
Pathogen-Mediated Alterations of Insect Chemical Communication: From Pheromones to Behavior




Abstract
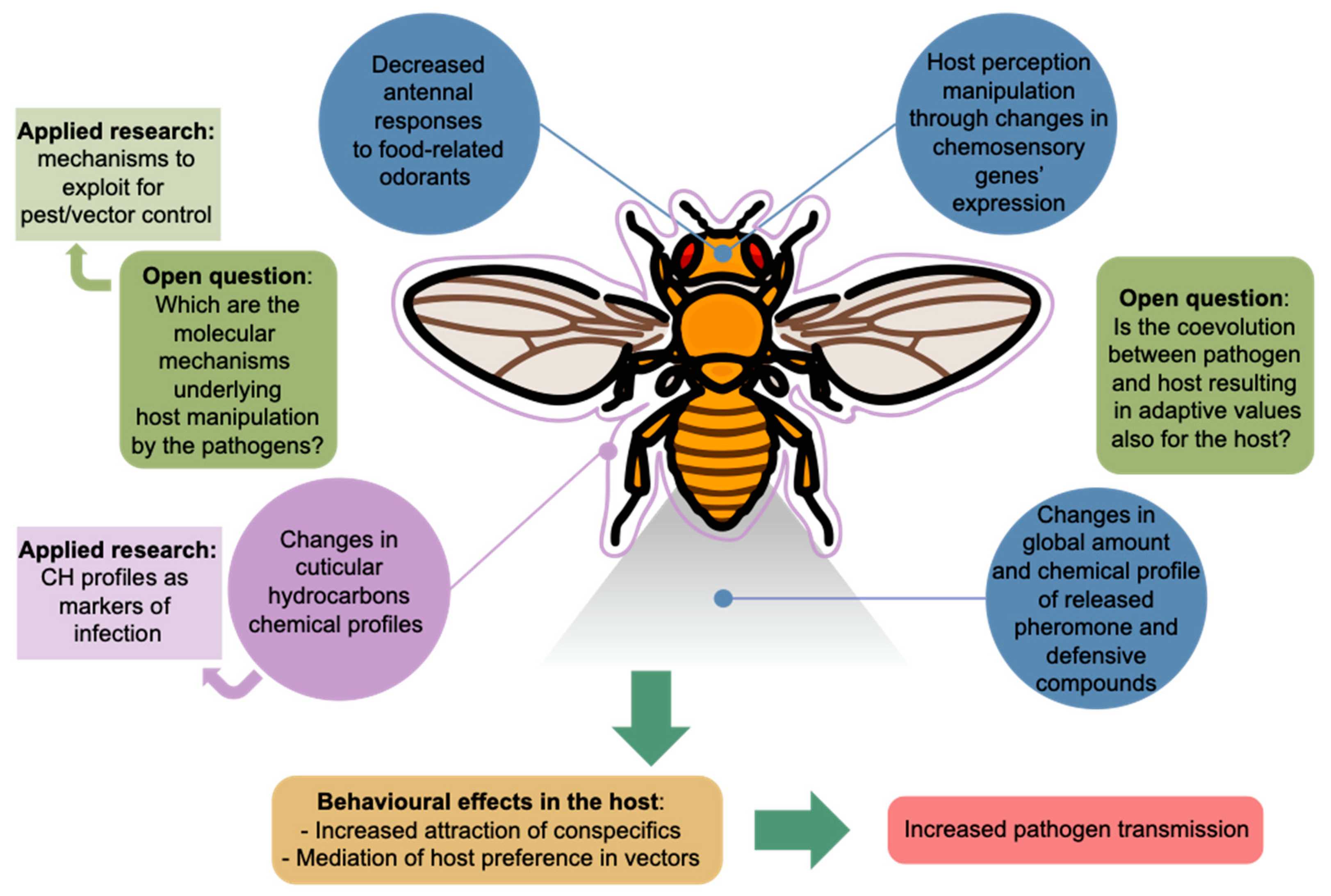
:1. Introduction
2. Bacteria
3. Viruses
4. Fungi
5. Eukaryotes
5.1. Phylum Apicomplexa
5.2. Phylum Euglenozoa
5.3. Phylum Platyhelminthes
5.4. Phylum Nematoda
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ingram, W.M.; Goodrich, L.M.; Robey, E.A.; Eisen, M.B. Mice infected with low-virulence strains of Toxoplasma gondii lose their innate aversion to cat urine, even after extensive parasite clearance. PLoS ONE 2013, 8, e75246. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Lunde, K.B.; Thurman, E.M.; Ritchie, E.G.; Wray, S.N.; Sutherland, D.R.; Kapfer, J.M.; Frest, T.J.; Bowerman, J.; Blaustein, A.R. Parasite (Ribeiroia ondatrae) infection linked to amphibian malformations in the western United States. Ecol. Monogr. 2002, 72, 151–168. [Google Scholar] [CrossRef]
- Libersat, F.; Delago, A.; Gal, R. Manipulation of host behavior by parasitic insects and insect parasites. Annu. Rev. Entomol. 2009, 54, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Spagnoli, S.; Sanders, J.; Kent, M.L. The common neural parasite Pseudoloma neurophilia causes altered shoaling behaviour in adult laboratory zebrafish (Danio rerio) and its implications for neurobehavioural research. J. Fish Dis. 2017, 40, 443–446. [Google Scholar] [CrossRef]
- Sanchez, M.I.; Ponton, F.; Schmidt-Rhaesa, A.; Hughes, D.P.; Misse, D.; Thomas, F. Two steps to suicide in crickets harbouring hairworms. Anim. Behav. 2008, 76, 1621–1624. [Google Scholar] [CrossRef]
- Hughes, D.P.; Wappler, T.; Labandeira, C.C. Ancient death-grip leaf scars reveal ant-fungal parasitism. Biol. Lett. 2011, 7, 67–70. [Google Scholar] [CrossRef]
- Wesołowska, W.; Wesołowski, T. Do Leucochloridium sporocysts manipulate the behaviour of their snail hosts? J. Zool. 2014, 292, 151–155. [Google Scholar] [CrossRef]
- Burand, J.P.; Tan, W.; Kim, W.; Nojima, S.; Roelofs, W. Infection with the insect virus Hz-2v alters mating behavior and pheromone production in female Helicoverpa zea moths. J. Insect Sci. 2005, 5, 6. [Google Scholar] [CrossRef]
- Orlovskis, Z.; Canale, M.C.; Thole, V.; Pecher, P.; Lopes, J.R.S.; Hogenhout, S.A. Insect-borne plant pathogenic bacteria: Getting a ride goes beyond physical contact. Curr. Opin. Insect Sci. 2015, 9, 16–23. [Google Scholar] [CrossRef]
- Mann, R.S.; Ali, J.G.; Hermann, S.L.; Tiwari, S.; Pelz-Stelinski, K.S.; Alborn, H.T.; Stelinski, L.L. Induced release of a plant-defense volatile “deceptively” attracts insect vectors to plants infected with a bacterial pathogen. PLoS Pathog. 2012, 8, e1002610. [Google Scholar] [CrossRef]
- Cui, H.; Wang, Y.; Xue, L.; Chu, J.; Yan, C.; Fu, J.; Chen, M.; Innes, R.W.; Zhou, J.-M. Pseudomonas syringae effector protein AvrB perturbs Arabidopsis hormone signaling by activating MAP kinase 4. Cell Host Microbe 2010, 7, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Orlovskis, Z.; Hogenhout, S.A. A bacterial parasite effector mediates insect vector attraction in host plants independently of developmental changes. Front. Plant Sci. 2016, 7, 885. [Google Scholar] [CrossRef] [PubMed]
- Berdoy, M.; Webster, J.P.; Macdonald, D.W. Fatal attraction in rats infected with Toxoplasma gondii. Proc. R. Soc. London. Ser. B Biol. Sci. 2000, 267, 1591–1594. [Google Scholar] [CrossRef] [PubMed]
- Hari Dass, S.A.; Vyas, A. Toxoplasma gondii infection reduces predator aversion in rats through epigenetic modulation in the host medial amygdala. Mol. Ecol. 2014, 23, 6114–6122. [Google Scholar] [CrossRef]
- Vyas, A.; Kim, S.-K.; Giacomini, N.; Boothroyd, J.C.; Sapolsky, R.M. Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc. Natl. Acad. Sci. USA 2007, 104, 6442–6447. [Google Scholar] [CrossRef]
- Parlog, A.; Schlüter, D.; Dunay, I.R. Toxoplasma gondii-induced neuronal alterations. Parasite Immunol. 2015, 37, 159–170. [Google Scholar] [CrossRef]
- Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hywel-Jones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. The life of a dead ant: The expression of an adaptive extended phenotype. Am. Nat. 2009, 174, 424–433. [Google Scholar] [CrossRef]
- Hunter, P. Extended phenotype redux. How far can the reach of genes extend in manipulating the environment of an organism? EMBO Rep. 2009, 10, 212. [Google Scholar] [CrossRef]
- Dawkins, R. The Extended Phenotype: The Long Reach of the Gene; Oxford University Press: Oxford, UK, 2008; ISBN 978-0198788911. [Google Scholar]
- Hughes, D.P.; Araújo, J.P.M.; Loreto, R.G.; Quevillon, L.; de Bekker, C.; Evans, H.C. From so simple a beginning: The evolution of behavioral manipulation by fungi. Adv. Genet. 2016, 94, 437–469. [Google Scholar] [CrossRef]
- Hohorst, W.; Graefe, G. Ameisen—Obligatorische Zwischenwirte des Lanzettegels (Dicrocoelium dendriticum). Naturwissenschaften 1961, 48, 229–230. [Google Scholar] [CrossRef]
- Moore, J. The behavior of parasitized animals. Bioscience 1995, 45, 89–96. [Google Scholar] [CrossRef]
- Maeyama, T.; Terayama, M.; Matsumoto, T. The abnormal behavior of Colobopsis sp. (Hymenoptera, Formicidae) parasitized by Mermis (Nematoda) in Papua New Guinea. Sociobiology 1994, 24, 115–119. [Google Scholar]
- Lovett, B.; St. Leger, R.J.; de Fine Licht, H.H. Going gentle into that pathogen-induced goodnight. J. Invertebr. Pathol. 2020, 174, 107398. [Google Scholar] [CrossRef] [PubMed]
- Will, I.; Attardo, G.M.; de Bekker, C. Multiomic interpretation of fungus-infected ant metabolomes during manipulated summit disease. Sci. Rep. 2023, 13, 14363. [Google Scholar] [CrossRef]
- de Bekker, C.; Ohm, R.A.; Loreto, R.G.; Sebastian, A.; Albert, I.; Merrow, M.; Brachmann, A.; Hughes, D.P. Gene expression during zombie ant biting behavior reflects the complexity underlying fungal parasitic behavioral manipulation. BMC Genom. 2015, 16, 620. [Google Scholar] [CrossRef]
- Trinh, T.; Ouellette, R.; de Bekker, C. Getting lost: The fungal hijacking of ant foraging behaviour in space and time. Anim. Behav. 2021, 181, 165–184. [Google Scholar] [CrossRef]
- Will, I.; Das, B.; Trinh, T.; Brachmann, A.; Ohm, R.A.; de Bekker, C. Genetic underpinnings of host manipulation by Ophiocordyceps as revealed by comparative transcriptomics. G3 Genes Genomes Genet. 2020, 10, 2275–2296. [Google Scholar] [CrossRef]
- Loreto, R.G.; Hughes, D.P. The metabolic alteration and apparent preservation of the zombie ant brain. J. Insect Physiol. 2019, 118, 103918. [Google Scholar] [CrossRef]
- Zheng, S.; Loreto, R.; Smith, P.; Patterson, A.; Hughes, D.; Wang, L. Specialist and generalist fungal parasites induce distinct biochemical changes in the mandible muscles of their host. Int. J. Mol. Sci. 2019, 20, 4589. [Google Scholar] [CrossRef]
