


Presence of Anaplasma phagocytophilum Ecotype I in UK Ruminants and Associated Zoonotic Risk

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and DNA Extraction
2.2. PCR for the Amplification and Sequencing of the groEL Gene
2.3. Nucleotide Sequence Analysis
2.4. Phylogenetic Analysis
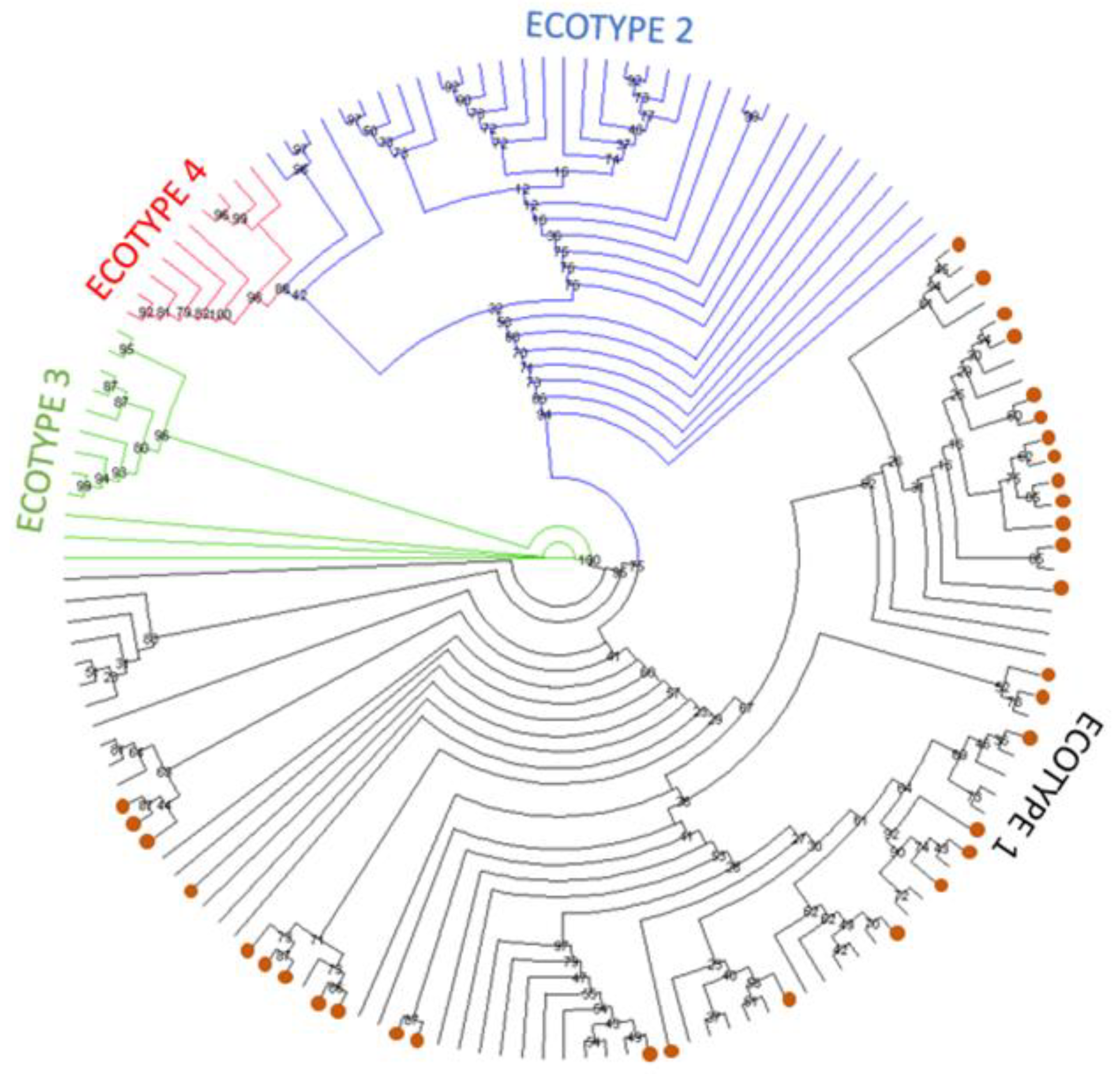
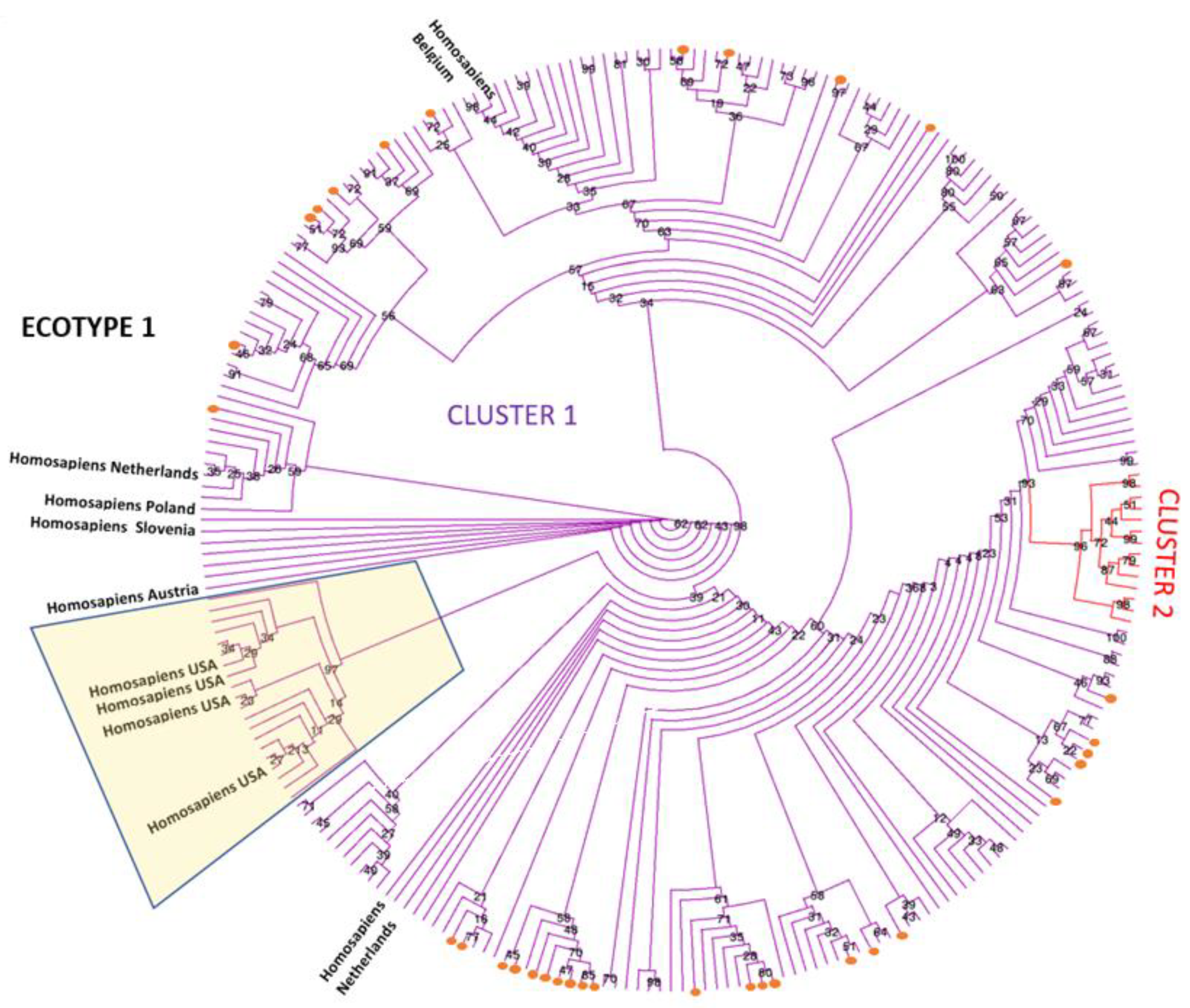
3. Results
3.1. Primer Selection and groEL PCR Assays on Ruminant Samples
3.2. Sequence Analysis
3.3. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging tick-borne diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef] [PubMed]
- Remesar, S.; Díaz, P.; Prieto, A.; García-Dios, D.; Fernández, G.; López, C.M.; Panadero, R.; Díez-Baños, P.; Morrondo, P. Prevalence and molecular characterization of Anaplasma phagocytophilum in roe deer (Capreolus capreolus) from Spain. Ticks Tick Borne Dis. 2020, 11, 101351. [Google Scholar] [CrossRef] [PubMed]
- Rikihisa, Y. Mechanism of obligatory intracellular infection with Anaplasma phagocytophilum. Clin. Microbiol. Rev. 2011, 24, 469–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum-a widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Keesing, F.; Hersh, M.H.; Tibbetts, M.; McHenry, D.J.; Duerr, S.; Brunner, J.; Killilea, M.; LoGiudice, K.; Schmidt, K.A.; Ostfeld, R.S. Reservoir competence of vertebrate hosts for Anaplasma phagocytophilum. Emerg. Infect. Dis. 2012, 18, 2013–2016. [Google Scholar] [CrossRef]
- Atif, F.A. Anaplasma marginale and Anaplasma phagocytophilum: Rickettsiales pathogens of veterinary and public health significance. Parasitol. Res. 2015, 114, 3941–3957. [Google Scholar] [CrossRef]
- Rar, V.; Tkachev, S.; Tikunova, N. Genetic diversity of Anaplasma bacteria: Twenty years later. Infect. Genet. Evol. 2021, 91, 104833. [Google Scholar] [CrossRef]
- Woldehiwet, Z. Anaplasma phagocytophilum in ruminants in Europe. Ann. N. Y. Acad. Sci. 2006, 1078, 446–460. [Google Scholar] [CrossRef]
- Hauck, D.; Jordan, D.; Springer, A.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasites Vectors 2020, 13, 176. [Google Scholar] [CrossRef] [Green Version]
