


Morphological and Molecular Identification of Dactylogyrus gobiocypris (Monogenea: Dactylogyridae) on Gills of a Model Fish, Gobiocypris rarus (Cypriniformes: Gobionidae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Collection
2.2. Morphological Identification
2.3. DNA extraction, Amplification and Sequencing
2.4. Molecular Analyses
2.5. Histopathology Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite Species | Host Species | Locality | GenBank ID | Refs |
|---|---|---|---|---|
| Dactylogyrus cryptomeres | Gobio gobio | Morava River basin, Czech Republic | AJ564123 | [28] |
| Dactylogyrus finitimus | Romanogobio albipinatus | Morava River basin, Czech Republic | AJ564133 | [28] |
| Dactylogyrus squameus | Pseudorasbora parva | Morava River basin, Czech Republic | AJ564156 | [28] |
| Dactylogyrus gobiocypris | Gobiocypris rarus | Wuhan City, China | OP441417 | Present study |
| Dactylogyrus achmerowi | Cyprinus carpio | Morava River basin, Czech Republic | AJ564108 | [28] |
| Dactylogyrus extensus | Cyprinus carpio | Morava River basin, Czech Republic | AJ564129 | [28] |
| Dactylogyrus vastator | Carassius auratus | Liangzi Lake, China | KC876016 | [37] |
| Dactylogyrus intermedius | Carassius auratus | Liangzi Lake, China | KC876017 | [37] |
| Dactylogyrus lamellatus | Ctenopharyngodon idella | Morava River basin, Czech Republic | AJ564141 | [28] |
| Outgroup | ||||
| Thaparocleidus vistulensis | Silurus glanis | Morava River basin, Czech Republic | AJ490165 | [38] |
3. Results
3.1. Taxonomic Summary
Dactylogyrus gobiocypris Yao, 1995
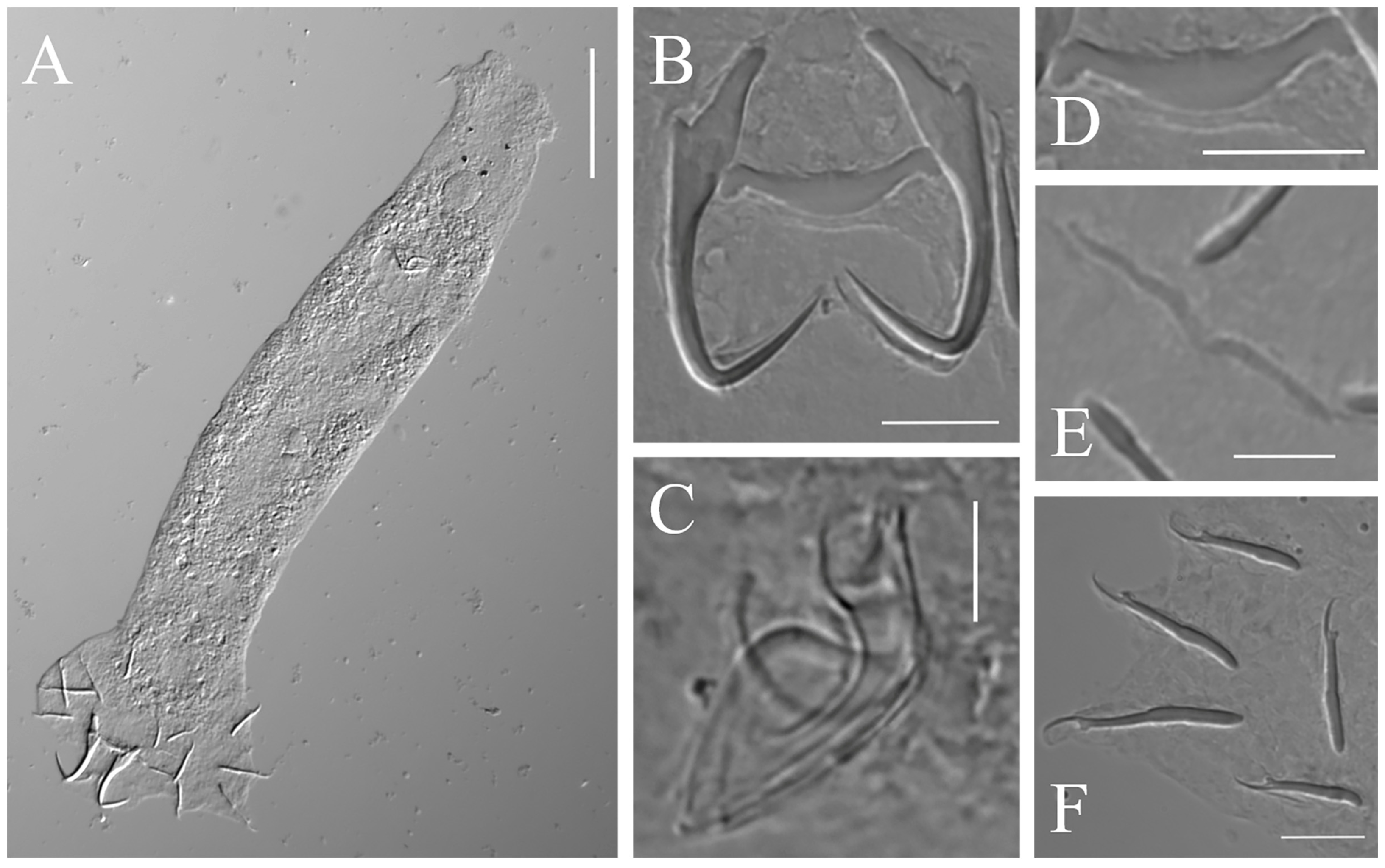
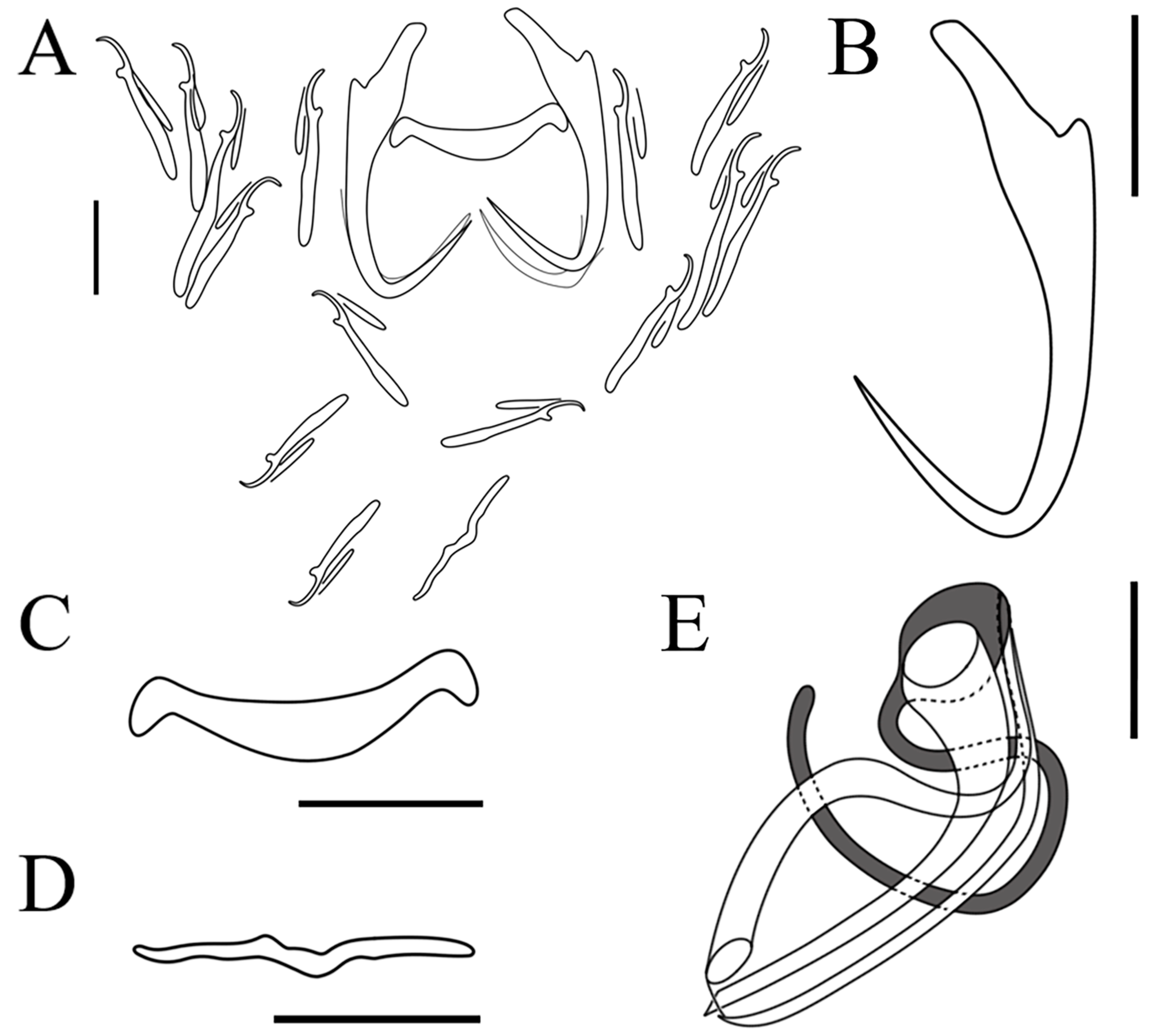
3.2. Morphological Characterization
3.3. Molecular Analyses
3.4. Histopathology Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, M.; Fu, T. Description of a new genus and species of Danioninae from China (Cypriniformes: Cyprinidae). Acta Zootaxonomica Sin. 1983, 8, 434–437. [Google Scholar]
- He, Y.; Wang, J.; Blanchet, S.; Lek, S. Genetic structure of an endangered endemic fish (Gobiocypris rarus) in the upper Yangtze River. Biochem. Syst. Ecol. 2012, 43, 214–225. [Google Scholar] [CrossRef]
- Zhou, Y.; Cheng, S.; Hu, W.; Sun, M. A new toxicity test organism-Gobiocypris rarus. Zool. Res. 1995, 16, 59. [Google Scholar]
