatogastric fasciolosis in Cape Verde. Bull. Soc. Pathol. Exot. 1998, 91, 148–150. [Google Scholar]
- Almeida, A.P. Produção e Caracterização de Antigénios de Fasciola Gigantica e Sua Avaliação no Imunodiagnóstico de Fasciolosis. Master’s Thesis, Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Lisboa, Portugal, 2010. [Google Scholar]
- Graham, C.S.; Brodie, S.B.; Weller, P.F. Imported Fasciola hepatica infection in the United States and treatment with triclabendazole. Clin. Infect. Dis. 2001, 33, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Corredoura, A.S.; Beato, V.; Moraes, F.; Sousa, S.; Pinheiro, N.; Graça, J.P.; Abecasis, P. Fasciolíase hepática humana tratada com triclabendazol. Med. Interna 2002, 9, 39. [Google Scholar]
- Rosa, F.; Crespo, M.V.; Simões, M.; Évora, I.C.; Moreira, E.; Ferreira, M.L. Epidemiologia e controlo da fasciolosis em Cabo Verde—Estudos entre 1994 e 1999. Acta Parasitol. Port. 2004, 11, 9–15. [Google Scholar]
- Semedo, C.M.A. Fasciolosis no Concelho de São Domingos, Ilha de Santiago, Cabo Verde. Diploma Thesis, Instituto Superior de Educação, Universidade de Cabo Verde, Praia, Cabo Verde, 2006. [Google Scholar]
- Pile, E.A.; Ahid, S.M.M.; Silva, D.N.S. Variação mensal da fasciolíase entre pacientes com distúrbios gastro-hepáticos em Santiago, Cabo Verde. Biológico 2008, 70, 15–18. [Google Scholar]
- Centeio, A.R. Prejuízos Económicos e Proposta de Modelo Estratégico Para o Controle da Fasciolosis Hepática. Diploma Thesis, Centro de Formação Agrária, Instituto Nacional de Investigação e Desenvolvimento Agrário, Universidade de Cabo Verde, São Jorge dos Órgãos, Cabo Verde, 2008. [Google Scholar]
- Matos, L.M.M.; Pile, E.A.; Almada, C.H.B.A.; Lima, H.S.C.; Ahid, S.M.M. Distribuição geopolítica da fasciolosis hepática na ilha de Santiago, Cabo Verde. Arq. Inst. Biol. São Paulo 2011, 78, 435–437. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Funatsu, I.R.; Bargues, M.D. Fasciola hepatica and lymnaeid snails occurring at very high altitude in South America. Parasitology 2001, 123, 115–127. [Google Scholar] [CrossRef]
- Citer, L. Endemic disease monitoring using abattoir surveillance—A pilot project. In Proceedings of the MINTRAC, Meat Inspection and Quality Assurance National Conference, Sidney, Australia, 16–17 October 2008. [Google Scholar]
- Roberts, L.S.; Janovy, J. Gerald, D. Schmidt and Larry, S. Roberts Foundations of Parasitology, 8th ed.; McGraw-Hill: New York, NY, USA, 2008. [Google Scholar]
- Nafarnda, W.D.; Ajayi, I.E.; Shawulu, J.C.; Kawe, M.S.; Omeiza, G.K.; Sani, N.A.; Tenuche, O.Z.; Dantong, D.D.; Tags, S.Z. Bacteriological Quality of Abattoir Effluents Discharged into Water Bodies in Abuja, Nigeria. Int. Sch. Res. Netw. 2012, 2012, 515689. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.R.L.; Paula, V.S.; Oliveira, J.M.; Marchevsky, R.S.; Pinto, M.A. Hepatitis E virus in swine and effluent samples from slaughterhouses in Brazil. Vet. Microbiol. 2011, 149, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Pinto, J.N. Direito à Alimentação e Segurança Alimentar e Nutricional nos Países da CPLP. 2011. Available online: https://www.fao.org/3/i3348e/i3348e.pdf (accessed on 23 November 2022).
- Mascarenhas, D.D.P. Os Desafios da Protecção Social Para Alcançar a Segurança Alimentar e Nutricional. In Proceedings of the Conferência dos Ministros do Trabalho e Segurança Social e dos Assuntos Sociais da CPLP, Maputo, Mozambique, 23–25 April 2013. [Google Scholar]
- Ministério do Desenvolvimento Rural, RGA2004. Available online: https://ine.cv/wp-content/uploads/2016/11/no-de-exploracoes-com-pecuaria-por-ilha-segundo-especie-bovina-2004.pdf (accessed on 23 November 2022).