- Kobmoo, N.; Wichadakul, D.; Arnamnart, N.; Rodríguez De La Vega, R.C.; Luangsa-ard, J.J.; Giraud, T. A genome scan of diversifying selection in Ophiocordyceps zombie-ant fungi suggests a role for enterotoxins in co-evolution and host specificity. Mol. Ecol. 2018, 27, 3582–3598. [Google Scholar] [CrossRef]
- Mangold, C.A.; Ishler, M.J.; Loreto, R.G.; Hazen, M.L.; Hughes, D.P. Zombie ant death grip due to hypercontracted mandibular muscles. J. Exp. Biol. 2019, 222, jeb200683. [Google Scholar] [CrossRef] [PubMed]
- Will, I.; Beckerson, W.C.; de Bekker, C. Using machine learning to predict protein-protein interactions between a zombie ant fungus and its carpenter ant host. Sci. Rep. 2023, 13, 13821. [Google Scholar] [CrossRef]
- Klein, S.L. Parasite manipulation of the proximate mechanisms that mediate social behavior in vertebrates. Physiol. Behav. 2003, 79, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Langager, M.M.; Adelman, J.S.; Hawley, D.M. Let’s stick together: Infection enhances preferences for social grouping in a songbird species. Ecol. Evol. 2023, 13, e10627. [Google Scholar] [CrossRef]
- Stephenson, J.F. Parasite-induced plasticity in host social behaviour depends on sex and susceptibility. Biol. Lett. 2019, 15, 20190557. [Google Scholar] [CrossRef]
- Rode, N.O.; Lievens, E.J.P.; Flaven, E.; Segard, A.; Jabbour-Zahab, R.; Sanchez, M.I.; Lenormand, T. Why join groups? Lessons from parasite-manipulated Artemia. Ecol. Lett. 2013, 16, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Ge, M.; Wu, M.; Duan, F.; Liang, J.; Chen, M.; Gracida, X.; Liu, H.; Yang, W.; Dar, A.R.; et al. Pathogenic bacteria modulate pheromone response to promote mating. Nature 2023, 613, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Nadler, L.E.; Adamo, S.A.; Hawley, D.M.; Binning, S.A. Mechanisms and consequences of infection-induced phenotypes. Funct. Ecol. 2023, 37, 796–800. [Google Scholar] [CrossRef]
- Porras, M.F.; Agudelo-Cantero, G.A.; Santiago-Martínez, M.G.; Navas, C.A.; Loeschcke, V.; Sørensen, J.G.; Rajotte, E.G. Fungal infections lead to shifts in thermal tolerance and voluntary exposure to extreme temperatures in both prey and predator insects. Sci. Rep. 2021, 11, 21710. [Google Scholar] [CrossRef]
- Andrade, L.C.; Majerowicz, D.; Oliveira, P.L.; Guarneri, A.A. Alterations in energy metabolism of Rhodnius prolixus induced by Trypanosoma rangeli infection. Insect Biochem. Mol. Biol. 2023, 159, 103987. [Google Scholar] [CrossRef]
- Evans, O.; Caragata, E.P.; McMeniman, C.J.; Woolfit, M.; Green, D.C.; Williams, C.R.; Franklin, C.E.; O’Neill, S.L.; McGraw, E.A. Increased locomotor activity and metabolism of Aedes aegypti infected with a lifeshortening strain of Wolbachia pipientis. J. Exp. Biol. 2009, 212, 1436–1441. [Google Scholar] [CrossRef]
- Tallon, A.K.; Lorenzo, M.G.; Moreira, L.A.; Martinez Villegas, L.E.; Hill, S.R.; Ignell, R. Dengue infection modulates locomotion and host seeking in Aedes aegypti. PLoS Negl. Trop. Dis. 2020, 14, e0008531. [Google Scholar] [CrossRef] [PubMed]
- McNeil, J.N.; Millar, J.G. Chemical communication: Pheromones and allelochemicals. In The Insects; Simpson, S.J., Douglas, A.E., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 857–900. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behavior; Cambridge University Press: Cambridge, UK, 2014; ISBN 9780521112901. [Google Scholar]
- Yew, J.Y.; Chung, H. Insect pheromones: An overview of function, form, and discovery. Prog. Lipid Res. 2015, 59, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.T.; Vaníčková, L. Tephritid fruit fly semiochemicals: Current knowledge and future perspectives. Insects 2021, 12, 408. [Google Scholar] [CrossRef] [PubMed]
- Engl, T.; Kaltenpoth, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386–397. [Google Scholar] [CrossRef]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- van den Bosch, T.J.M.; Welte, C.U. Detoxifying symbionts in agriculturally important pest insects. Microb. Biotechnol. 2017, 10, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Macke, E.; Tasiemski, A.; Massol, F.; Callens, M.; Decaestecker, E. Life history and eco-evolutionary dynamics in light of the gut microbiota. Oikos 2017, 126, 508–531. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Khan, M.M.; Bamisile, B.S.; Hafeez, M.; Qasim, M.; Rasheed, M.T.; Rasheed, M.A.; Ahmad, S.; Shahid, M.I.; Xu, Y. Role of insect gut microbiota in pesticide degradation: A review. Front. Microbiol. 2022, 13, 870462. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Oliver, K.M.; Martinez, A.J. How resident microbes modulate ecologically-important traits of insects. Curr. Opin. Insect Sci. 2014, 4, 1–7. [Google Scholar] [CrossRef]
- Attardo, G.M.; Scolari, F.; Malacrida, A. Bacterial symbionts of tsetse flies: Relationships and functional interactions between tsetse flies and their symbionts. In Results and Problems in Cell Differentiation; Springer Science and Business Media Deutschland GmbH: Berlin/Heidelberg, Germany, 2020; Volume 69, pp. 497–536. [Google Scholar]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C.J. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef]
- Panteleev, D.Y.; Goryacheva, I.I.; Andrianov, B.V.; Reznik, N.L.; Lazebny, O.E.; Kulikov, A.M. The endosymbiotic bacterium Wolbachia enhances the nonspecific resistance to insect pathogens and alters behavior of Drosophila melanogaster. Russ. J. Genet. 2007, 43, 1066–1069. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T.; Matsumoto, S.; Simon, J.C.; Fukatsu, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef]
- Flórez, L.V.; Biedermann, P.H.W.; Engl, T.; Kaltenpoth, M. Defensive symbioses of animals with prokaryotic and eukaryotic microorganisms. Nat. Prod. Rep. 2015, 32, 904–936. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Pheromones and signature mixtures: Defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J. Comp. Physiol. A. Neuroethol. Sens. Neural. Behav. Physiol. 2010, 196, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, C.P.; Osborne, G.O.; Mulcock, A.P. Production of an insect sex attractant by symbiotic bacteria. Nature 1971, 230, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.S. The ecology of yeasts in the bark beetle holobiont: A century of research revisited. Microb Ecol. 2015, 69, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Kather, R.; Martin, S.J. Evolution of cuticular hydrocarbons in the Hymenoptera: A meta-analysis. J. Chem. Ecol. 2015, 41, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Parasite manipulation of host behavior: An update and frequently asked questions. In Advances in the Study of Behavior; Academic Press: Cambridge, MA, USA, 2010; Volume 41, pp. 151–186. [Google Scholar]
