
Thousand Cankers Disease in Walnut Trees in Europe: Current Status and Management

 , , , and
, , , and
Abstract
:1. Introduction
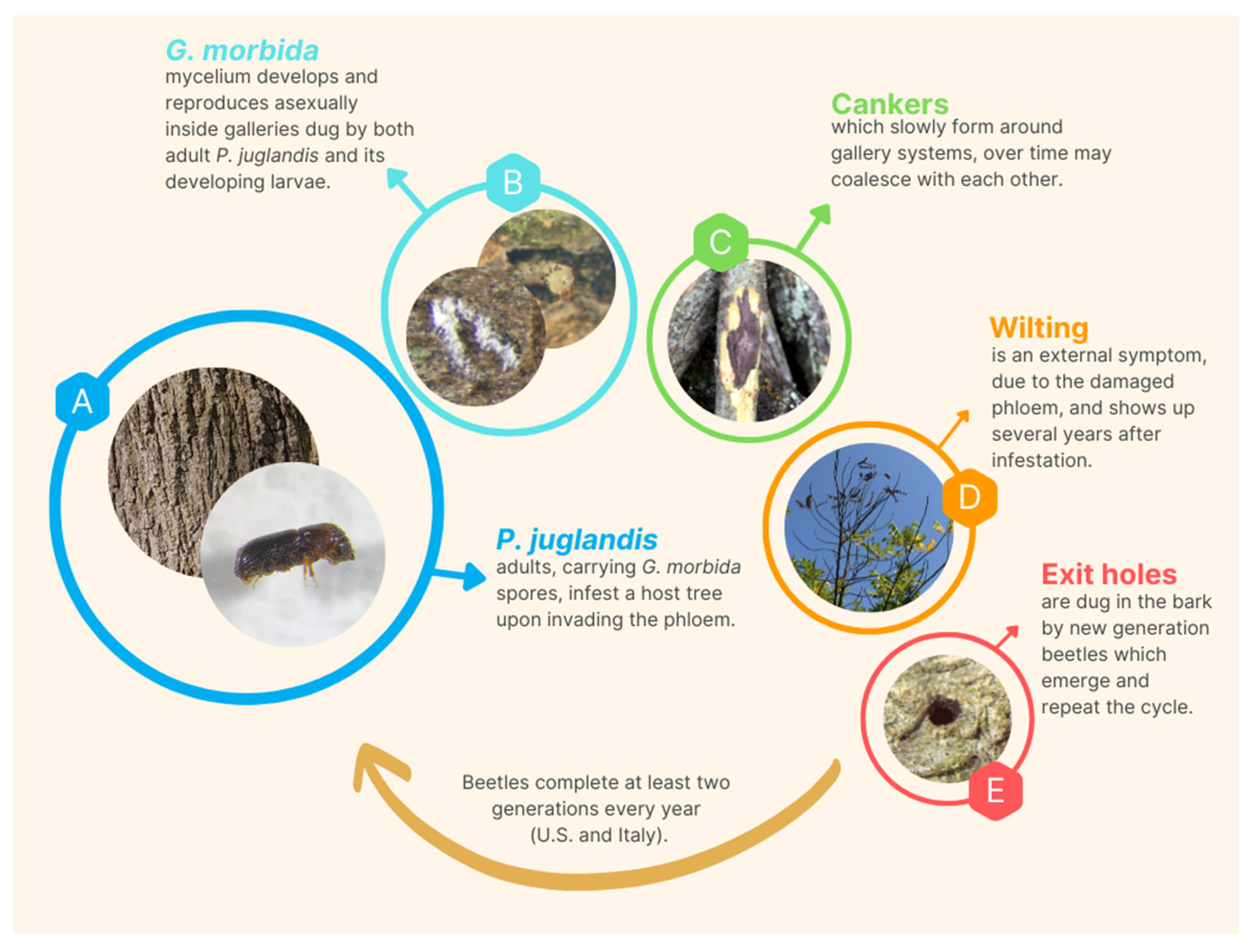
2. The Pathogen Geosmithia morbida
3. The Vector Pityophthorus juglandis
4. Interactions between Geosmithia morbida and Pityophthours juglandis
5. Potential Alternative Vectors of Geosmithia morbida
6. Symptomatology
7. Susceptible Hosts
8. TCD Integrated Management
8.1. Diagnosis
8.2. Prevention (Surveillance and Monitoring)
8.3. Eradication and Containment
8.4. Public Outreach
9. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nicolescu, V.N.; Rédei, K.; Vor, T.; Bastien, J.C.; Brus, R.; Benčať, T.; Đodan, M.; Cvjetkovic, B.; Andrašev, S.; La Porta, N.; et al. A review of black walnut (Juglans nigra L.) ecology and management in Europe. Trees 2020, 34, 1087–1112. [Google Scholar] [CrossRef]
- Rehnus, M.; Sorg, J.-P.; Pasinelli, G. Habitat and Cavity Tree Selection by White-Winged Woodpeckers Dendrocopos leucopterus in the Walnut-Fruit Forests of Kyrgyzstan. Acta Ornithol. 2011, 46, 80–92. [Google Scholar] [CrossRef]
- Newton, L.; Fowler, G. Pathway Assessment: Geosmithia sp. and Pityophthorus juglandis Blackman Movement from the Western into the Eastern United States; US Department of Agriculture Animal and Plant Health Inspection Service: Raleigh, NC, USA, 2009; p. 50. [Google Scholar]