- Zhong, X.; Xu, Y.; Liang, Y.; Liao, T.; Wang, J. The Chinese rare minnow (Gobiocypris rarus) as an in vivo model for endocrine disruption in freshwater teleosts: A full life-cycle test with diethylstilbestrol. Aquat. Toxicol. 2005, 71, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, W. Gobiocypris rarus as a Chinese native model organism: History and current situation. Asian J. Ecotoxicol. 2017, 12, 20–33. [Google Scholar] [CrossRef]
- Li, Z.; Chen, L.; Wu, Y.; Li, P.; Li, Y.; Ni, Z. Effects of mercury on oxidative stress and gene expression of potential biomarkers in larvae of the Chinese rare minnow Gobiocypris rarus. Arch. Environ. Contam. Toxicol. 2014, 67, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, L.; Li, D.; Ling, F.; Wang, G. Developmental toxicity in rare minnow (Gobiocypris rarus) embryos exposed to Cu, Zn and Cd. Ecotoxicol. Environ. Saf. 2014, 104, 269–277. [Google Scholar] [CrossRef]
- Wu, W.; Li, W.; Xu, Y.; Wang, J. Long-term toxic impact of 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin on the reproduction, sexual differentiation, and development of different life stages of Gobiocypris rarus and Daphnia magna. Ecotoxicol. Environ. Saf. 2001, 48, 293–300. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, T.; Hu, X.; Wang, G. Developmental toxicity of 3, 4-dichloroaniline on rare minnow (Gobiocypris rarus) embryos and larvae. Chemosphere 2013, 90, 1132–1139. [Google Scholar] [CrossRef]
- Yan, S.; Wang, J.; Zheng, Z.; Ji, F.; Yan, L.; Yang, L.; Zha, J. Environmentally relevant concentrations of benzophenones triggered DNA damage and apoptosis in male Chinese rare minnows (Gobiocypris rarus). Environ. Int. 2022, 164, 107260. [Google Scholar] [CrossRef]
- Zha, J.; Sun, L.; Spear, P.A.; Wang, Z. Comparison of ethinylestradiol and nonylphenol effects on reproduction of Chinese rare minnows (Gobiocypris rarus). Ecotoxicol. Environ. Saf. 2008, 71, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, Y.; Wang, J.; Tao, Y.; Dai, J. Toxicogenomic analysis of the hepatic effects of perfluorooctanoic acid on rare minnows (Gobiocypris rarus). Toxicol. Appl. Pharmacol. 2008, 226, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, L.; Zhang, G.; Guan, Y.; Wang, Z. Effect of low-dose malathion on the gonadal development of adult rare minnow Gobiocypris rarus. Ecotoxicol. Environ. Saf. 2016, 125, 135–140. [Google Scholar] [CrossRef]
- Gibson, D.I.; Timofeeva, T.A.; Gerasev, P.I. A catalogue of the nominal species of the monogenean genus Dactylogyrus Diesing, 1850 and their host genera. Syst. Parasitol. 1996, 35, 3–48. [Google Scholar] [CrossRef]
- Wu, B.; Lang, S.; Wang, J. Fauna sinica. Platyhelminthes. Monogenea; Science Press: Beijing, China, 2000; pp. 222–268. [Google Scholar]
