
Influence of Maternal BLV Infection on miRNA and tRF Expression in Calves



Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. White Blood Cell Preservation
2.3. BLV Antibody and BLV PVL Determination
2.4. RNA Extraction, Library Preparation, and Sequencing
2.5. Small RNA Processing
2.6. Differential Expression Analysis
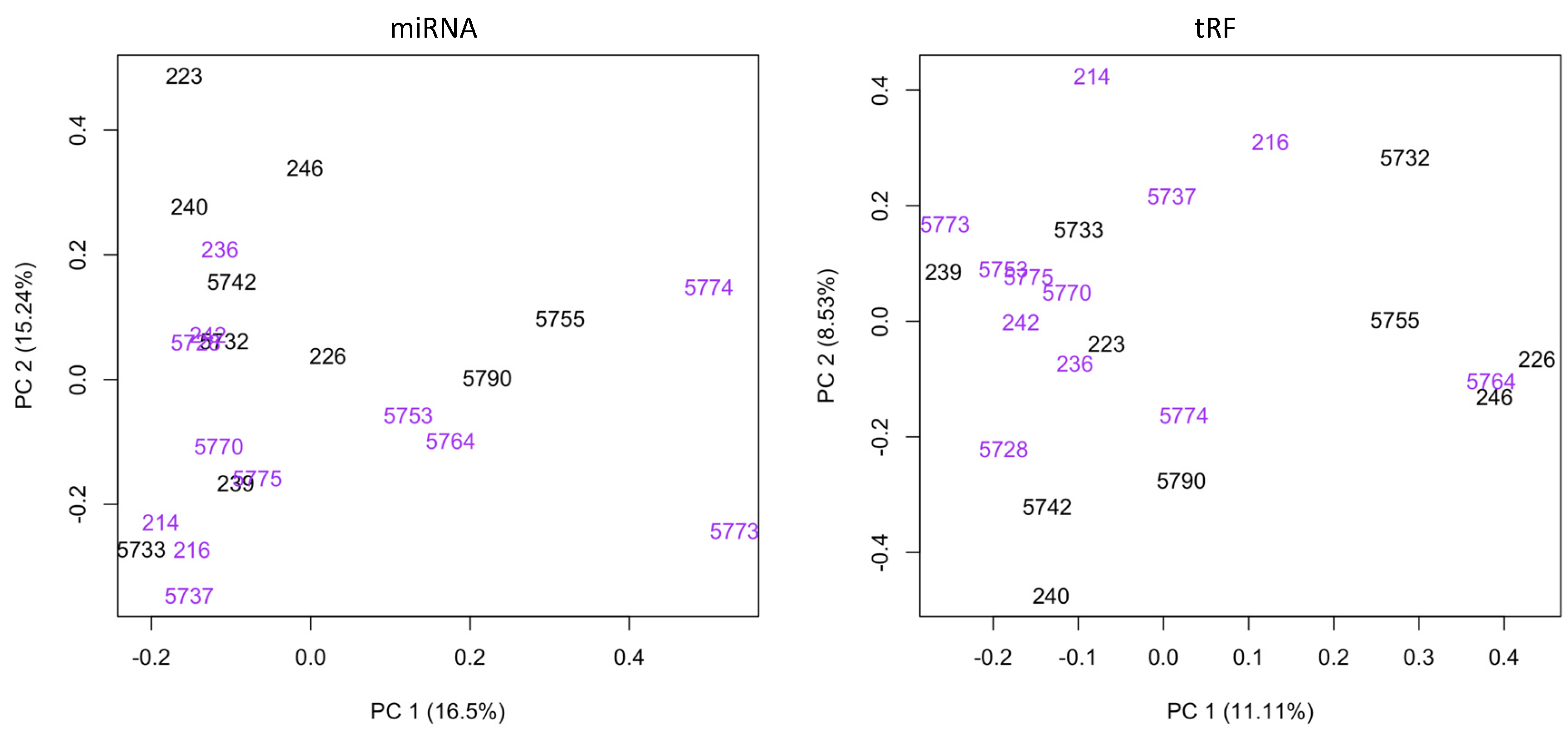
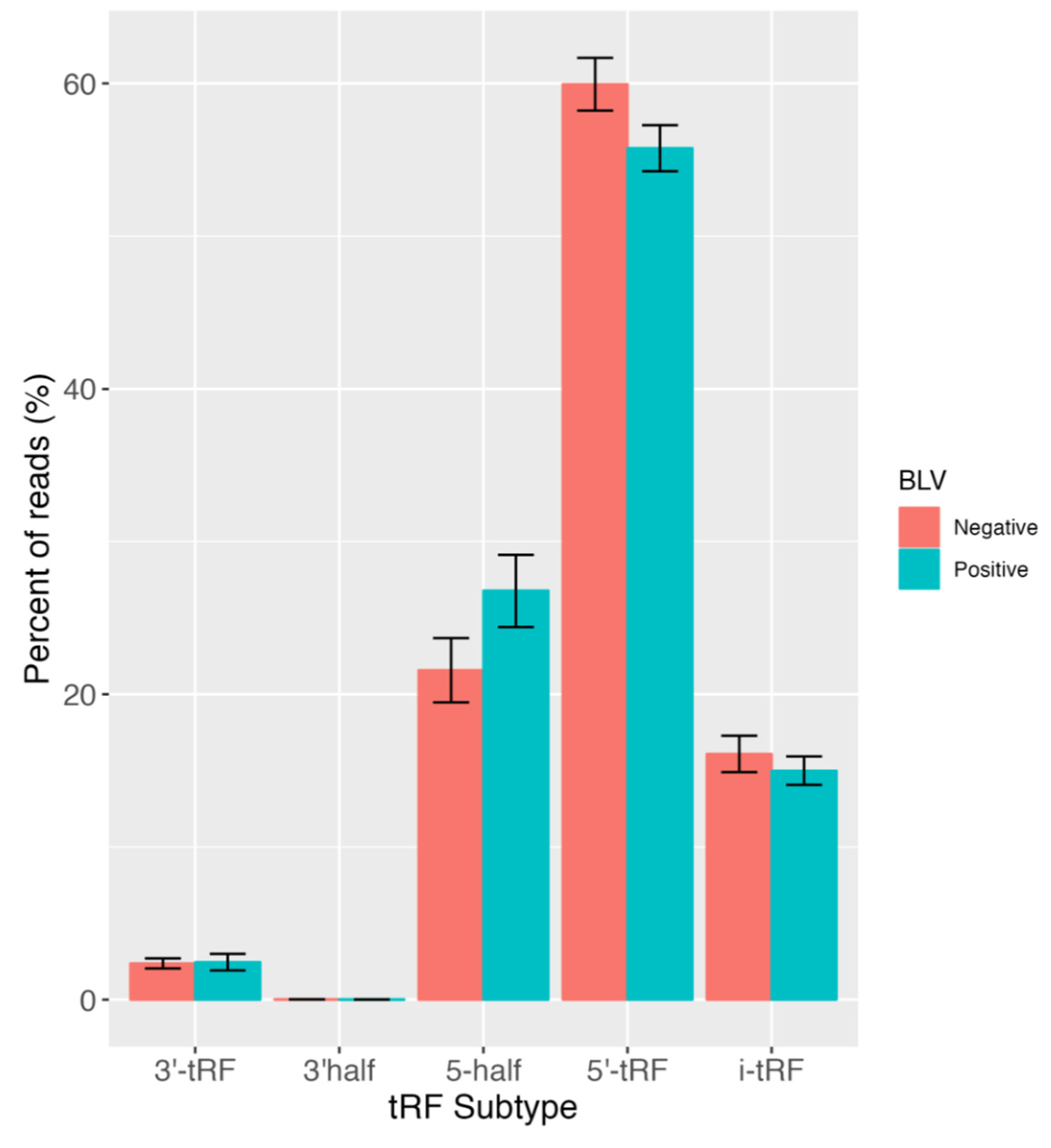
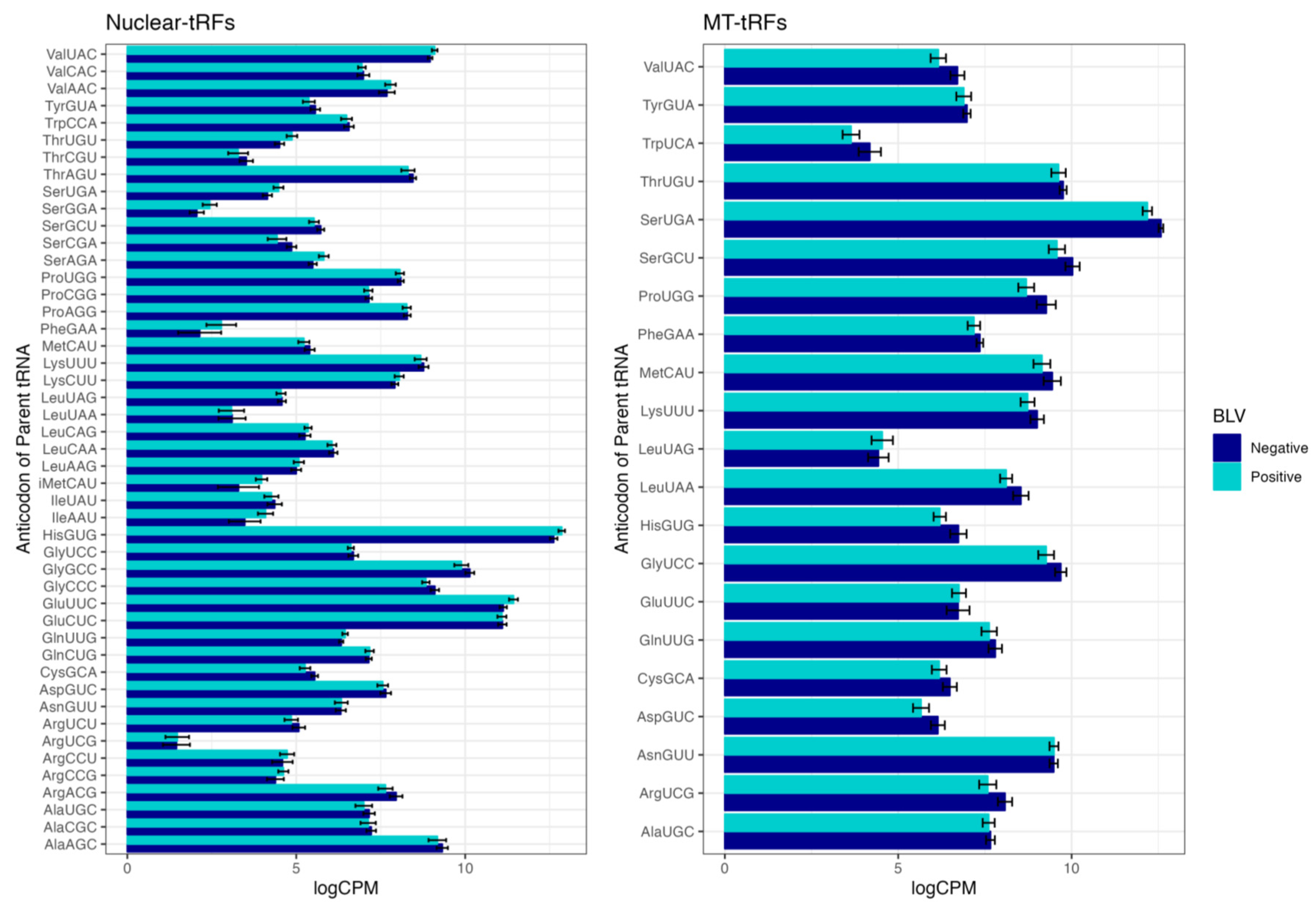
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laporta, J.; Ferreira, F.C.; Ouellet, V.; Dado-Senn, B.; Almeida, A.K.; De Vries, A.; Dahl, G.E. Late-gestation heat stress impairs daughter and granddaughter lifetime performance. J. Dairy Sci. 2020, 103, 7555–7568. [Google Scholar] [CrossRef] [PubMed]
- Moriel, P.; Piccolo, M.B.; Artioli, L.F.; Marques, R.S.; Poore, M.H.; Cooke, R.F. Short-term energy restriction during late gestation of beef cows decreases postweaning calf humoral immune response to vaccination. J. Anim. Sci. 2016, 94, 2542–2552. [Google Scholar] [CrossRef] [PubMed]
- Alfaradhi, M.Z.; Ozanne, S.E. Developmental programming in response to maternal overnutrition. Front. Genet. 2011, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, D.; St-Amour, I.; Cisbani, G.; Rousseau, L.S.; Cicchetti, F. The different effects of LPS and poly I:C prenatal immune challenges on the behavior, development and inflammatory responses in pregnant mice and their offspring. Brain Behav. Immun. 2014, 38, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Legoff, L.; D’Cruz, S.C.; Tevosian, S.; Primig, M.; Smagulova, F. Transgenerational Inheritance of Environmentally Induced Epigenetic Alterations during Mammalian Development. Cells 2019, 8, 1559. [Google Scholar] [CrossRef] [PubMed]