- Kremer, D.; Čavlović, J.; Božić, M.; Dubravac, T. Distribution and Management of Black Walnut (Juglans nigra L.) in Croatia. Period. Biol. 2008, 110, 317–321. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Tisserat, N.; Cranshaw, W.; Leatherman, D.; Utley, C.; Alexander, K. Black Walnut Mortality in Colorado Caused by the Walnut Twig Beetle and Thousand Cankers Disease. Plant Health Prog. 2009, 10, 10. [Google Scholar] [CrossRef]
- Cranshaw, W.; Tisserat, N. Walnut Twig Beetle and Thousand Cankers Disease of Black Walnut. Ph.D. Thesis, Colorado State University Extension, Fort Collins, CO, USA, 2008. Available online: www.ext.colostate.edu/pubs/insect/0812_alert.pdf (accessed on 23 September 2022).
- Kolařík, M.; Freeland, E.; Utley, C.; Tisserat, N. Geosmithia morbida sp. Nov., a new phytopathogenic species living in symbiosis with the walnut twig beetle (Pityophthorus juglandis) on Juglans in USA. Mycologia 2011, 103, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.F.; Windham, M.T.; Haun, W.G.; Wiggins, G.J.; Lambdin, P.L. Initial assessment of thousand cankers disease on black walnut, Juglans nigra, in Eastern Tennessee. Forests 2011, 2, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Montecchio, L.; Faccoli, M. First record of thousand cankers disease Geosmithia morbida and walnut twig beetle Pityophthorus juglandis on Juglans nigra in Europe. Plant Disease 2014, 98, 696. [Google Scholar] [CrossRef] [PubMed]
- European and Mediterranean Plant Protection Organisation (EPPO). Pest Risk Analysis for Thousand Cankers Disease (Geosmithia morbida and Pityophthorus juglandis). 2015. Available online: http://www.eppo.int/QUARANTINE/Pest_Risk_Analysis/PRA_intro.htm (accessed on 1 October 2022).