- Pugachev, O.; Gerasev, P.; Gussev, A.; Ergens, R.; Khotenowsky, I. Guide to Monogenoidea of Freshwater Fish of Palaearctic and Amur Regions; Ledizione-LediPublishing: Milan, Italy, 2009; pp. 398–525. [Google Scholar]
- Yao, W. A new species of Dactylogyrus parasitic from gills of Gobiocypris rarus. Trans. Res. Fish Dis. 1995, 2, 119–120. [Google Scholar]
- Wierzbicka, J. Monogenoidea of gills of certain Cyprinidae fish species. Acta Parasitol. Pol. 1974, 22, 149–163. [Google Scholar]
- Jalali, B.; Barzegar, M. Dactylogyrids (Dactylogyridae: Monogenea) on common carp (Cyprinus carpio L.) in freshwaters of Iran and description of the pathogenicity of D. sahuensis. J. Agric. Sci. Technol. 2005, 7, 9–16. [Google Scholar]
- Rahanandeh, M.; Sharifpour, I.; Jalali, B.; Kazemi, R.; Fatideh, B.; Sabet, S. Survey on Dactylogyrosis in Caspian frisian roach (Rutilus frisii kutum) caused by Dactylogyrus frisii. Glob. Vet. 2010, 4, 515–518. [Google Scholar]
- Reed, P.; Francis-Floyd, R.; Klinger, R.; Petty, D. Monogenean parasites of fish. In Publication Series of Fisheries and Aquatic Sciences Department, Institute of Food and Agricultural Sciences, University of Florida; University of Florida: Gainesville, FL, USA, 2009; pp. 1–10. [Google Scholar]
- Tu, X.; Ling, F.; Huang, A.; Wang, G. The first report of Dactylogyrus formosus Kulwiec, 1927 (Monogenea: Dactylogyridae) from goldfish (Carassius auratus) in central China. Parasitol. Res. 2015, 114, 2689–2696. [Google Scholar] [CrossRef]
- Dove, A.D.; Ernst, I. Concurrent invaders-four exotic species of Monogenea now established on exotic freshwater fishes in Australia. Int. J. Parasitol. 1998, 28, 1755–1764. [Google Scholar] [CrossRef]
- Ergens, R. The suitability of ammonium picrate-glycerin in preparing slides of lower Monogenoidea. Folia Parasitol. 1969, 16, 320. [Google Scholar]
- Malmberg, G. The excretory systems and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Ark. För Zool. 1970, 23, 1–235. [Google Scholar]
- Paladini, G.; Gustinelli, A.; Fioravanti, M.L.; Hansen, H.; Shinn, A.P. The first report of Gyrodactylus salaris Malmberg, 1957 (Platyhelminthes, Monogenea) on Italian cultured stocks of rainbow trout (Oncorhynchus mykiss Walbaum). Vet. Parasitol. 2009, 165, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimková, A.; Pečínková, M.; Řehulková, E.; Vyskočilová, M.; Ondračková, M. Dactylogyrus species parasitizing European Barbus species: Morphometric and molecular variability. Parasitology 2007, 134, 1751–1765. [Google Scholar] [CrossRef] [PubMed]