- Dykxhoorn, D.M.; Novina, C.D.; Sharp, P.A. Killing the messenger: Short RNAs that silence gene expression. Nat. Rev. Mol. Cell Biol. 2003, 4, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; Van Dongen, S.; Inoue, K.; Enright, A.J.; Schier, A.F. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Gu, J.; Wang, M.; Zhang, J.; Chen, Y.; Jiang, P.; Zhu, T.; Zhang, X. Emerging roles of tRNA-derived fragments in cancer. Mol. Cancer 2023, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Xie, Y.; Zhang, S.; Song, X.; Xiao, B.; Yan, Z. tRNA-derived fragments: Mechanisms underlying their regulation of gene expression and potential applications as therapeutic targets in cancers and virus infections. Theranostics 2021, 11, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes. Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, M.; Honda, S.; Kirino, Y. Transfer RNA as a source of small functional RNA. J. Mol. Biol. Mol. Imaging 2014, 1, 8. [Google Scholar]
- Shigematsu, M.; Kirino, Y. tRNA-Derived Short Non-coding RNA as Interacting Partners of Argonaute Proteins. Gene Regul. Syst. Bio 2015, 9, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.; Mouillet, J.F.; Mishima, T.; Chu, T.; Sadovsky, E.; Coyne, C.B.; Parks, W.T.; Surti, U.; Sadovsky, Y. Expression and trafficking of placental microRNAs at the feto-maternal interface. FASEB J. 2017, 31, 2760–2770. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Tsui, N.B.; Lo, Y.M. Placental RNA in maternal plasma: Toward noninvasive fetal gene expression profiling. Ann. N. Y. Acad. Sci. 2006, 1075, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Lippolis, J.D.; Casas, E. Expression Profiles and Interaction of MicroRNA and Transcripts in Response to Bovine Leukemia Virus Exposure. Front. Vet. Sci. 2022, 9, 887560. [Google Scholar] [CrossRef] [PubMed]
- Taxis, T.M.; Kehrli, M.E., Jr.; D’Orey-Branco, R.; Casas, E. Association of Transfer RNA Fragments in White Blood Cells With Antibody Response to Bovine Leukemia Virus in Holstein Cattle. Front. Genet. 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Ma, H.; Lippolis, J.D. Expression of Viral microRNAs in Serum and White Blood Cells of Cows Exposed to Bovine Leukemia Virus. Front. Vet. Sci. 2020, 7, 536390. [Google Scholar] [CrossRef] [PubMed]
- Aida, Y.; Murakami, H.; Takahashi, M.; Takeshima, S.N. Mechanisms of pathogenesis induced by bovine leukemia virus as a model for human T-cell leukemia virus. Front. Microbiol. 2013, 4, 328. [Google Scholar] [CrossRef] [PubMed]
- Bendixen, H.J. Epidemiological studies of bovine leukosis in Denmark. Proc. R. Soc. Med. 1966, 59, 657–660. [Google Scholar] [PubMed]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Herd-level determinants of bovine leukaemia virus prevalence in dairy farms. J. Dairy Res. 2012, 79, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Boes, K.M.; Durham, A.C. Bone Marrow, Blood Cells, and the Lymphoid/Lymphatic System. Pathol. Basis Vet. Dis. 2017, 6, 724–804.e2. [Google Scholar] [CrossRef]