- Montecchio, L.; Vettorazzo, M.; Faccoli, M. Thousand cankers disease in Europe: An overview. EPPO Bull. 2016, 46, 335–340. [Google Scholar] [CrossRef]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ginetti, B.; Pelleri, F.; Panzavolta, T. Thousand Cankers Disease caused by Geosmithia morbida and its insect vector Pityophthorus juglandis first reported on Juglans nigra in Tuscany, Central Italy. Plant Dis. 2019, 103, 369. [Google Scholar] [CrossRef]
- Kolařík, M.; Hulcr, J.; Tisserat, N.; De Beer, W.; Kostovčík, M.; Kolaříková, Z.; Seybold, S.J.; Rizzo, D.M. Geosmithia associated with bark beetles and woodborers in the western USA: Taxonomic diversity and vector specificity. Mycologia 2017, 109, 185–199. [Google Scholar] [CrossRef] [Green Version]
- Moricca, S.; Bracalini, M.; Benigno, A.; Panzavolta, T. Observations on the non-native Thousand Cankers Disease of Walnut in Europe’s southernmost outbreak. Glob. Ecol. Conserv. 2020, 23, e01159. [Google Scholar] [CrossRef]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ghelardini, L.; Furtado, E.L.; Marino, C.L.; Panzavolta, T. Thousand cankers disease in Juglans: Optimizing sampling and identification procedures for the vector Pityophthorus juglandis, and the causal agent Geosmithia Morb. MethodsX 2020, 7, 101174. [Google Scholar] [CrossRef]
- Veselská, T.; Skelton, J.; Kostovčík, M.; Hulcr, J.; Baldrian, P.; Chudíčková, M.; Cajthaml, T.; Vojtová, T.; Garcia-Fraile, P.; Kolařík, M. Adaptive traits of bark and ambrosia beetle-associated fungi. Fungal Ecol. 2019, 41, 165–176. [Google Scholar] [CrossRef]
- Tisserat, N.; Cranshaw, W.; Putnam, M.L.; Pscheidt, J.; Leslie, C.A.; Murray, M.; Hoffman, J.; Barkley, Y.; Alexander, K.; Seybold, S.J. Thousand Cankers Disease is Widespread in Black Walnut in the Western United States. Plant Health Prog. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Daniels, D.; Nix, K.; Wadl, P.; Vito, L.; Wiggins, G.; Windham, M.; Ownley, B.H.; Lambdin, P.L.; Grant, J.F.; Merten, P.; et al. Thousand cankers disease complex: A forest health issue that threatens Juglans species across the U.S. Forests 2016, 7, 260. [Google Scholar] [CrossRef] [Green Version]
- Moricca, S.; Ragazzi, A. Fungal endophytes in Mediterranean oak forests: A lesson from Discula quercina. Phytopathology 2008, 98, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazis, R.; Poplawski, L.; Klingeman, W.; Boggess, S.L.; Trigiano, R.N.; Graves, A.D.; Seybold, S.J.; Hadziabdic, D. Mycobiota associated with insect galleries in walnut with thousand cankers disease reveals a potential natural enemy against Geosmithia morbida. Fungal Biol. 2018, 122, 241–253. [Google Scholar] [CrossRef]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982. [Google Scholar]
- Seybold, S.J.; Dallara, P.L.; Hishinuma, S.M.; Flint, M.L. Detecting and Identifying the Walnut Twig Beetle: Monitoring Guidelines for the Invasive Vector of Thousand Cankers Disease of Walnut; UC IPM Program, University of California Agriculture and Natural Resources: Oakland, CA, USA, 2013; Available online: http://ipm.ucanr.edu/PDF/PESTNOTES/WTB_trapping.pdf (accessed on 22 September 2022).
- LaBonte, J.R.; Rabaglia, R.J. A Screening Aid for the Identification of the Walnut Twig Beetle, Pityophthorus juglandis Blackman; Plant Protection and Conservation Programs, Oregon Dept. of Agriculture: Salem, OR, USA, 2015; Available online: https://conference.ifas.ufl.edu/npdn/Docs/Workshop%20Materials/Walnut%20Twig%20Beetle%202015.pdf (accessed on 12 September 2022).