- Šimková, A.; Morand, S.; Jobet, E.; Gelnar, M.; Verneau, O. Molecular phylogeny of congeneric monogenean parasites (Dactylogyrus): A case of intrahost speciation. Evolution 2004, 58, 1001–1018. [Google Scholar] [CrossRef]
- Sharma, P.; Agarwal, N.; Kumar, S. Ribosomal DNA and morphological analysis of Dactylogyrus species from freshwater fishes of India. J. Parasit. Dis. 2011, 35, 210–214. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.; Wang, G. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, K. Studies on gill parasitosis of the grasscarp (Ctenopharyngodon idella) caused by Dactylogyrus lamellatus Achmerov, 1952. Acta Vet. Acad. Sci. Hung. 1972, 22, 9–24. [Google Scholar] [PubMed]
- Cheng, J.; Pan, Y.; Ma, X.; Wang, G.; Li, W. Morphological and molecular identification of 7 species of Dactylogyrus on gills of the goldfish (Carassius auratus). Acta Hydrobiol. Sin. 2023, 47, 345–354. [Google Scholar]
- Šimková, A.; Plaisance, L.; Matějusová, I.; Morand, S.; Verneau, O. Phylogenetic relationships of the Dactylogyridae Bychowsky, 1933 (Monogenea: Dactylogyridea): The need for the systematic revision of the Ancyrocephalinae Bychowsky, 1937. Syst. Parasitol. 2003, 54, 1–11. [Google Scholar] [CrossRef]
- Gusev, A. Monogenetic trematodes of fish from the Amur River system. Tr. Zool. Inst. Akad. Nauk. SSSR 1955, 19, 171–398. [Google Scholar]
- Zolovs, M.; Ozuna, A.; Kirjušina, M. Seasonal variation of attachment apparatus and copulatory organ morphometric variables of Dactylogyrus crucifer Wagener, 1857 (Monogenea: Dactylogyridae) on the gills of roach (Rutilus rutilus L.) in Latvian water bodies. Acta Biol. Univ. Daugavp. 2012, 12, 191–198. [Google Scholar]
- Vignon, M.; Sasal, P. The use of geometric morphometrics in understanding shape variability of sclerotized haptoral structures of monogeneans (Platyhelminthes) with insights into biogeographic variability. Parasitol. Int. 2010, 59, 183–191. [Google Scholar] [CrossRef]
- Ling, F.; Tu, X.; Huang, A.; Wang, G. Morphometric and molecular characterization of Dactylogyrus vastator and D. intermedius in goldfish (Carassius auratus). Parasitol. Res. 2016, 115, 1755–1765. [Google Scholar] [CrossRef]
- Simkova, A.; Morand, S. Co-evolutionary patterns in congeneric monogeneans: A review of Dactylogyrus species and their cyprinid hosts. J. Fish Biol. 2008, 73, 2210–2227. [Google Scholar] [CrossRef]