- Keesey, I.W.; Koerte, S.; Khallaf, M.A.; Retzke, T.; Guillou, A.; Grosse-Wilde, E.; Buchon, N.; Knaden, M.; Hansson, B.S. Pathogenic bacteria enhance dispersal through alteration of Drosophila social communication. Nat. Commun. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Burand, J.P. The sexually transmitted insect virus, Hz-2V. Virol. Sin. 2009, 24, 428–435. [Google Scholar] [CrossRef]
- Kingan, T.G.; Bodnar, W.M.; Raina, A.K.; Shabanowitz, J.; Hunt, D.F. The loss of female sex pheromone after mating in the corn earworm moth Helicoverpa zea: Identification of a male pheromonostatic peptide. Proc. Natl. Acad. Sci. USA 1995, 92, 5082–5086. [Google Scholar] [CrossRef]
- Llopis-Giménez, A.; Caballero-Vidal, G.; Jacquin-Joly, E.; Crava, C.M.; Herrero, S. Baculovirus infection affects caterpillar chemoperception. Insect Biochem. Mol. Biol. 2021, 138, 103648. [Google Scholar] [CrossRef]
- Pantha, P.; Chalivendra, S.; Oh, D.H.; Elderd, B.D.; Dassanayake, M. A tale of two transcriptomic responses in agricultural pests via host defenses and viral replication. Int. J. Mol. Sci. 2021, 22, 3568. [Google Scholar] [CrossRef]
- Sabatier, L.; Jouanguy, E.; Dostert, C.; Zachary, D.; Dimarcq, J.-L.; Bulet, P.; Imler, J.-L. Pherokine-2 and -3. Two Drosophila molecules related to pheromone/odor-binding proteins induced by viral and bacterial infections. Eur. J. Biochem. 2003, 270, 3398–3407. [Google Scholar] [CrossRef]
- Sim, S.; Ramirez, J.L.; Dimopoulos, G. Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulate infection and blood-feeding behavior. PLoS Pathog. 2012, 8, e1002631. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.; Bohn, M.C.; Dornbusch, S.; Becker, S.C.; Stern, M. Influence of RVFV infection on olfactory perception and behavior in Drosophila melanogaster. Pathogens 2023, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Ban, L.; Ahmed, E.; Ninkovic, V.; Delp, G.; Glinwood, R. Infection with an insect virus affects olfactory behaviour and interactions with host plant and natural enemies in an aphid. Entomol. Exp. Appl. 2008, 127, 108–117. [Google Scholar] [CrossRef]
- Palottini, F.; Manrique, G. Compounds released by disturbed adults of the haematophagous bug Triatoma infestans (Hemiptera: Reduviidae): Behavioural effects of single compounds and binary mixtures. Physiol. Entomol. 2016, 41, 234–240. [Google Scholar] [CrossRef]
- Lobo, L.S.; Girotti, J.R.; Mijailovsky, S.J.; Fernandes, É.K.K.; Luz, C.; Pedrini, N. Synthesis and secretion of volatile short-chain fatty acids in Triatoma infestans infected with Beauveria bassiana. Med. Vet. Entomol. 2018, 32, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Lecuona, R.; Riba, G.; Cassier, P.; Clement, J.L. Alterations of insect epicuticular hydrocarbons during infection with Beauveria bassiana or B. brongniartii. J. Invertebr. Pathol. 1991, 58, 10–18. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Alaux, C.; Tchamitchan, S.; Brunet, J.-L.; Plettner, E.; Belzunces, L.P.; Le Conte, Y. Nosema spp. infection alters pheromone production in honey bees (Apis mellifera). J. Chem. Ecol. 2010, 36, 522–525. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Crauser, D.; Beslay, D.; Costagliola, G.; Soubeyrand, S.; Kretzchmar, A.; Le Conte, Y. Flight behavior and pheromone changes associated to Nosema ceranae infection of honey bee workers (Apis mellifera) in field conditions. J. Invertebr. Pathol. 2013, 113, 42–51. [Google Scholar] [CrossRef]
- Alaux, C.; Folschweiller, M.; McDonnell, C.; Beslay, D.; Cousin, M.; Dussaubat, C.; Brunet, J.L.; Conte, Y. Le Pathological effects of the microsporidium Nosema ceranae on honey bee queen physiology (Apis mellifera). J. Invertebr. Pathol. 2011, 106, 380–385. [Google Scholar] [CrossRef]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef]
- Mayack, C.; Broadrup, R.L.; Schick, S.J.; Eppley, E.J.; Khan, Z.; Macherone, A. Increased alarm pheromone component is associated with Nosema ceranae infected honeybee colonies. R. Soc. Open Sci. 2021, 8, rsos.210194. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.E.; Oma, E.A. Pest control by Nosema locustae a pathogen of grasshoppers and crickets. In Microbial Control of Pests and Plant Diseases, 1970–1980; Burgess, H.D., Ed.; Academic Press: New York, NY, USA, 1981; pp. 573–585. [Google Scholar]
- Shi, W.; Guo, Y.; Xu, C.; Tan, S.; Miao, J.; Feng, Y.; Zhao, H.; St. Leger, R.J.; Fang, W. Unveiling the mechanism by which microsporidian parasites prevent locust swarm behavior. Proc. Natl. Acad. Sci. USA 2014, 111, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Roy, H.E.; Baverstock, J.; Chamberlain, K.; Pell, J.K. Do aphids infected with entomopathogenic fungi continue to produce and respond to alarm pheromone? Biocontrol Sci. Technol. 2005, 15, 859–866. [Google Scholar] [CrossRef]
- Paszkiewicz, M.; Gołębiowski, M.; Sychowska, J.; Boguś, M.I.; Włóka, E.; Stepnowski, P. The effect of the entomopathogenic fungus Conidiobolus coronatus on the composition of cuticular and internal lipids of Blatta orientalis females. Physiol. Entomol. 2016, 41, 111–120. [Google Scholar] [CrossRef]
- Pull, C.D.; Ugelvig, L.V.; Wiesenhofer, F.; Grasse, A.V.; Tragust, S.; Schmitt, T.; Brown, M.J.F.; Cremer, S. Destructive disinfection of infected brood prevents systemic disease spread in ant colonies. Elife 2018, 7, e32073. [Google Scholar] [CrossRef]
- Naundrup, A.; Bohman, B.; Kwadha, C.A.; Jensen, A.B.; Becher, P.G.; De Fine Licht, H.H. Pathogenic fungus uses volatiles to entice male flies into fatal matings with infected female cadavers. ISME J. 2022, 16, 2388–2397. [Google Scholar] [CrossRef]
- Kelly, M.; Su, C.-Y.; Schaber, C.; Crowley, J.R.; Hsu, F.-F.; Carlson, J.R.; Odom, A.R. Malaria parasites produce volatile mosquito attractants. MBio 2015, 6, e00235-15. [Google Scholar] [CrossRef]
- Claudio-Piedras, F.; Recio-Tótoro, B.; Cime-Castillo, J.; Condé, R.; Maffei, M.; Lanz-Mendoza, H. Dietary and Plasmodium challenge effects on the cuticular hydrocarbon profile of Anopheles albimanus. Sci. Rep. 2021, 11, 11258. [Google Scholar] [CrossRef]
- Ebrahim, S.A.M.; Dweck, H.K.M.; Weiss, B.L.; Carlson, J.R. A volatile sex attractant of tsetse flies. Science 2023, 379, 660. [Google Scholar] [CrossRef]
- Hu, C.; Rio, R.V.M.; Medlock, J.; Haines, L.R.; Nayduch, D.; Savage, A.F.; Guz, N.; Attardo, G.M.; Pearson, T.W.; Galvani, A.P.; et al. Infections with immunogenic trypanosomes reduce tsetse reproductive fitness: Potential impact of different parasite strains on vector population structure. PLoS Negl. Trop. Dis. 2008, 2, e192. [Google Scholar] [CrossRef]