- Faccoli, M.; Simonato, M.; Rassati, D. Life history and geographical distribution of the walnut twig beetle, Pityophthorus juglandis (Coleoptera: Scolytinae), in southern Europe. J. Appl. Entomol. 2016, 140, 697–705. [Google Scholar] [CrossRef]
- Kees, A.M.; Hefty, A.R.; Venette, R.C.; Seybold, S.J.; Aukema, B.H. Flight capacity of the walnut twig beetle (Coleoptera: Scolytidae) on a laboratory flight mill. Environ. Entomol. 2017, 46, 633–641. [Google Scholar] [CrossRef]
- Marchioro, M.; Faccoli, M. Dispersal and colonization risk of the Walnut Twig Beetle, Pityophthorus juglandis, in southern Europe. J. Pest Sci. 2022, 95, 303–313. [Google Scholar] [CrossRef]
- Hishinuma, S.M. Interactions among the Walnut Twig Beetle, Pityophthorus juglandis, the Pathogenic Fungus, Geosmithia morbida, and Host Species in Thousand Cankers Disease in California. Ph.D. Thesis, University of California, Davis, CA, USA, 2017. [Google Scholar]
- Wilstermann, A.; Hoppe, B.; Schrader, G.; Delbianco, A.; Vos, S. Pest survey card on Geosmithia morbida and its vector Pityophthorus juglandis. EFSA Support. Publ. 2020, 17, 1894E. [Google Scholar] [CrossRef]
- Audley, J. Assessment of Pityophthorus juglandis Colonization Characteristics and Implications for Further Spread of Thousand Cankers Disease. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2015; p. 88.
- Luna, E.K.; Sitz, R.A.; Cranshaw, W.S.; Tisserat, N.A. The effect of temperature on survival of Pityophthorus juglandis (Coleoptera: Curculionidae). Environ. Entomol. 2013, 42, 1085–1091. [Google Scholar] [CrossRef]
- Luna, E.K.; Cranshaw, W.S.; Tisserat, N.A. Attraction of walnut twig beetle Pityophthorus juglandis (Coleoptera: Curculionidae) to the fungus Geosmithia morbida. Plant Health Progr. 2014, 15, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Blood, B.L.; Klingeman, W.E.; Paschen, M.A.; Hadžiabdic, Ð.; Couture, J.J.; Ginzel, M.D. Behavioral responses of Pityophthorus juglandis (Coleoptera: Curculionidae: Scolytinae) to volatiles of black walnut and Geosmithia morbida (Ascomycota: Hypocreales: Bionectriaceae), the causal agent of thousand cankers disease. Environ. Entomol. 2018, 47, 412–421. [Google Scholar] [CrossRef]
- Moore, M.; Juzwik, J.; Miller, F.; Roberts, L.; Ginzel, M.D. Detection of Geosmithia morbida on numerous insect species in four eastern States. Plant Health Prog. 2019, 20, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Chahal, K.; Gazis, R.; Klingeman, W.; Hadziabdic, D.; Lambdin, P.; Grant, J.; Windham, M. Assessment of alternative candidate subcortical insect vectors from walnut crowns in habitats quarantined for thousand cankers disease. Environ. Entomol. 2019, 48, 882–893. [Google Scholar] [CrossRef] [PubMed]
- EPPO. Thousand Cankers Disease Found in Lombardia, Emilia-Romagna and Toscana (Italy) EPPO Reporting Service (2019) 2019/102. Available online: https://gd.eppo.int/reporting/article-6532 (accessed on 2 September 2022).
- Graves, A.D.; Coleman, T.W.; Flint, M.L.; Seybold, S.J. Walnut twig beetle and thousand cankers disease: Field identification guide. UC IPM Progr. Univ. Calif. Agric. Nat. Resour. 2009. Available online: http://www.ipm.ucdavis.edu/thousandcankers (accessed on 2 September 2022).
- Cranshaw, W.; Tisserat, N. Questions and Answers about Thousand Cankers Disease of Walnut; Department of Bioagricultural Sciences and Pest Management, Colorado State University: Fort Collins, CO, USA, 2012; Available online: http://bspm.agsci.-colostate.edu/files/2013/03/Questions-and-AnswersRevision-April-2012.pdf (accessed on 11 September 2022).