- Li, W.X.; Zou, H.; Wu, S.G.; Xiong, F.; Li, M.; Ma, X.R.; Marcogliese, D.J.; Locke, S.A.; Wang, G.T. Composition and diversity of communities of Dactylogyrus spp. in wild and farmed goldfish Carassius auratus. J. Parasitol. 2018, 104, 353–358. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Wang, J. Genetic structure of IHB rare minnow (Gobiocypris rarus). J. Hydroecol. 2018, 39, 83–90. [Google Scholar]
- Zhang, C.; Li, D.; Chi, C.; Ling, F.; Wang, G. Dactylogyrus intermedius parasitism enhances Flavobacterium columnare invasion and alters immune-related gene expression in Carassius auratus. Dis. Aquat. Org. 2015, 116, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Horoszewicz, L. The influence of parasites, handling of fish and the methods of investigations on the evaluation of their tolerance and thermal resistance. Rocz Nauk. Roln 1972, 94, 35–53. [Google Scholar]
- Sindermann, C.J. Pollution-associated diseases and abnormalities of fish and shellfish: A review. Fish. Bull. 1979, 76, 717–749. [Google Scholar]
- Su, L.; Xu, C.; Cai, L.; Qiu, N.; Hou, M.; Wang, J. Susceptibility and immune responses after challenge with Flavobacterium columnare and Pseudomonas fluorescens in conventional and specific pathogen-free rare minnow (Gobiocypris rarus). Fish Shellfish. Immunol. 2020, 98, 875–886. [Google Scholar] [CrossRef]
- Perevozchenko, I.; Davydov, C. DDT and its metabolites in some cestodes in fishes. Gidrobiol. Zhurnal 1974, 10, 86–90. [Google Scholar]
- Boyce, N.P.; Yamada, S.B. Effects of a parasite, Eubothrium salvelini (Cestoda: Pseudophyllidea), on the resistance of juvenile sockeye salmon, Oncorhynchus nerka, to Zinc. J. Fish. Board Can. 1977, 34, 706–709. [Google Scholar] [CrossRef]
- Pascoe, D.; Cram, P. The effect of parasitism on the toxicity of cadmium to the three-spined stickleback, Gasterosteus aculeatus L. J. Fish Biol. 1977, 10, 467–472. [Google Scholar] [CrossRef]
- Moles, A. Sensitivity of parasitized coho salmon fry to crude oil, toluene, and naphthalene. Trans. Am. Fish. Soc. 1980, 109, 293–297. [Google Scholar] [CrossRef]






| Source of Data | N | Dactylogyrus gobiocypris | Dactylogyrus trullaeformis | |
|---|---|---|---|---|
| Gobiocypris rarus | G. rarus | Gnathopogon strigatus | ||
| Squalidus chankaensis | ||||
| this Study | Yao, 1995 [17] (n = 7) | Gussev, 1955 [39] | ||
| Body | ||||
| Total length | 32 | 182.0 ± 34.6 (117.7–248.4) | 102.5–113.0 | 150–300 |