- Gracey, J.F.; Collins, D.S.; Huey, R.J. Meat Hygiene, 10th ed.; WB Saunders: Loanhead, Scotland, 1999. [Google Scholar]
- Vignau, M.L.; Venturini, L.M.; Romero, J.R.; Eiras, D.F.; Basso, W.U. Parasitología Práctica y Modelos de Enfermedades Parasitarias en los Animales Domésticos, 1st ed.; Facultad de Ciencias Veterinarias, Universidad Nacional de La Plata: La Plata, Argentina, 2005. [Google Scholar]
- Stothard, J.R.; Hughes, S.; Rollinson, D. Variation within the internal transcribed spacer (ITS) of ribosomal DNA genes of intermediate snail hosts within the genus Bulinus (Gastropoda: Planorbidae). Acta Trop. 1996, 61, 19–29. [Google Scholar] [CrossRef]
- Rokni, M.B.; Mirhendi, H.; Mizani, A.; Mohebali, M.; Sharbatkhori, M.; Kia, E.B.; Abdoli, H.; Izadi, S. Identification and differentiation of Fasciola hepatica and Fasciola gigantica using a simple PCR-restriction enzyme method. Exp. Parasitol. 2010, 124, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Matos, L.M.M. Taxa de Ocorrência e Redistribuição Geopolítica da Fasciolose Hepática na Ilha de Santiago, Cabo Verde. Diploma Thesis, Instituto de Investigação e Desenvolvimento Agrário, Centro de Formação Agrária, Universidade de Cabo Verde, São Jorge dos Órgãos, Cabo Verde, 2008. [Google Scholar]
- Amaral, I. Santiago de Cabo Verde, a Terra e os Homens; Junta de Investigação do Ultramar: Lisboa, Portugal, 1964. [Google Scholar]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards). Scientific Opinion on the public health hazards to be covered by inspection of meat (bovine animals). EFSA J. 2013, 11, 3265. [Google Scholar] [CrossRef]
- Fairweather, I. Reducing the future threat from (liver) fluke: Realistic prospect or quixotic fantasy? Vet. Parasitol. 2011, 180, 133–143. [Google Scholar] [CrossRef]
- Norbury, L.J. Structure, Function and Evolutionary Studies of Fasciola Cathepsin L-like Proteases. PhD Thesis, RMIT University, Melbourne, Australia, 2008. [Google Scholar]
- Rapsch, C.; Schweizer, G.; Grimm, F.; Kohler, L.; Bauer, C.; Deplazes, P.; Braun, U.; Torgerson, P.R. Estimating the true prevalence of Fasciola hepatica in cattle slaughtered in Switzerland in the absence of an absolute diagnostic test. Int. J. Parasitol. 2006, 36, 1153–1158. [Google Scholar] [CrossRef]
- Abunna, F.; Asfaw, L.; Megersa, B.; Regassa, A. Bovine fasciolosis: Coprological, abattoir survey and its economic impact due to liver condemnation at Soddo municipal abattoir, Southern Ethiopia. Trop. Anim. Health Prod. 2010, 42, 289–292. [Google Scholar] [CrossRef]
- Chand, M.A.; Herman, J.S.; Partridge, D.G.; Hewitt, K.; Chiodini, P.L. Imported Human Fascioliasis, United Kingdom. Emerg. Infect. Dis. 2009, 15, 1876–1877. [Google Scholar] [CrossRef]
- Hashimoto, K.; Watanobe, T.; Liu, C.X.; Iinit, I.; Blair, D.; Onishi, S.; Agatsuma, T. Mitochondrial DNA and nuclear DNA indicate that the Japanese Fasciola species is F. gigantica. Parasitol. Res. 1997, 83, 220–225. [Google Scholar] [CrossRef]
- Saijuntha, W.; Tantrawatpan, C.; Agatsuma, T.; Wang, C.; Intapan, P.M.; Maleewong, W.; Petney, T.N. Revealing genetic hybridization and DNA recombination of Fasciola hepatica and Fasciola gigantica in nuclear introns of the hybrid Fasciola flukes. Mol. Biochem. Parasitol. 2018, 223, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Agatsuma, T.; Arakawa, Y.; Iwagami, M.; Honzako, Y.; Cahyaningsih, U.; Kang, S.Y.; Hong, S.J. Molecular evidence of natural hybridization between Fasciola hepatica and F. gigantica. Parasitol. Int. 2000, 49, 231–238. [Google Scholar] [CrossRef]