- Seybold, S.J.; Klingeman, W.E.; Hishinuma, S.M.; Coleman, T.W.; Graves, A.D. Status and impact of walnut twig beetle in urban forest, orchard, and native forest ecosystems. J. For. 2019, 117, 152–163. [Google Scholar]
- Hishinuma, S.M.; Dallara, P.L.; Yaghmour, M.A.; Zerillo, M.M.; Parker, C.M.; Roubtsova, T.V.; Nguyen, T.L.; Tisserat, N.A.; Bostock, R.M.; Flint, M.L.; et al. Wingnut (Juglandaceae) as a new generic host for Pityophthorus juglandis (Coleoptera: Curculionidae) and the thousand cankers disease pathogen, Geosmithia morbida (Ascomycota: Hypocreales). Can. Entomol. 2016, 148, 83–91. [Google Scholar] [CrossRef]
- Zerillo, M.M.; Caballero, J.I.; Woeste, K.; Graves, A.D.; Hartel, C.; Pscheidt, J.W.; Tonos, J.; Broders, K.; Cranshaw, W.; Seybold, S.J.; et al. Population structure of Geosmithia morbida, the causal agent of thousand cankers disease of walnut trees in the United States. PLoS ONE 2014, 9, e112847. [Google Scholar] [CrossRef] [Green Version]
- Utley, C.; Nguyen, T.; Roubtsova, T.; Coggeshall, M.; Ford, T.M.; Grauke, L.J.; Graves, A.D.; Leslie, C.A.; McKenna, J.; Woeste, K.; et al. Susceptibility of walnut and hickory species to Geosmithia morbida. Plant Dis. 2013, 97, 601–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecchio, L.; Fanchin, G.; Simonato, M.; Faccoli, M. First record of Thousand Cankers Disease fungal pathogen Geosmithia morbida and Walnut Twig Beetle Pityophthorus juglandis on Juglans regia in Europe. Plant Dis. 2014, 98, 1445. [Google Scholar] [CrossRef] [PubMed]
- Seybold, S.J.; Coleman, T.W.; Dallara, P.L.; Dart, N.L.; Graves, A.D.; Pederson, L.A.; Spichiger, S.E. Recent collecting reveals new state records and geographic extremes in the distribution of the walnut twig beetle Pityophthorus juglandis Blackman (Coleoptera: Scolytidae), in the United States. Pan Pac. Entomol. 2012, 88, 277–280. [Google Scholar] [CrossRef]
- Hadziabdic, D.; Vito, L.M.; Windham, M.T.; Pscheidt, J.W.; Trigiano, R.N.; Kolařík, M. Genetic differentiation and spatial structure of Geosmithia morbida, the causal agent of thousand cankers disease in black walnut (Juglans nigra). Curr. Genet. 2014, 60, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Knížek, M.; Beaver, R. Taxonomy and systematics of bark and ambrosia beetles. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.C., Evans, H.F., Eds.; Kluwer Academic Publishers: Dordrecht, the Netherlands, 2004; pp. 41–54. [Google Scholar]
- Hamelin, R.C.; Roe, A.D. Genomic biosurveillance of forest invasive alien enemies: A story written in code. Evol. Appl. 2020, 13, 95–115. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, J.; Potvin, A.; Pelletier, G.; Stewart, D.; Feau, N.; Alayon, D.I.O.; Dale, A.L.; Coehlo, A.; Uzunovic, A.; Bilodeau, G.J.; et al. Molecular detection of 10 of the most unwanted alien forest pathogens in Canada using real-time PCR. PLoS ONE 2015, 10, e0134265. [Google Scholar] [CrossRef]