- Mancuso, S.; Mattana, M.; Carlisi, M.; Santoro, M.; Siragusa, S. Effects of B-Cell Lymphoma on the Immune System and Immune Recovery after Treatment: The Paradigm of Targeted Therapy. Int. J. Mol. Sci. 2022, 23, 3368. [Google Scholar] [CrossRef] [PubMed]
- White, T.L.; Moore, D.A. Reasons for whole carcass condemnations of cattle in the United States and implications for producer education and veterinary intervention. J. Am. Vet. Med. Assoc. 2009, 235, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Rezac, D.J.; Thomson, D.U.; Siemens, M.G.; Prouty, F.L.; Reinhardt, C.D.; Bartle, S.J. A survey of gross pathologic conditions in cull cows at slaughter in the Great Lakes region of the United States. J. Dairy Sci. 2014, 97, 4227–4235. [Google Scholar] [CrossRef] [PubMed]
- Panei, C.J.; Takeshima, S.N.; Omori, T.; Nunoya, T.; Davis, W.C.; Ishizaki, H.; Matoba, K.; Aida, Y. Estimation of bovine leukemia virus (BLV) proviral load harbored by lymphocyte subpopulations in BLV-infected cattle at the subclinical stage of enzootic bovine leucosis using BLV-CoCoMo-qPCR. BMC Vet. Res. 2013, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.H.; Lucas, M.H.; Swallow, C. Comparison of the agar gel immunodiffusion test and ELISA in the detection of bovine leukosis virus antibody in cattle persistently infected with bovine virus diarrhoea virus. Vet. Immunol. Immunopathol. 1989, 22, 275–281. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture. Bovine Leukosis Virus (BLV) on U.S. Dairy Operations; Animal and Plant Health Inspection Service: Washington, DC, USA, 2008.