- Hurd, H.; Parry, G. Metacestode-induced depression of the production of, and response to, sex pheromone in the intermediate host Tenebrio molitor. J. Invertebr. Pathol. 1991, 58, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Stevens, L. Behavioral changes in Tribolium beetles infected with a tapeworm: Variation in effects between beetle species and among genetic strains. Am. Nat. 1994, 143, 830–847. [Google Scholar] [CrossRef]
- Yan, G.; Stevens, L. Selection by parasites on components of fitness in Tribolium beetles: The effect of intraspecific competition. Am. Nat. 1995, 146, 795–813. [Google Scholar] [CrossRef]
- Yan, G.; Phillips, T.W. Influence of tapeworm infection on the production of aggregation pheromone and defensive compounds in Tribolium castaneum. J. Parasitol. 1996, 82, 1037. [Google Scholar] [CrossRef] [PubMed]
- Pai, A.; Yan, G. Effects of tapeworm infection on male reproductive success and mating vigor in the red flour beetle, Tribolium castaneum. J. Parasitol. 2003, 89, 516–521. [Google Scholar] [CrossRef]
- Keymer, A.E. The influence of Hymenolepis diminuta on the survival and fecundity of the intermediate host, Tribolium confusum. Parasitology 1980, 81, 405–421. [Google Scholar] [CrossRef]
- Robb, T.; Reid, M.L. Parasite-induced changes in the behaviour of cestode-infected beetles: Adaptation or simple pathology? Can. J. Zool. 1996, 74, 1268–1274. [Google Scholar] [CrossRef]
- Novak, M.; Modha, A.; Blackburn, B.J. d-[1-13C]Glucose metabolism of Tribolium confusum parasitized by hymenolepid metacestodes. J. Invertebr. Pathol. 1993, 62, 302–307. [Google Scholar] [CrossRef]
- Zhang, X.; Machado, R.A.R.; Van Doan, C.; Arce, C.C.M.; Hu, L.; Robert, C.A.M. Entomopathogenic nematodes increase predation success by inducing cadaver volatiles that attract healthy herbivores. Elife 2019, 8, e46668. [Google Scholar] [CrossRef]
- Mazzoni, V.; Anfora, G. Behavioral manipulation for pest control. Insects 2021, 12, 287. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Poidevin, M.; Pradervand, S.; Lemaitre, B. Drosophila intestinal response to bacterial infection: Activation of host defense and stem cell proliferation. Cell Host Microbe 2009, 5, 200–211. [Google Scholar] [CrossRef]
- Chambers, M.C.; Song, K.H.; Schneider, D.S. Listeria monocytogenes infection causes metabolic shifts in Drosophila melanogaster. PLoS ONE 2012, 7, e50679. [Google Scholar] [CrossRef]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster—From microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Neyen, C.; Lemaitre, B. Sensing Gram-negative bacteria: A phylogenetic perspective. Curr. Opin. Immunol. 2016, 38, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Basset, A.; Khush, R.S.; Braun, A.; Gardan, L.; Boccard, F.; Hoffmann, J.A.; Lemaitre, B. The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response. Proc. Natl. Acad. Sci. USA 2000, 97, 3376–3381. [Google Scholar] [CrossRef] [PubMed]
- Vodovar, N.; Vinals, M.; Liehl, P.; Basset, A.; Degrouard, J.; Spellman, P.; Boccard, F.; Lemaitre, B. Drosophila host defense after oral infection by an entomopathogenic Pseudomonas species. Proc. Natl. Acad. Sci. USA 2005, 102, 11414–11419. [Google Scholar] [CrossRef] [PubMed]
- Flyg, C.; Kenne, K.; Boman, H.G. Insect pathogenic properties of Serratia marcescens: Phage-resistant mutants with a decreased resistance to Cecropia immunity and a decreased virulence to Drosophila. J. Gen. Microbiol. 1980, 120, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Liehl, P.; Blight, M.; Vodovar, N.; Boccard, F.; Lemaitre, B. Prevalence of local immune response against oral infection in a Drosophila/Pseudomonas infection model. PLoS Pathog. 2006, 2, e56. [Google Scholar] [CrossRef]
- Younes, S.; Al-Sulaiti, A.; Nasser, E.A.A.; Najjar, H.; Kamareddine, L. Drosophila as a model organism in host–pathogen interaction studies. Front. Cell. Infect. Microbiol. 2020, 10, 214. [Google Scholar] [CrossRef]
- Douglas, A.E. The Drosophila model for microbiome research. Lab Anim. 2018, 47, 157–164. [Google Scholar] [CrossRef]
- Herzog, G.A.; Phillips, J.R. Manifestation of an abnormal reproductive system in a laboratory strain of the bollworm Heliothis zea. J. Georg. Entomol. Soc. 1982, 17, 506. [Google Scholar]
- Hamm, J.J.; Carpenter, J.E.; Styer, E.L. Oviposition day effect on incidence of agonadal progeny of Helicoverpa zea (Lepidoptera: Noctuidae) infected with a virus. Ann. Entomol. Soc. Am. 1996, 89, 266–275. [Google Scholar] [CrossRef]
- Rallis, C.P.; Burand, J.P. Pathology and ultrastructure of Hz-2V infection in the agonadal female corn earworm, Helicoverpa zea. J. Invertebr. Pathol. 2002, 81, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Burand, J.P.; Rallis, C.P.; Tan, W. Horizontal transmission of Hz-2V by virus infected Helicoverpa zea moths. J. Invertebr. Pathol. 2004, 85, 128–131. [Google Scholar] [CrossRef]
- Raina, A.K.; Adams, J.R.; Lupiani, B.; Lynn, D.E.; Kim, W.; Burand, J.P.; Dougherty, E.M. Further characterization of the gonad-specific virus of corn earworm, Helicoverpa zea. J. Invertebr. Pathol. 2000, 76, 6–12. [Google Scholar] [CrossRef]
- Rallis, C.P.; Burand, J.P. Pathology and ultrastructure of the insect virus, Hz-2V, infecting agonadal male corn earworms, Helicoverpa zea. J. Invertebr. Pathol. 2002, 80, 81–89. [Google Scholar] [CrossRef]
- Kamita, S.G.; Nagasaka, K.; Chua, J.W.; Shimada, T.; Mita, K.; Kobayashi, M.; Maeda, S.; Hammock, B.D. A baculovirus-encoded protein tyrosine phosphatase gene induces enhanced locomotory activity in a lepidopteran host. Proc. Natl. Acad. Sci. USA 2005, 102, 2584–2589. [Google Scholar] [CrossRef]
- Miller, L.K. The Baculoviruses; Springer: Berlin/Heidelberg, Germany, 1997; ISBN 0306456419. [Google Scholar]
- Elderd, B.D. Developing models of disease transmission: Insights from ecological studies of insects and their baculoviruses. PLoS Pathog. 2013, 9, e1003372. [Google Scholar] [CrossRef]
- Labeur, C.; Dallerac, R.; Wicker-Thomas, C. Involvement of desat1 gene in the control of Drosophila melanogaster pheromone biosynthesis. Genetica 2002, 114, 269–274. [Google Scholar] [CrossRef]
- Bousquet, F.; Nojima, T.; Houot, B.; Chauvel, I.; Chaudy, S.; Dupas, S.; Yamamoto, D.; Ferveur, J.-F. Expression of a desaturase gene, desat1, in neural and nonneural tissues separately affects perception and emission of sex pheromones in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 249–254. [Google Scholar] [CrossRef]
- Hortsch, M.; Olson, A.; Fishman, S.; Soneral, S.N.; Marikar, Y.; Dong, R.; Jacobs, J.R. The expression of MDP-1, a component of Drosophila embryonic basement membranes, is modulated by apoptotic cell death. Int. J. Dev. Biol. 2003, 42, 33–42. [Google Scholar]