- Morissette, E.; Massung, R.F.; Foley, J.E.; Alleman, A.R.; Foley, P.; Barbet, A.F. Diversity of Anaplasma phagocytophilum strains, USA. Emerg. Infect. Dis. 2009, 15, 928–931. [Google Scholar] [CrossRef]
- Foley, J.; Nieto, N.C.; Madigan, J.; Sykes, J. Possible differential host tropism in Anaplasma phagocytophilum strains in the Western United State. Ann. N. Y. Acad. Sci. 2008, 1149, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Bown, K.J.; Lambin, X.; Ogden, N.H.; Begon, M.; Telford, G.; Woldehiwet, Z.; Birtles, R.J. Delineating Anaplasma phagocytophilum ecotypes in coexisting, discrete enzootic cycles. Emerg. Infect. Dis. 2009, 15, 1948–1954. [Google Scholar] [CrossRef]
- Dugat, T.; Lagrée, A.; Maillard, R.; Boulouis, H.; Haddad, N. Opening the black box of Anaplasma phagocytophilum diversity: Current situation and future perspectives. Front. Cell. Infect. Microbiol. 2015, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; Maanen, C.V.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasites Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaarsma, R.I.; Sprong, H.; Takumi, K.; Kazimirova, M.; Silaghi, C.; Mysterud, A.; Rudolf, I.; Beck, R.; Földvári, G.; Tomassone, L.; et al. Anaplasma phagocytophilum evolves in geographical and biotic niches of vertebrates and ticks. Parasites Vectors 2019, 12, 328. [Google Scholar] [CrossRef]
- Grassi, L.; Franzo, G.; Martini, M.; Mondin, A.; Cassini, R.; Drigo, M.; Pasotto, D.; Vidorin, E.; Menandro, M.L. Ecotyping of Anaplasma phagocytophilum from wild ungulates and ticks shows circulation of zoonotic strains in northeastern Italy. Animals 2021, 11, 310. [Google Scholar] [CrossRef]
- Katargina, O.; Geller, J.; Alekseev, A.; Dubinina, H.; Efremova, G.; Mishaeva, N.; Vasilenko, V.; Kuznetsova, T.; Järvekülg, L.; Vene, S.; et al. Identification of Anaplasma phagocytophilum in tick populations in Estonia, the European part of Russia and Belarus. Clin. Microbiol. Infect. 2012, 18, 40–46. [Google Scholar] [CrossRef] [Green Version]
- von Loewenich, F.D.; Baumgarten, B.U.; Schröppel, K.; Geissdörfer, W.; Röllinghoff, M.; Bogdan, C. High diversity of ankA sequences of Anaplasma phagocytophilum among Ixodes ricinus ticks in Germany. J. Clin. Microbiol. 2003, 41, 5033–5040. [Google Scholar] [CrossRef] [Green Version]