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J. Dairy. Sci. 2012, 95, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Bautista, C.T.; Sanchez, J.L.; Montano, S.M.; Laguna-Torres, A.; Suarez, L.; Sanchez, J.; Campos, P.; Gallardo, C.; Mosquera, C.; Villafane, M.; et al. Seroprevalence of and risk factors for HIV-1 infection among female commercial sex workers in South America. Sex. Transm. Infect. 2006, 82, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Morimoto, T.; Shimanoe, C.; Ono, R.; Otani, K.; Mawatari, M. Risk factors for progression of the severity of locomotive syndrome: A two-year longitudinal observational study. J. Orthop. Sci. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, H.C.; Norby, B.; Droscha, C.J.; Sordillo, L.M.; Coussens, P.M.; Bartlett, P.C. Bovine leukemia virus detection and dynamics following experimental inoculation. Res. Vet. Sci. 2020, 133, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Pavliscak, L.A.; Nirmala, J.; Singh, V.K.; Sporer, K.R.B.; Taxis, T.M.; Kumar, P.; Goyal, S.M.; Mor, S.K.; Schroeder, D.C.; Wells, S.J.; et al. Tracing Viral Transmission and Evolution of Bovine Leukemia Virus through Long Read Oxford Nanopore Sequencing of the Proviral Genome. Pathogens 2021, 10, 1191. [Google Scholar] [CrossRef]
- Lohr, C.E.; Sporer, K.R.B.; Brigham, K.A.; Pavliscak, L.A.; Mason, M.M.; Borgman, A.; Ruggiero, V.J.; Taxis, T.M.; Bartlett, P.C.; Droscha, C.J. Phenotypic Selection of Dairy Cattle Infected with Bovine Leukemia Virus Demonstrates Immunogenetic Resilience through NGS-Based Genotyping of BoLA MHC Class II Genes. Pathogens 2022, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinf. 2010, 11, 485. [Google Scholar] [CrossRef]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Tseng, E.; Rowan, T.N.; Low, W.Y.; Zimin, A.; Couldrey, C.; et al. De novo assembly of the cattle reference genome with single-molecule sequencing. GigaScience 2020, 9, giaa021. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
- Risso, D.; Schwartz, K.; Sherlock, G.; Dudoit, S. GC-content normalization for RNA-Seq data. BMC Bioinf. 2011, 12, 480. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Korsmo, H.W.; Dave, B.; Trasino, S.; Saxena, A.; Liu, J.; Caviglia, J.M.; Edwards, K.; Dembitzer, M.; Sheeraz, S.; Khaldi, S.; et al. Maternal Choline Supplementation and High-Fat Feeding Interact to Influence DNA Methylation in Offspring in a Time-Specific Manner. Front. Nutr. 2022, 9, 841787. [Google Scholar] [CrossRef]
- Haas, J.D.; Nistala, K.; Petermann, F.; Saran, N.; Chennupati, V.; Schmitz, S.; Korn, T.; Wedderburn, L.R.; Forster, R.; Krueger, A.; et al. Expression of miRNAs miR-133b and miR-206 in the Il17a/f locus is co-regulated with IL-17 production in alphabeta and gammadelta T cells. PLoS ONE 2011, 6, e20171. [Google Scholar] [CrossRef]
- Worley, J.N.; Flores, K.A.; Yang, X.; Chase, J.A.; Cao, G.; Tang, S.; Meng, J.; Atwill, E.R. Prevalence and Genomic Characterization of Escherichia coli O157:H7 in Cow-Calf Herds throughout California. Appl. Environ. Microbiol. 2017, 83, e00734-17. [Google Scholar] [CrossRef]