- Oren, E.; Klingeman, W.; Gazis, R.; Moulton, J.; Lambdin, P.; Coggeshall, M.; Hulcr, J.; Seybold, S.J.; Hadziabdic, D. A novel molecular toolkit for rapid detection of the pathogen and primary vector of thousand cankers disease. PLoS ONE 2018, 13, e0185087. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Bruscoli, T.; Bracalini, M.; Benigno, A.; Salemi, C.; Del Nista, D.; Aronadio, A.; et al. A duplex real-time PCR with probe for simultaneous detection of Geosmithia morbida and its vector Pityophthorus juglandis. PLoS ONE 2020, 15, e0241109. [Google Scholar] [CrossRef]
- Rizzo, D.; Moricca, S.; Bracalini, M.; Benigno, A.; Bernardo, U.; Luchi, N.; Da Lio, D.; Nugnes, F.; Cappellini, G.; Salemi, C.; et al. Rapid detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the loop-mediated isothermal amplification (LAMP) method. Plants 2021, 10, 1048. [Google Scholar] [CrossRef]
- Rizzo, D.; Aglietti, C.; Benigno, A.; Bracalini, M.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Aronadio, A.; Francia, C.; Luchi, N.; et al. Loop-Mediated Isothermal Amplification (LAMP) and SYBR Green qPCR for Fast and Reliable Detection of Geosmithia morbida (Kolařik) in Infected Walnut. Plants 2022, 11, 1239. [Google Scholar] [CrossRef]
- Hadziabdic, D.; Wadl, P.A.; Vito, L.M.; Boggess, S.L.; Scheffler, B.E.; Windham, M.T.; Trigiano, R.N. Development and characterization of sixteen microsatellite loci for Geosmithia morbida, the causal agent of thousand canker disease in black walnut (Juglans nigra). Conserv.Genet.Resour. 2012, 4, 287–289. [Google Scholar] [CrossRef]
- Stackhouse, T.; Boggess, S.L.; Hadziabdic, D.; Trigiano, R.N.; Ginzel, M.D.; Klingeman, W.E. Conventional gel electrophoresis and TaqMan probes enable rapid confirmation of Thousand Cankers Disease from diagnostic samples. Plant Dis. 2021, 105, 3171–3180. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.Y.; Botella, J.R. Advanced DNA-based point-of-care diagnostic methods for plant diseases detection. Front. Plant Sci. 2017, 8, 2016. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, G.; Prasannath, K. Recent advances in molecular diagnostics of fungal plant pathogens: A mini review. Front. Cell. Infect. Microbiol. 2021, 10, 600234. [Google Scholar] [CrossRef]
- Panzavolta, T.; Bracalini, M.; Benigno, A.; Moricca, S. Alien invasive pathogens and pests harming trees, forests, and plantations: Pathways, global consequences and management. Forests 2021, 12, 1364. [Google Scholar] [CrossRef]
- EU. Consolidated Text: Commission Implementing Regulation (EU) 2019/2072 of 28 November 2019 Establishing Uniform Conditions for the Implementation of Regulation (EU) 2016/2031 of the European Parliament and the Council, as Regards Protective Measures against Pests of Plants, and Repealing Commission Regulation (EC) No 690/2008 and Amending Commission Implementing Regulation (EU) 2018/2019. 2022. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02019R2072-20220411 (accessed on 2 September 2022).
- EU. Consolidated Text: Commission Implementing Regulation (EU) 2018/2019 of 18 December 2018 Establishing a Provisional List of High Risk Plants, Plant Products or Other Objects, within the Meaning of Article 42 of Regulation (EU) 2016/2031 and a List of Plants for Which Phytosanitary Certificates Are Not Required for Introduction into the Union, within the Meaning of Article 73 of that Regulation. 2022. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02018R2019-20220331 (accessed on 2 September 2022).