- Gravot, E.; Thomas-Orillard, M.; Jeune, B. Virulence variability of the Drosophila C virus and effects of the microparasite on demographic parameters of the host (Drosophila melanogaster). J. Invertebr. Pathol. 2000, 75, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Jousset, F.X.; Plus, N. Study of the vertical transmission and horizontal transmission of “Drosophila melanogaster” and “Drosophila immigrans” picornavirus (author’s transl). Ann. Microbiol. 1975, 126, 231–249. [Google Scholar]
- Thomas-Orillard, M. Interaction between a picornavirus and a wild population of Drosophila melanogaster. Oecologia 1988, 75, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Levy, F.; Bulet, P.; Ehret-Sabatier, L. Proteomic analysis of the systemic immune response of Drosophila. Mol. Cell. Proteom. 2004, 3, 156–166. [Google Scholar] [CrossRef]
- Moret, Y.; Schmid-Hempel, P. Immune defence in bumble-bee offspring. Nature 2001, 414, 506. [Google Scholar] [CrossRef] [PubMed]
- Traniello, J.F.A.; Rosengaus, R.B.; Savoie, K. The development of immunity in a social insect: Evidence for the group facilitation of disease resistance. Proc. Natl. Acad. Sci. USA 2002, 99, 6838–6842. [Google Scholar] [CrossRef]
- D’Arcy, C.J.; Burnett, P.A.; Hewings, A.D. Detection, biological effects, and transmission of a virus of the aphid Rhopalosiphum padi. Virology 1981, 114, 268–272. [Google Scholar] [CrossRef]
- Hack, W.H.; Ricciardi, A.I.; Oscherov, B.; Olivetti de Bravi, M.G. Composition of the secretion of Brindley’s gland in Triatominae. Med. B. Aires. 1980, 40, 178–180. [Google Scholar]
- Manrique, G.; Vitta, A.C.R.; Ferreira, R.A.; Zani, C.L.; Unelius, C.R.; Lazzari, C.R.; Diotaiuti, L.; Lorenzo, M.G. Chemical communication in Chagas disease Vectors. Source, identity, and potential function of volatiles released by the metasternal and Brindley’s glands of Triatoma infestans adults. J. Chem. Ecol. 2006, 32, 2035–2052. [Google Scholar] [CrossRef]
- Crespo, J.G.; Manrique, G. Mating behavior of the hematophagous bug Triatoma infestans: Role of Brindley’s and metasternal glands. J. Insect Physiol. 2007, 53, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Palottini, F.; González, A.; Manrique, G. Filling dynamics of the Brindley’s glands in the blood-sucking bug Triatoma infestans (Hemiptera: Reduviidae). J. Insect Physiol. 2014, 71, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, G.J.; Bagnères, A.G. Insect hydrocarbons biology, biochemistry, and chemical ecology. In Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; pp. 1–492. [Google Scholar] [CrossRef]
- Carlson, D.A.; Mayer, M.S.; Silhacek, D.L.; James, J.D.; Beroza, M.; Bierl, B.A. Sex attractant pheromone of the house fly: Isolation, identification and synthesis. Science 1971, 174, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Antony, C.; Jallon, J.-M. The chemical basis for sex recognition in Drosophila melanogaster. J. Insect Physiol. 1982, 28, 873–880. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Dillwith, J.W.; Adams, T.S. Biosynthesis and endocrine regulation of sex pheromone production in Diptera. In Pheromone Biochemistry; Elsevier: Amsterdam, The Netherlands, 1987; pp. 217–250. [Google Scholar]
- Sprenger, P.P.; Menzel, F. Cuticular hydrocarbons in ants (Hymenoptera: Formicidae) and other insects: How and why they differ among individuals, colonies, and species. Myrmecol. News 2020, 30, 1–26. [Google Scholar]
- Makki, R.; Cinnamon, E.; Gould, A.P. The development and functions of oenocytes. Annu. Rev. Entomol. 2014, 59, 405–425. [Google Scholar] [CrossRef]
- Ferveur, J.F.; Savarit, F.; O’Kane, C.J.; Sureau, G.; Greenspan, R.J.; Jallon, J.M. Genetic feminization of pheromones and its behavioral consequences in Drosophila males. Science 1997, 276, 1555–1558. [Google Scholar] [CrossRef]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized hepatocyte-like cells regulate Drosophila lipid metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef]
- Huang, K.; Liu, Y.; Perrimon, N. Roles of insect oenocytes in physiology and their relevance to human metabolic diseases. Front. Insect Sci. 2022, 2, 859847. [Google Scholar] [CrossRef]
- Juárez, M.P.; Fernández, G.C. Cuticular hydrocarbons of triatomines. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 711–730. [Google Scholar] [CrossRef]
- Dillwith, J.W.; Blomquist, G.J.; Nelson, D.R. Biosynthesis of the hydrocarbon components of the sex pheromone of the housefly, Musca domestica L. Insect Biochem. 1981, 11, 247–253. [Google Scholar] [CrossRef]
- Chung, H.; Carroll, S.B. Wax, sex and the origin of species: Dual roles of insect cuticular hydrocarbons in adaptation and mating. BioEssays 2015, 37, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, G.J.; Ginzel, M.D. Chemical ecology, biochemistry, and molecular biology of insect hydrocarbons. Annu. Rev. Entomol. 2021, 66, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Cook, H.W. Fatty acid desaturation and chain elongation in eukaryotes. New Compr. Biochem. 1996, 31, 129–152. [Google Scholar] [CrossRef]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Le Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef]
- Balabanidou, V.; Kampouraki, A.; Maclean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef]
- Kefi, M.; Balabanidou, V.; Douris, V.; Lycett, G.; Feyereisen, R.; Vontas, J. Two functionally distinct CYP4G genes of Anopheles gambiae contribute to cuticular hydrocarbon biosynthesis. Insect Biochem. Mol. Biol. 2019, 110, 52–59. [Google Scholar] [CrossRef]
- Calla, B.; MacLean, M.; Liao, L.H.; Dhanjal, I.; Tittiger, C.; Blomquist, G.J.; Berenbaum, M.R. Functional characterization of CYP4G11—A highly conserved enzyme in the western honey bee Apis mellifera. Insect Mol. Biol. 2018, 27, 661–674. [Google Scholar] [CrossRef]
- MacLean, M.; Nadeau, J.; Gurnea, T.; Tittiger, C.; Blomquist, G.J. Mountain pine beetle (Dendroctonus ponderosae) CYP4Gs convert long and short chain alcohols and aldehydes to hydrocarbons. Insect Biochem. Mol. Biol. 2018, 102, 11–20. [Google Scholar] [CrossRef]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2020, 126, 219–234. [Google Scholar] [CrossRef]
- Fan, Y.; Chase, J.; Sevala, V.L.; Schal, C. Lipophorin-facilitated hydrocarbon uptake by oocytes in the German cockroach Blattella germanica (L.). J. Exp. Biol. 2002, 205, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.P.; Bailey, A.M.; Cobb, B.; Vilcinskas, A. Fungi as elicitors of insect immune responses. Arch. Insect Biochem. Physiol. 2000, 44, 49–68. [Google Scholar] [CrossRef]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef]