| Total width | 32 | 44.8 ± 9.6 (27.8–74.1) | 27.5–28.1 | 30–50 |
| Pharynx diameter | 33 | 11.2 ± 2.2 (7.2–16.8) | 34.5 | 16–19 |
| Anchor | ||||
| Total length | 55 | 26.4 ± 1.6 (23.8–30.1) | 26.5–30.0 | 27–30 |
| Base length | 55 | 21.1 ± 1.5 (18.2–24.3) | 21–24 | |
| Outer root length | 55 | 1.0 ± 0.2 (0.6–1.9) | 1.5 | 1–2 |
| Inner root length | 55 | 7.2 ± 0.7 (5.9–9.0) | 7.5–10.0 | 6–8 |
| Point length | 55 | 11.5 ± 0.8 (9.5–13.4) | 12.5–13.0 | 11–13 |
| Ventral bar | ||||
| Total length | 56 | 3.6 ± 0.7 (2.4–5.7) | ||
| Medium length | 56 | 2.3 ± 0.6 (1.5–4.6) | 1.8–3.4 | 1 |
| Width | 56 | 17.9 ± 1.4 (14.8–21.0) | 17.5–20.0 | 14–16 |
| Dorsal bar | ||||
| Total length | 52 | 2.4 ± 0.4 (1.5–3.5) | ||
| Medium length | 52 | 1.0 ± 0.2 (0.5–1.5) | 1.0 | 2 |
| Width | 52 | 15.8 ± 1.6 (13.0–20.6) | 11.0–15.0 | 10–19 |
| Marginal hook | ||||
| Total length | 55 | 17.1 ± 1.5 (14.3–22.8) | 16.5–25.0 | 15–23 |
| Sickle length | 55 | 5.0 ± 0.4 (4.0–6.3) | ||
| Shaft length | 55 | 12.1 ± 1.5 (9.3–17.7) | ||
| Filament loop length | 55 | 7.7 ± 0.5 (6.9–9.0) | ||
| Copulatory complex | ||||
| Penis length | 30 | 11.9 ± 1.2 (10.3–14.4) | 15.0–20.0 | 16–21 |
| Accessory piece | 30 | 15.5 ± 1.1 (13.4–17.1) | 17.5–21.3 | 17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, J.; Zou, H.; Li, M.; Wang, J.; Wang, G.; Li, W. Morphological and Molecular Identification of Dactylogyrus gobiocypris (Monogenea: Dactylogyridae) on Gills of a Model Fish, Gobiocypris rarus (Cypriniformes: Gobionidae). Pathogens 2023, 12, 206. https://doi.org/10.3390/pathogens12020206
Cheng J, Zou H, Li M, Wang J, Wang G, Li W. Morphological and Molecular Identification of Dactylogyrus gobiocypris (Monogenea: Dactylogyridae) on Gills of a Model Fish, Gobiocypris rarus (Cypriniformes: Gobionidae). Pathogens. 2023; 12(2):206. https://doi.org/10.3390/pathogens12020206
Chicago/Turabian StyleCheng, Jiangwen, Hong Zou, Ming Li, Jianwei Wang, Guitang Wang, and Wenxiang Li. 2023. "Morphological and Molecular Identification of Dactylogyrus gobiocypris (Monogenea: Dactylogyridae) on Gills of a Model Fish, Gobiocypris rarus (Cypriniformes: Gobionidae)" Pathogens 12, no. 2: 206. https://doi.org/10.3390/pathogens12020206
APA StyleCheng, J., Zou, H., Li, M., Wang, J., Wang, G., & Li, W. (2023). Morphological and Molecular Identification of Dactylogyrus gobiocypris (Monogenea: Dactylogyridae) on Gills of a Model Fish, Gobiocypris rarus (Cypriniformes: Gobionidae). Pathogens, 12(2), 206. https://doi.org/10.3390/pathogens12020206