- Rugman-Jones, P.F.; Seybold, S.J.; Graves, A.D.; Stouthamer, R. Phylogeography of the walnut twig beetle, Pityophthorus juglandis, the vector of thousand cankers disease in North American walnut trees. PLoS ONE 2015, 10, e0118264. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.E., III; Fraedrich, S.W.; Taylor, A.; Merten, P.; Myers, S.W. Efficacy of heat treatment for the thousand cankers disease vector and pathogen in small black walnut logs. J. Econ. Entomol. 2014, 107, 174–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juzwik, J.; Yang, A.; Heller, S.; Moore, M.; Chen, Z.; White, M.; Wantuch, H.; Ginzel, M.; Mack, R. Vacuum Steam Treatment Effectiveness for Eradication of the Thousand Cankers Disease Vector and Pathogen in Logs From Diseased Walnut Trees. J. Econ. Entomol. 2021, 114, 100–111. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture (USDA). Thousand Cankers Disease Survey Guidelines for 2014; Forest Service (FS) and Plant Protection and Quarantine (PPQ): Riverdale, MD, USA, 2014. [Google Scholar]
- Seybold, S.J.; Dallara, P.L.; Nelson, L.J.; Graves, A.D.; Hishinuma, S.M.; Gries, R. Methods of Monitoring and Controlling the Walnut Twig Beetle, Pityophthorus juglandis. U.S. Patent Application No. 13/548.319, 13 July 2012. [Google Scholar]
- Seybold, S.J.; Haugen, D.; Graves, A. Pest Alert: Thousand Cankers Disease; United States Department of Agriculture, Forest Service, North-Eastern Area State and Private Forestry: Riverdale, MD, USA, 2013; Available online: www.na.fs.fed.us (accessed on 2 September 2022).
- Griffin, G.J. Status of thousand cankers disease on eastern black walnut in the eastern United States at two locations over 3 years. For. Pathol. 2015, 45, 203–214. [Google Scholar] [CrossRef]
- Haun, W.G.; Powell, S.; Strohmeier, C.; Kirksey, J. State of Tennessee Thousand Cankers Disease Action Plan; Tennessee Department of Agriculture, Division of Regulatory Services and Division of Forestry: Nashville, TN, USA, 2010. Available online: http://www.tn.gov/agriculture/publications/regulatory/TN_TCD_ActionPlan.pdf (accessed on 15 October 2022).
- Smallwood, C.J.; Ethington, M.W.; Ginzel, M.D. Managing Thousand Cankers Disease in High-value Plantings of Black Walnut (Fagales: Juglandaceae) in Washington State. J. Integr. Pest Manag. 2022, 13, 5. [Google Scholar] [CrossRef]
- Croll, D.; McDonald, B.A. The genetic basis of local adaptation for pathogenic fungi in agricultural ecosystems. Mol. Ecol. 2017, 26, 2027–2040. [Google Scholar] [CrossRef] [PubMed]
- Carroll, G. Fungal Endophytes in Stems and Leaves: From Latent Pathogen to Mutualistic Symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Ownley, B.H.; Dee, M.M.; Gwinn, K.D. Effect of conidial seed treatment rate of entomopathogenic Beauveria bassiana 11–98 on endophytic colonization of tomato seedlings and control of Rhizoctonia disease. In Proceedings of the 2008 American Phytopathological Society Annual Meeting, Minneapolis, MN, USA, 26–30 July 2008; p. 101. [Google Scholar]
- Lestari, A.; Rao, S. Isolation and pathogenicity of naturally-occurring entomopathogenic fungi to unique bark beetle field crop pest. In Proceedings of the 63rd Annual Meeting of the Entomological Society of America, Minneapolis, MN, USA, 10–13 November 2013. [Google Scholar]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [PubMed]
- Jakobs-Schönwandt, D.; Lohse, R.; Patel, A.V. Cultivation and formulation of an endophytic Beauveria bassiana strain. In Proceedings of the 2011 American Phytopathological Society Annual Meeting, Honolulu, HI, USA, 6–10 August 2011. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control. 2021, 159, 104634. [Google Scholar] [CrossRef]