- Lu, H.L.; St. Leger, R.J. Insect immunity to entomopathogenic fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar] [CrossRef]
- Lovett, B.; St. Leger, R.J. The insect pathogens. Microbiol. Spectr. 2017, 5, 925–943. [Google Scholar] [CrossRef]
- Napolitano, R.; Juárez, M.P. Entomopathogenous fungi degrade epicuticular hydrocarbons of Triatoma infestans. Arch. Biochem. Biophys. 1997, 344, 208–214. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Zhang, S.; Keyhani, N.O. Targeting of insect epicuticular lipids by the entomopathogenic fungus Beauveria bassiana: Hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microbiol. 2013, 4, 42518. [Google Scholar] [CrossRef]
- Hansen, A.N.; De Fine Licht, H.H. Why are there so few examples of entomopathogenic fungi that manipulate host sexual behaviors? Fungal Ecol. 2018, 38, 21–27. [Google Scholar] [CrossRef]
- Murray, Z.L.; Keyzers, R.A.; Barbieri, R.F.; Digby, A.P.; Lester, P.J. Two pathogens change cuticular hydrocarbon profiles but neither elicit a social behavioural change in infected honey bees, Apis mellifera (Apidae: Hymenoptera). Austral Entomol. 2016, 55, 147–153. [Google Scholar] [CrossRef]
- van Dooren, G.G.; Striepen, B. The algal past and parasite present of the apicoplast. Annu. Rev. Microbiol. 2013, 67, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Drijfhout, F.P.; Kather, R.; Martin, S.J. The role of cuticular hydrocarbons in insects. In Behavioral and Chemical Ecology; Zhang, W., Liu, H., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2009; pp. 91–114. ISBN 978-1-60741-099-7. [Google Scholar]
- Klewer, N.; Růžička, Z.; Schulz, S. (Z)-Pentacos-12-ene, an oviposition-deterring pheromone of Cheilomenes sexmaculata. J. Chem. Ecol. 2007, 33, 2167–2170. [Google Scholar] [CrossRef] [PubMed]
- Guédot, C.; Millar, J.G.; Horton, D.R.; Landolt, P.J. Identification of a sex attractant pheromone for male winterform pear psylla, Cacopsylla pyricola. J. Chem. Ecol. 2009, 35, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, A.A.; Pirk, C.W.W.; Crewe, R.M.; Njagi, P.G.N.; Gordon, I.; Torto, B. Nestmate recognition and the role of cuticular hydrocarbons in the African termite raiding ant Pachycondyla analis. J. Chem. Ecol. 2010, 36, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Foitzik, S.; Fröba, J.; Rüger, M.H.; Witte, V. Competition over workers: Fertility signalling in wingless queens of Hypoponera opacior. Insectes Soc. 2011, 58, 271–278. [Google Scholar] [CrossRef]
- Zinck, L.; Denis, D.; Hora, R.R.; Alaux, C.; Lenoir, A.; Hefetz, A.; Jaisson, P. Behavioral and chemical correlates of long-term queen adoption in the facultative polygynous ant Ectatomma tuberculatum. J. Insect Behav. 2009, 22, 362–374. [Google Scholar] [CrossRef]
- Adams, K.L.; Sawadogo, S.P.; Nignan, C.; Niang, A.; Paton, D.G.; Robert Shaw, W.; South, A.; Wang, J.; Itoe, M.A.; Werling, K.; et al. Cuticular hydrocarbons are associated with mating success and insecticide resistance in malaria vectors. Commun. Biol. 2021, 4, 911. [Google Scholar] [CrossRef]
- Weiss, I.; Hofferberth, J.; Ruther, J.; Stökl, J. Varying importance of cuticular hydrocarbons and iridoids in the species-specific mate recognition pheromones of three closely related Leptopilina species. Front. Ecol. Evol. 2015, 3, 131910. [Google Scholar] [CrossRef]
- Polerstock, A.R.; Eigenbrode, S.D.; Klowden, M.J. Mating alters the cuticular hydrocarbons of female Anopheles gambiae sensu stricto and Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 545–552. [Google Scholar] [CrossRef]
- Simarro, P.P.; Cecchi, G.; Franco, J.R.; Paone, M.; Diarra, A.; Ruiz-Postigo, J.A.; Fèvre, E.M.; Mattioli, R.C.; Jannin, J.G. Estimating and mapping the population at risk of sleeping sickness. PLoS Negl. Trop. Dis. 2012, 6, e1859. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.A.M. Africa’s Bane: The Tsetse Fly; Manly, T.M., Ed.; Collins: London, UK, 1969; ISBN 0002110369. [Google Scholar]
- Attardo, G.M.; Guz, N.; Strickler-Dinglasan, P.; Aksoy, S. Molecular aspects of viviparous reproductive biology of the tsetse fly (Glossina morsitans morsitans): Regulation of yolk and milk gland protein synthesis. J. Insect Physiol. 2006, 52, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Oropeza-Rodriguez, E.; Clifton, B.D.; Ranz, J.M. On the genetic basis of the effect of Spiroplasma on the male reproductive fitness of Glossina fuscipes fuscipes. PLoS Pathog. 2022, 18, e1010442. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Lahondère, C.; Attardo, G.M.; Michalkova, V.; Oyen, K.; Xiao, Y.; Aksoy, S. Warm blood meal increases digestion rate and milk protein production to maximize reproductive output for the tsetse fly, Glossina morsitans. Insects 2022, 13, 997. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.N. Male accessory secretions, their use and replenishment in Glossina (Diptera, Glossinidae). Bull. Entomol. Res. 1974, 64, 533–539. [Google Scholar] [CrossRef]
- Odhiambo, T.R.; Kokwaro, E.D.; Sequeira, L.M. Histochemical and ultrastructural studies of the male accessory reproductive glands and spermatophore of the tsetse, Glossina morsitans morsitans Westwood. Int. J. Trop. Insect Sci. 1983, 4, 227–236. [Google Scholar] [CrossRef]
- Hill, P.; Saunders, D.S.; Campbell, J.A. The production of “symbiont-free” Glossina morsitans and an associated loss of female fertility. Trans. R. Soc. Trop. Med. Hyg. 1973, 67, 727–728. [Google Scholar] [CrossRef]
- Gillott, C.; Langley, P.A. The control of receptivity and ovulation in the tsetse fly, Glossina morsitans. Physiol. Entomol. 1981, 6, 269–281. [Google Scholar] [CrossRef]
- Dame, D.A.; Ford, H.R. Multiple mating of Glossina morsitans Westw. and its potential effect on the sterile male technique. Bull. Entomol. Res. 1968, 58, 213–219. [Google Scholar] [CrossRef]
- Kokwaro, E.D.; Okot-Kotber, B.M.; Odhiambo, T.R.; Murithi, J.K. Biochemical and immunochemical evidence for the origin of the spermatophore material in Glossina morsitans morsitans Westwood. Experientia 1987, 43, 448–451. [Google Scholar] [CrossRef]