- Viidu, D.A.; Motus, K. Implementation of a pre-calving vaccination programme against rotavirus, coronavirus and enterotoxigenic Escherichia coli (F5) and association with dairy calf survival. BMC Vet. Res. 2022, 18, 59. [Google Scholar] [CrossRef]
- Koutsoulidou, A.; Mastroyiannopoulos, N.P.; Furling, D.; Uney, J.B.; Phylactou, L.A. Expression of miR-1, miR-133a, miR-133b and miR-206 increases during development of human skeletal muscle. BMC Dev. Biol. 2011, 11, 34. [Google Scholar] [CrossRef]
- Jacometo, C.B.; Alharthi, A.S.; Zhou, Z.; Luchini, D.; Loor, J.J. Maternal supply of methionine during late pregnancy is associated with changes in immune function and abundance of microRNA and mRNA in Holstein calf polymorphonuclear leukocytes. J. Dairy. Sci. 2018, 101, 8146–8158. [Google Scholar] [CrossRef]
- Do, D.N.; Dudemaine, P.L.; Fomenky, B.E.; Ibeagha-Awemu, E.M. Integration of miRNA and mRNA Co-Expression Reveals Potential Regulatory Roles of miRNAs in Developmental and Immunological Processes in Calf Ileum during Early Growth. Cells 2018, 7, 134. [Google Scholar] [CrossRef]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef]
- Wystub, K.; Besser, J.; Bachmann, A.; Boettger, T.; Braun, T. miR-1/133a clusters cooperatively specify the cardiomyogenic lineage by adjustment of myocardin levels during embryonic heart development. PLoS Genet. 2013, 9, e1003793. [Google Scholar] [CrossRef]
- Xu, P.; Wu, Q.; Yu, J.; Rao, Y.; Kou, Z.; Fang, G.; Shi, X.; Liu, W.; Han, H. A Systematic Way to Infer the Regulation Relations of miRNAs on Target Genes and Critical miRNAs in Cancers. Front. Genet. 2020, 11, 278. [Google Scholar] [CrossRef]
- Kuscu, C.; Kumar, P.; Kiran, M.; Su, Z.; Malik, A.; Dutta, A. tRNA fragments (tRFs) guide Ago to regulate gene expression post-transcriptionally in a Dicer-independent manner. RNA 2018, 24, 1093–1105. [Google Scholar] [CrossRef]
- Ying, S.Y.; Chang, D.C.; Lin, S.L. The microRNA (miRNA): Overview of the RNA genes that modulate gene function. Mol. Biotechnol. 2008, 38, 257–268. [Google Scholar] [CrossRef]
- Olvedy, M.; Scaravilli, M.; Hoogstrate, Y.; Visakorpi, T.; Jenster, G.; Martens-Uzunova, E.S. A comprehensive repertoire of tRNA-derived fragments in prostate cancer. Oncotarget 2016, 7, 24766–24777. [Google Scholar] [CrossRef]
- Krishna, S.; Yim, D.G.; Lakshmanan, V.; Tirumalai, V.; Koh, J.L.; Park, J.E.; Cheong, J.K.; Low, J.L.; Lim, M.J.; Sze, S.K.; et al. Dynamic expression of tRNA-derived small RNAs define cellular states. EMBO Rep. 2019, 20, e47789. [Google Scholar] [CrossRef]
- Goldkamp, A.K.; Li, Y.; Rivera, R.M.; Hagen, D.E. Differentially expressed tRNA-derived fragments in bovine fetuses with assisted reproduction induced congenital overgrowth syndrome. Front. Genet. 2022, 13, 1055343. [Google Scholar] [CrossRef]
- Kim, D.; Grun, D.; van Oudenaarden, A. Dampening of expression oscillations by synchronous regulation of a microRNA and its target. Nat. Genet. 2013, 45, 1337–1344. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, J.; He, Z.; Fan, X.; Mao, X.; Yang, M.; Yang, D. tRNA-derived fragments as New Hallmarks of Aging and Age-related Diseases. Aging Dis. 2021, 12, 1304–1322. [Google Scholar] [CrossRef]