- Langenwalder, D.B.; Schmidt, S.; Silaghi, C.; Skuballa, J.; Pantchev, N.; Matei, I.A.; Mihalca, A.D.; Gilli, U.; Zajkowska, J.; Ganter, M.; et al. The absence of the drhm gene is not a marker for human-pathogenicity in European Anaplasma phagocytophilum strains. Parasites Vectors 2020, 13, 238. [Google Scholar] [CrossRef]
- Aardema, M.L.; von Loewenich, F.D. Varying influences of selection and demography in host-adapted populations of the tick-transmitted bacterium, Anaplasma phagocytophilum. BMC Evol. Biol. 2015, 15, 58. [Google Scholar] [CrossRef]
- Aardema, M.L.; Bates, N.V.; Archer, Q.E.; von Loewenich, F.D. Demographic expansions and the emergence of host specialization in genetically distinct ecotypes of the tick-transmitted bacterium Anaplasma phagocytophilum. Appl. Environ. Microbiol. 2022, 88, e0061722. [Google Scholar] [CrossRef] [PubMed]
- Gandy, S.; Hansford, K.; McGinley, L.; Cull, B.; Smith, R.; Semper, A.; Brooks, T.; Fonville, M.; Sprong, H.; Phipps, P.; et al. Prevalence of Anaplasma phagocytophilum in questing Ixodes ricinus nymphs across twenty recreational areas in England and Wales. Ticks Tick Borne Dis. 2022, 13, 101965. [Google Scholar] [CrossRef] [PubMed]
- Olsthoorn, F.; Sprong, H.; Fonville, M.; Rocchi, M.; Medlock, J.; Gilbert, L.; Ghazoul, J. Occurrence of tick-borne pathogens in questing Ixodes ricinus ticks from Wester Ross, Northwest Scotland. Parasites Vectors 2021, 14, 430. [Google Scholar] [CrossRef] [PubMed]
- Occhibove, F.; McKeown, N.J.; Risley, C.; Ironside, J.E. Eco-epidemiological screening of multi-host wild rodent communities in the UK reveals pathogen strains of zoonotic interest. Int. J. Parasitol. Parasites Wildl. 2022, 17, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, M.; Dagleish, M.; McInnes, C. Tick bites and tick-transmitted diseases. Vet. Rec. 2018, 182, 609. [Google Scholar] [CrossRef]
- Daniel, R.; Hopkins, B.A.M.; Rocchi, M.S.; Wessels, M.; Floyd, T. High mortality in a sheep flock caused by coinfection of coinfection of louping ill virus and Anaplasma phagocytophilum. Vet. Rec. Case Rep. 2020, 8, e000980. [Google Scholar] [CrossRef]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [Green Version]