- Benoit, J.B.; Hansen, I.A.; Attardo, G.M.; Michalková, V.; Mireji, P.O.; Bargul, J.L.; Drake, L.L.; Masiga, D.K.; Aksoy, S. Aquaporins are critical for provision of water during lactation and intrauterine progeny hydration to maintain tsetse fly reproductive success. PLoS Negl. Trop. Dis. 2014, 8, e2517. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Benoit, J.B.; Michalkova, V.; Patrick, K.R.; Krause, T.B.; Aksoy, S. The homeodomain protein Ladybird Late regulates synthesis of milk proteins during pregnancy in the tsetse fly (Glossina morsitans). PLoS Negl. Trop. Dis. 2014, 8, e2645. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Benoit, J.B.; Michalkova, V.; Aksoy, E.; Takac, P.; Abd-Alla, A.M.M.; Malacrida, A.R.; Aksoy, S.; Attardo, G.M. The spermatophore in Glossina morsitans morsitans: Insights into male contributions to reproduction. Sci. Rep. 2016, 6, 20334. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Attardo, G.M.; Aksoy, E.; Weiss, B.; Savini, G.; Takac, P.; Abd-Alla, A.; Parker, A.G.; Aksoy, S.; Malacrida, A.R. Symbiotic microbes affect the expression of male reproductive genes in Glossina m. morsitans. BMC Microbiol. 2018, 18, 169. [Google Scholar] [CrossRef]
- Attardo, G.M.; Tam, N.; Parkinson, D.; Mack, L.K.; Zahnle, X.J.; Arguellez, J.; Takáč, P.; Malacrida, A.R. Interpreting morphological adaptations associated with viviparity in the tsetse fly Glossina morsitans (Westwood) by three-dimensional analysis. Insects 2020, 11, 651. [Google Scholar] [CrossRef]
- Lord, J.S.; Leyland, R.; Haines, L.R.; Barreaux, A.M.G.; Bonsall, M.B.; Torr, S.J.; English, S. Effects of maternal age and stress on offspring quality in a viviparous fly. Ecol. Lett. 2021, 24, 2113–2122. [Google Scholar] [CrossRef]
- Son, J.H.; Weiss, B.L.; Schneider, D.I.; Dera, K.M.; Gstöttenmayer, F.; Opiro, R.; Echodu, R.; Saarman, N.P.; Attardo, G.M.; Onyango, M.; et al. Infection with endosymbiotic Spiroplasma disrupts tsetse (Glossina fuscipes fuscipes) metabolic and reproductive homeostasis. PLoS Pathog. 2021, 17, e1009539. [Google Scholar] [CrossRef]
- Savini, G.; Scolari, F.; Ometto, L.; Rota-Stabelli, O.; Carraretto, D.; Gomulski, L.M.; Gasperi, G.; Abd-Alla, A.M.M.; Aksoy, S.; Attardo, G.M.; et al. Viviparity and habitat restrictions may influence the evolution of male reproductive genes in tsetse fly (Glossina) species. BMC Biol. 2021, 19, 211. [Google Scholar] [CrossRef]
- Benoit, J.B.; Attardo, G.M.; Baumann, A.A.; Michalkova, V.; Aksoy, S. Adenotrophic viviparity in tsetse flies: Potential for population control and as an insect model for lactation. Annu. Rev. Entomol. 2015, 60, 351–371. [Google Scholar] [CrossRef]
- Chahda, J.S.; Soni, N.; Sun, J.S.; Ebrahim, S.A.M.; Weiss, B.L.; Carlson, J.R. The molecular and cellular basis of olfactory response to tsetse fly attractants. PLoS Genet. 2019, 15, e1008005. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Hurd, H. Immune stimulation and malaria infection impose reproductive costs in Anopheles gambiae via follicular apoptosis. Microbes Infect. 2006, 8, 308–315. [Google Scholar] [CrossRef]
- Hacker, C.S. The differential effect of Plasmodium gallinaceum on the fecundity of several strains of Aedes aegypti. J. Invertebr. Pathol. 1971, 18, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Hurd, H.; Arme, C. Pathophysiology of Hymenolepis diminuta infections in Tenebrio molitor: Effect of parasitism on haemolymph proteins. Parasitology 1984, 89, 253–262. [Google Scholar] [CrossRef]
- Hurd, H.; Arme, C. Hymenolepis diminuta: Influence of metacestodes on synthesis and secretion of fat body protein and its ovarian sequestration in the intermediate host, Tenebrio molitor. Parasitology 1986, 93, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Hurd, H.; Arme, C. Hymenolepis diminuta: Effect of infection upon the patency of the follicular epithelium in the intermediate host, Tenebrio molitor. J. Invertebr. Pathol. 1987, 49, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Hitchen, S.J.; Shostak, A.W.; Belosevic, M. Hymenolepis diminuta (Cestoda) induces changes in expression of select genes of Tribolium confusum (Coleoptera). Parasitol. Res. 2009, 105, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Helms, A.M.; Ray, S.; Matulis, N.L.; Kuzemchak, M.C.; Grisales, W.; Tooker, J.F.; Ali, J.G. Chemical cues linked to risk: Cues from below-ground natural enemies enhance plant defences and influence herbivore behaviour and performance. Funct. Ecol. 2019, 33, 798–808. [Google Scholar] [CrossRef]
- Jehan, C.; Sabarly, C.; Rigaud, T.; Moret, Y. Age-specific fecundity under pathogenic threat in an insect: Terminal investment versus reproductive restraint. J. Anim. Ecol. 2022, 91, 101–111. [Google Scholar] [CrossRef]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction-immunity trade-offs in insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef]
- Páez, D.J.; Fleming-Davies, A.E. Understanding the Evolutionary Ecology of host–pathogen Interactions Provides Insights into the Outcomes of Insect Pest Biocontrol. Viruses 2020, 12, 141. [Google Scholar] [CrossRef]
- Slessor, K.N.; Kaminski, L.A.; King, G.G.S.; Borden, J.H.; Winston, M.L. Semiochemical basis of the retinue response to queen honey bees. Nature 1988, 332, 354–356. [Google Scholar] [CrossRef]
- Van Oystaeyen, A.; Oliveira, R.C.; Holman, L.; van Zweden, J.S.; Romero, C.; Oi, C.A.; D’Ettorre, P.; Khalesi, M.; Billen, J.; Wäckers, F.; et al. Conserved class of queen pheromones stops social insect workers from reproducing. Science 2014, 343, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Hefetz, A. Primer pheromones in social hymenoptera. Annu. Rev. Entomol. 2008, 53, 523–542. [Google Scholar] [CrossRef]
- Nunes, T.M.; Oldroyd, B.P.; Elias, L.G.; Mateus, S.; Turatti, I.C.; Lopes, N.P. Evolution of queen cuticular hydrocarbons and worker reproduction in stingless bees. Nat. Ecol. Evol. 2017, 1, 0185. [Google Scholar] [CrossRef]
- Vargo, E.L. Mutual pheromonal inhibition among queens in polygyne colonies of the fire ant Solenopsis invicta. Behav. Ecol. Sociobiol. 1992, 31, 205–210. [Google Scholar] [CrossRef]
- Vergoz, V.; Schreurs, H.A.; Mercer, A.R. Queen pheromone blocks aversive learning in young worker bees. Science 2007, 317, 384–386. [Google Scholar] [CrossRef]
- Princen, S.A.; Oliveira, R.C.; Ernst, U.R.; Millar, J.G.; Van Zweden, J.S.; Wenseleers, T. Honeybees possess a structurally diverse and functionally redundant set of queen pheromones. Proc. R. Soc. B 2019, 286, 20190517. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Fan, Y.; Hoover, S.E.R.; Winston, M.L. Genome-wide analysis reveals differences in brain gene expression patterns associated with caste and reproductive status in honey bees (Apis mellifera). Mol. Ecol. 2007, 16, 4837–4848. [Google Scholar] [CrossRef]
- Hoover, S.E.R.; Keeling, C.I.; Winston, M.L.; Slessor, K.N. The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften 2003, 90, 477–480. [Google Scholar] [CrossRef]
- Traynor, K.S.; Le Conte, Y.; Page, R.E. Queen and young larval pheromones impact nursing and reproductive physiology of honey bee (Apis mellifera) workers. Behav. Ecol. Sociobiol. 2014, 68, 2059–2073. [Google Scholar] [CrossRef]