- Gong, M.; Deng, Y.; Xiang, Y.; Ye, D. The role and mechanism of action of tRNA-derived fragments in the diagnosis and treatment of malignant tumors. Cell Commun. Signal. CCS 2023, 21, 62. [Google Scholar] [CrossRef]
- Raisch, J.; Darfeuille-Michaud, A.; Nguyen, H.T. Role of microRNAs in the immune system, inflammation and cancer. World J. Gastroenterol. 2013, 19, 2985–2996. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Calf ID | Sex 1 | Calf ELISA OD 2 | Dam BLV Status 3 |
|---|---|---|---|
| 1 | M | 2.507 | Negative |
| 2 | F | 2.253 | Negative |
| 3 | M | 2.441 | Negative |
| 4 | F | 2.585 | Negative |
| 5 | F | 1.246 | Negative |
| 6 | M | 2.461 | Negative |
| 7 | F | 1.952 | Negative |
| 8 | F | 2.257 | Negative |
| 9 | M | 2.756 | Negative |
| 10 | M | 2.765 | Negative |
| 11 | F | 2.061 | Positive |
| 12 | F | 2.622 | Positive |
| 13 | M | 3.093 | Positive |
| 14 | M | 1.611 | Positive |
| 15 | F | 2.720 | Positive |
| 16 | F | 2.820 | Positive |
| 17 | F | 2.596 | Positive |
| 18 | F | 2.325 | Positive |
| 19 | M | 2.232 | Positive |
| 20 | F | 1.927 | Positive |
| 21 | F | 2.052 | Positive |
| 22 | M | 2.909 | Positive |
| miRNAs/tRFs | logFC 1 | logCPM 2 | LR 3 | p-Value | FDR 4 |
|---|---|---|---|---|---|
| bta-miR-1 | −6.835 | 5.777 | 31.975 | 1.56 × 10−8 | 1.61 × 10−5 |
| bta-miR-206 | −6.072 | 4.258 | 28.535 | 9.20 × 10−8 | 4.74 × 10−5 |
| bta-miR-133a | −4.401 | 3.196 | 20.003 | 7.73 × 10−6 | 2.65 × 10−3 |
| bta-miR-133b | −4.653 | 0.494 | 16.522 | 4.81 × 10−5 | 1.24 × 10−2 |
| bta-miR-2450d | 4.102 | −3.528 | 14.012 | 1.82 × 10−4 | 3.74 × 10−2 |
| tRF-27-W4R951KHZKK | 2.874 | 8.612 | 27.967 | 1.23 × 10−7 | 2.11 × 10−3 |
| tRF-36-8JZ8RN58X2NF79E | −4.028 | 4.762 | 20.405 | 6.27 × 10−6 | 4.81 × 10−2 |
| tRF-20-0PF05B2I | −3.743 | 4.868 | 19.300 | 1.12 × 10−5 | 4.81 × 10−2 |
| tRF-22-S3M8309NF | 4.074 | 5.061 | 19.040 | 1.28 × 10−5 | 4.81 × 10−2 |
| tRF-26-M87SFR2W9J0 | 3.269 | 5.613 | 18.861 | 1.41 × 10−5 | 4.81 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldkamp, A.K.; Lahuis, C.H.; Hagen, D.E.; Taxis, T.M. Influence of Maternal BLV Infection on miRNA and tRF Expression in Calves. Pathogens 2023, 12, 1312. https://doi.org/10.3390/pathogens12111312
Goldkamp AK, Lahuis CH, Hagen DE, Taxis TM. Influence of Maternal BLV Infection on miRNA and tRF Expression in Calves. Pathogens. 2023; 12(11):1312. https://doi.org/10.3390/pathogens12111312
Chicago/Turabian StyleGoldkamp, Anna K., Ciarra H. Lahuis, Darren E. Hagen, and Tasia M. Taxis. 2023. "Influence of Maternal BLV Infection on miRNA and tRF Expression in Calves" Pathogens 12, no. 11: 1312. https://doi.org/10.3390/pathogens12111312
APA StyleGoldkamp, A. K., Lahuis, C. H., Hagen, D. E., & Taxis, T. M. (2023). Influence of Maternal BLV Infection on miRNA and tRF Expression in Calves. Pathogens, 12(11), 1312. https://doi.org/10.3390/pathogens12111312