- Woldehiwet, Z.; Horrocks, B.K. Antigenicity of ovine strains of Anaplasma phagocytophilum grown in tick cells and ovine granulocytes. J. Comp. Pathol. 2005, 132, 322–328. [Google Scholar] [CrossRef]
- Alberti, A.; Addis, M.F.; Sparagano, O.; Zobba, R.; Chessa, B.; Cubeddu, T.; Parpaglia, M.L.; Ardu, M.; Pittau, M. Anaplasma phagocytophilum, Sardinia, Italy. Emerg. Infect. Dis. 2005, 11, 1322–1324. [Google Scholar] [CrossRef]
- IUPAC-IUB Comm. on Biochem. Nomenclature (CBN). Abbreviations and symbols for nucleic acids, polynucleotides, and their constituents. Biochemistry 1970, 9, 4022–4027. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.F.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavaré, S.; Miura, R.M. Some probabilistic and statistical problems in the analysis of DNA sequences. Lectures on mathematics in the life sciences. Am. Math. Soc. 1986, 17, 57–86. [Google Scholar]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3- An interactive viewer for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Jouglin, M.; Chagneau, S.; Faille, F.; Verheyden, H.; Bastian, S.; Malandrin, L. Detecting and characterizing mixed infections with genetic variants of Anaplasma phagocytophilum in roe deer (Capreolus capreolus) by developing an ankA cluster-specific nest-ed PCR. Parasites Vectors 2017, 10, 377. [Google Scholar] [CrossRef]
- Crosby, F.L.; Eskeland, S.; Bø-Granquist, E.G.; Munderloh, U.G.; Price, L.D.; Al-Khedery, B.; Stuen, S.; Barbet, A.F. Comparative Whole Genome Analysis of an Anaplasma phagocytophilum Strain Isolated from Norwegian Sheep. Pathogens 2022, 11, 601. [Google Scholar] [CrossRef]
- Zeilstra-Ryalls, J.; Fayet, O.; Georgopoulos, C. The universally conserved GroE (Hsp60) chaperonins. Annu. Rev. Microbiol. 1991, 45, 301–325. [Google Scholar] [CrossRef]
- Villar, M.; Ayllón, N.; Kocan, K.M.; Bonzón-Kulichenko, E.; Alberdi, P.; Blouin, E.F.; Weisheit, S.; Mateos-Hernández, L.; Cabezas-Cruz, A.; Bell-Sakyi, L.; et al. Identification and Characterization of Anaplasma phagocytophilum proteins involved in infection of the tick vector, Ixodes scapularis. PLoS ONE 2015, 10, e0137237. [Google Scholar] [CrossRef]