- Pankiw, T.; Page, R.E. Effect of pheromones, hormones, and handling on sucrose response thresholds of honey bees (Apis mellifera L.). J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2003, 189, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Trawinski, A.M.; Fahrbach, S.E. Queen mandibular pheromone modulates hemolymph ecdysteroid titers in adult Apis mellifera workers. Apidologie 2018, 49, 346–358. [Google Scholar] [CrossRef]
- Velarde, R.A.; Robinson, G.E.; Fahrbach, S.E. Coordinated responses to developmental hormones in the Kenyon cells of the adult worker honey bee brain (Apis mellifera L.). J. Insect Physiol. 2009, 55, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Fahrbach, S.E.; Trawinski, A.M.; Velarde, R.A. Actions of developmental hormones in adult social insects. In Hormones, Brain and Behavior, 3rd ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 2, pp. 349–365. ISBN 9780128036082. [Google Scholar]
- Nunes, C.; Sucena, É.; Koyama, T. Endocrine regulation of immunity in insects. FEBS J. 2021, 288, 3928–3947. [Google Scholar] [CrossRef] [PubMed]
- Keith, S.A. Steroid hormone regulation of innate immunity in Drosophila melanogaster. PLoS Genet. 2023, 19, e1010782. [Google Scholar] [CrossRef]
- Reynolds, R.A.; Kwon, H.; Smith, R.C. 20-hydroxyecdysone primes innate immune responses that limit bacterial and malarial parasite survival in Anopheles gambiae. mSphere 2020, 5, e00983-19. [Google Scholar] [CrossRef]
- Cory, J.S.; Hoover, K. Plant-mediated effects in insect–pathogen interactions. Trends Ecol. Evol. 2006, 21, 278–286. [Google Scholar] [CrossRef]
- van Dijk, L.J.A.; Ehrlén, J.; Tack, A.J.M. The timing and asymmetry of plant–pathogen–insect interactions. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201303. [Google Scholar] [CrossRef]


{kind=link}
| Pathogens (Superkingdom/Kingdom) | Insect Host Species (Order) | Effects on Host | References |
|---|---|---|---|
| Bacteria | |||
| Pseudomonas entomophila Serratia marcescens Pectobacterium carotovorum | Drosophila spp. Aedes aegypti (Diptera) | Increased pheromone emission | [70] |
| Viruses | |||
| Hz-2V | Helicoverpa zea (Lepidoptera) | Females: calling behavior, increased pheromone production, stronger male attraction. Males: cannot produce pheromonostatic peptide | [8,71,72] |
| SeMNPV 1 | Spodoptera exigua (Lepidoptera) | Changes in expression of larval odorant receptors resulting in behavioral responses to its ligands | [73] |
| AcMNPV 2 | Spodoptera frugiperda Trichoplusia ni (Lepidoptera) | Co-induction of Desaturase1 and bond (pheromone production/perception and conspecific signaling). Enrichment in functions related to fatty acid biosynthesis and pheromone metabolisms | [74] |
| DCV 3 | Drosophila spp. (Diptera) | Induction of pherokine-2 | [75] |
| DENV-2 | Aedes aegypti (Diptera) | Effects on the expression of chemosensory-related genes involved in the regulation of blood feeding | [76] |
| RVFV 4 | D. melanogaster (Diptera) | Changes in odor response and activity | [77] |
| RhPV 5 | Rhopalosiphum padi (Hemiptera) | Increased sensitiveness to the alarm pheromone | [78] |
| Fungi | |||
| Beauveria bassiana | Triatoma infestans (Hemiptera) | Differences in the profile of volatile organic compounds. Increased expression of genes involved in the synthesis of volatile short-chain fatty acids | [79,80] |
| Beauveria bassiana Beauveria brongniartii | Melolontha melolontha (Coleoptera) Ostrinia nubilalis (Lepidoptera) | Alterations in cuticular hydrocarbon profile | [81] |
| Nosema spp. | Apis mellifera (Hymenoptera) | Workers: Increased production of the primer foraging pheromone ethyl oleate. Increased flight activity and mortality. Queen: Increased vitellogenin titer, antioxidant capacity, and mandibular pheromones. Increased synthesis of alarm pheromone component. Alterations in cuticular hydrocarbon profile | [82,83,84,85,86] |
| Paranosema (Nosema) locustae | Locusta migratoria manilensis (Orthoptera) | Inhibition of aggregation behavior due to decreased production of aggregation pheromone | [87,88] |
| Pandora neoaphidis | Acyrthosiphon pisum (Hemiptera) | Increased release of alarm pheromone | [89] |
| Conidiobolus coronatus | Blatta orientalis (Blattodea) | Increased production of cuticular hydrocarbons and other surface compounds | [90] |
| Metarhizium brunneum | Lasius neglectus (Hymenoptera) | Changes in cuticular hydrocarbon profile and alteration in volatile chemical cues emission | [91] |
| Entomophthora muscae | Musca domestica (Diptera) | Production of a mixture of volatile sesquiterpenes altering the CH profile in female cadavers | [92] |
| Eukaryota | |||
| Phylum Apicomplexa | |||
| Plasmodium falciparum | Anopheles gambiae (Diptera) | Production of terpenes putatively mediating mammalian host preference | [93] |
| Plasmodium berghei | Anopheles albimanus (Diptera) | Altered cuticular hydrocarbon profile | [94] |
| Phylum Euglenozoa | |||
| Trypanosoma brucei | Glossina morsitans morsitans (Diptera) | Altered chemical profiles of infected mated individuals. Reduced fecundity | [95,96] |
| Phylum Platyhelminthes | |||
| Hymenolepis diminuta | Tenebrio molitor (Coleoptera) | Decreased production of non-volatile copulatory pheromone in females. Negative effects on male response to pheromone | [97] |
| Tribolium castaneum (Coleoptera) | Altered behavior. Decreased male sperm precedence and fitness. Decreased production of defensive compounds | [98,99,100] | |
| Tribolium confusum (Coleoptera) | Reduced survival, fecundity, mating vigour, carbohydrate metabolism, and pheromone response. Upregulation of pheromone binding proteins | [101,102,103,104] | |
| Phylum Nematoda | |||
| Heterorhabditis bacteriophora | Diabrotica virgifera (Coleoptera) | Increased emission of volatiles such as butylated hydroxytoluene | [105] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyano, A.; Croce, A.C.; Scolari, F. Pathogen-Mediated Alterations of Insect Chemical Communication: From Pheromones to Behavior. Pathogens 2023, 12, 1350. https://doi.org/10.3390/pathogens12111350
Moyano A, Croce AC, Scolari F. Pathogen-Mediated Alterations of Insect Chemical Communication: From Pheromones to Behavior. Pathogens. 2023; 12(11):1350. https://doi.org/10.3390/pathogens12111350
Chicago/Turabian StyleMoyano, Andrea, Anna Cleta Croce, and Francesca Scolari. 2023. "Pathogen-Mediated Alterations of Insect Chemical Communication: From Pheromones to Behavior" Pathogens 12, no. 11: 1350. https://doi.org/10.3390/pathogens12111350
APA StyleMoyano, A., Croce, A. C., & Scolari, F. (2023). Pathogen-Mediated Alterations of Insect Chemical Communication: From Pheromones to Behavior. Pathogens, 12(11), 1350. https://doi.org/10.3390/pathogens12111350