- Zhang, F.; Huo, Y.; Cobb, A.B.; Luo, G.; Zhou, J.; Yang, G.; Wilson, G.W.; Zhang, Y. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar] [CrossRef] [PubMed]
- Klingeman, W.; Chahal, K.; Gazis, R.; Seybold, S.J.; Graves, A.D.; Windham, M.T.; Hadziabdic, D. Fungi associated with insect galleries and lesions in branches of TCD-impacted walnut and Geosmithia species diversity on field-collected beetles. In Proceedings of the Entomology 2017, Denver, CO, USA, 5–8 November 2017. [Google Scholar]
- Onufrak, A.J.; Williams, G.M.; Klingeman, W.E., III; Cregger, M.A.; Klingeman, D.M.; DeBruyn, J.M.; Ginzel, M.D.; Hadziabdic, D. Regional differences in the structure of Juglans nigra phytobiome reflect geographical differences in thousand cankers disease severity. Phytobiomes J. 2020, 4, 388–404. [Google Scholar] [CrossRef]
- Montecchio, F.; Faccoli, M.; Short, D.P.G.; Fanchin, G.; Geiser, D.M.; Kasson, M.T. First report of Fusarium solani Phylogenetic species 25 associated with early stages of thousand cankers disease of Juglans nigra and Juglans regia in Italy. Plant Dis. 2015, 99, 1183. [Google Scholar] [CrossRef]
- Williams, G.M. Thousand Cankers Disease of Eastern Black Walnut: Ecological Interactions in the Holobiont of a Bark Beetle-Fungal Disease. Ph.D. Thesis, Department of Forestry and Natural Resources West Lafayette, Purdue University, West Lafayette, IN, USA, 2021. [Google Scholar]
- Massey, C.L. Biology and taxonomy of nematode parasites and associates of bark beetles in the United States. In Agricultural Handbook; United States Department of Agriculture: Washington, DC, USA, 1974. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year of Records | Region | Province | Organism Found | Attacked Host | Stand Type * | Mode of Detection ** | Symptom Occurrence *** |
|---|---|---|---|---|---|---|---|
| 2013–2020 | Veneto | Vicenza Treviso Venezia Padova Rovigo Verona | WTB and GM | J. nigra J. regia | Single tree Tree grove Plantation | V + T | Yes |
| 2015–2016 | Piedmont | Turin Novara Cuneo Vercelli | WTB and GM | J. nigra | Single tree Tree row Plantation | V + T | Yes |
| 2015 | Friuli Venezia Giulia | Pordenone | WTB | - | - | T | Not |
| 2015–2016 | Lombardy | Not indicated | WTB and GM | Not indicated | Single tree | V + T | Yes |
| 2018–2022 | Tuscany | Florence | WTB and GM | J. nigra | Single tree Plantation | V + T | Yes |
| 2019 | Emilia Romagna | Reggio Emilia | WTB and GM | J. nigra | Tree row | V | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracalini, M.; Benigno, A.; Aglietti, C.; Panzavolta, T.; Moricca, S. Thousand Cankers Disease in Walnut Trees in Europe: Current Status and Management. Pathogens 2023, 12, 164. https://doi.org/10.3390/pathogens12020164
Bracalini M, Benigno A, Aglietti C, Panzavolta T, Moricca S. Thousand Cankers Disease in Walnut Trees in Europe: Current Status and Management. Pathogens. 2023; 12(2):164. https://doi.org/10.3390/pathogens12020164
Chicago/Turabian StyleBracalini, Matteo, Alessandra Benigno, Chiara Aglietti, Tiziana Panzavolta, and Salvatore Moricca. 2023. "Thousand Cankers Disease in Walnut Trees in Europe: Current Status and Management" Pathogens 12, no. 2: 164. https://doi.org/10.3390/pathogens12020164
APA StyleBracalini, M., Benigno, A., Aglietti, C., Panzavolta, T., & Moricca, S. (2023). Thousand Cankers Disease in Walnut Trees in Europe: Current Status and Management. Pathogens, 12(2), 164. https://doi.org/10.3390/pathogens12020164