- Huang, W.Y.; He, B.; Wang, C.R.; Zhu, X.Q. Characterisation of Fasciola species from Mainland China by ITS-2 ribosomal DNA sequence. Vet. Parasitol. 2004, 120, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; De, N.V.; Agatsuma, T.; Thi Nguyen, T.G.; Nguyen, Q.D.; McManus, D.P.; Blair, D. Human fasciolosis and the presence of hybrid/introgressed forms of Fasciola hepatica and Fasciola gigantica in Vietnam. Int. J. Parasitol. 2008, 38, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.; Dar, Y.; Ichikawa, M.; Fukuda, Y.; Tada, C.; Itagaki, T.; Nakai, Y. Identification of Fasciola species isolated from Egypt based on sequence analysis of genomic (ITS1 and ITS2) and mitochondrial (NDI and COI) gene markers. Parasitol. Int. 2011, 60, 5–12. [Google Scholar] [CrossRef]
- Evack, J.G.; Schmidt, R.S.; Boltryk, S.D.; Voss, T.S.; Batil, A.A.; Ngandolo, B.N.; Greter, H.; Utzinger, J.; Zinsstag, J.; Balmer, O. Molecular Confirmation of a Fasciola Gigantica X Fasciola Hepatica Hybrid in a Chadian Bovine. J. Parasitol. 2020, 10, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Hasanpour, H.; Falak, R.; Naddaf, S.R.; Mas-Coma, S.; Rokni, M.B.; Badirzadeh, A.; Mokhtarian, K.; Mohebali, M.; Jafarpour Azami, S.; Fadavi, A.; et al. Molecular Characterization of Fasciola spp. from Some Parts of Iran. Iran J. Public Health 2020, 49, 157–166. [Google Scholar] [CrossRef]
- Sanchez-Vazquez, M.J.; Lewis, F.I. Investigating the impact of fasciolosis on cattle carcass performance. Vet. Parasitol. 2013, 193, 307–311. [Google Scholar] [CrossRef]
- Scott, P. Liver Fluke Control in Cattle. NADIS Health Bulletin. 2012. Available online: https://www.nadis.org.uk/disease-a-z/cattle/endoparasites-in-cattle/liver-fluke-control-in-cattle/ (accessed on 2 January 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FCL Case Number | Liver Analysis (Presence of Flukes) | Fecal Analysis (Presence of Fasciola spp. Eggs) |
|---|---|---|
| 1 | 1 | - |
| 2 | 1 | 1 |
| 3 | 1 | 1 |
| 4 | 1 | 0 |
| 5 | 1 | 1 |
| 6 | 1 | 1 |
| 7 | 0 | 0 |
| 8 | 1 | 0 |
| 9 | 1 | 1 |
| 10 | 1 | 0 |
| 11 | 1 | 1 |
| 12 | 1 | 1 |
| Total | 11 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levy, S.; Calado, M.; Mateus, T.L.; Vieira-Pinto, M. First Molecular Identification of Fasciola gigantica in Slaughtered Cattle in Cape Verde: Prevalence, Gross Pathological Lesions, Genetic Identification and Coprological Analysis. Pathogens 2023, 12, 75. https://doi.org/10.3390/pathogens12010075
Levy S, Calado M, Mateus TL, Vieira-Pinto M. First Molecular Identification of Fasciola gigantica in Slaughtered Cattle in Cape Verde: Prevalence, Gross Pathological Lesions, Genetic Identification and Coprological Analysis. Pathogens. 2023; 12(1):75. https://doi.org/10.3390/pathogens12010075
Chicago/Turabian StyleLevy, Sara, Manuela Calado, Teresa Letra Mateus, and Madalena Vieira-Pinto. 2023. "First Molecular Identification of Fasciola gigantica in Slaughtered Cattle in Cape Verde: Prevalence, Gross Pathological Lesions, Genetic Identification and Coprological Analysis" Pathogens 12, no. 1: 75. https://doi.org/10.3390/pathogens12010075
APA StyleLevy, S., Calado, M., Mateus, T. L., & Vieira-Pinto, M. (2023). First Molecular Identification of Fasciola gigantica in Slaughtered Cattle in Cape Verde: Prevalence, Gross Pathological Lesions, Genetic Identification and Coprological Analysis. Pathogens, 12(1), 75. https://doi.org/10.3390/pathogens12010075