- Stuen, S.; Bergström, K. Serological investigation of granulocytic Ehrlichia infection in sheep in Norway. Acta Vet. Scand. 2001, 42, 331–338. [Google Scholar] [CrossRef]
- Hamšíková, Z.; Silaghi, C.; Takumi, K.; Rudolf, I.; Gunár, K.; Sprong, H.; Kazimírová, M. Presence of roe deer affects the occurrence of Anaplasma phagocytophilum ecotypes in questing Ixodes ricinus in different habitat types of Central Europe. Int. J. Environ. Res. Public Health 2019, 16, 4725. [Google Scholar] [CrossRef] [PubMed]
- Stigum, V.M.; Jaarsma, R.I.; Sprong, H.; Rolandsen, C.M.; Mysterud, A. Infection prevalence and ecotypes of Anaplasma phagocytophilum in moose Alces alces, red deer Cervus elaphus, roe deer Capreolus capreolus, and Ixodes ricinus ticks from Norway. Parasites Vectors 2019, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Dwużnik-Szarek, D.; Kowalec, M.; Alsarraf, M.; Bajer, A. Contribution of tick-borne diseases to mortality in juvenile free-living cervids. Ann. Agric. Environ. Med. 2022, 29, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Myczka, A.W.; Kaczor, S.; Filip-Hutsch, K.; Czopowicz, M.; Plis-Kuprianowicz, E.; Laskowski, Z. Prevalence and genotyping of Anaplasma phagocytophilum strains from wild animals, European Bison (Bison bonasus) and Eurasian Moose (Alces alces) in Poland. Animals 2022, 12, 1222. [Google Scholar] [CrossRef] [PubMed]
- Lesiczka, P.M.; Modry, D.; Sprong, H.; Fonville, M.; Pikula, J.; Piacek, V.; Heger, T.; Hrazdilova, K. Detection of Anaplasma phagocytophilum in European brown hares (Lepus europaeus) using three different methods. Zoonoses Public Health 2021, 68, 917–925. [Google Scholar] [CrossRef]
- Dugat, T.; Leblond, A.; Keck, N.; Lagrée, A.C.; Desjardins, I.; Joulié, A.; Pradier, S.; Durand, B.; Boulouis, H.J.; Haddad, N. One particular Anaplasma phagocytophilum ecotype infects cattle in the Camargue, France. Parasites Vectors 2017, 10, 371. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Guan, G.; Li, Y.; Liu, G.; Luo, J.; Yin, H. Anaplasma phagocytophilum in sheep and goats in central and southeastern China. Parasites Vectors 2016, 9, 593. [Google Scholar] [CrossRef] [Green Version]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Bakken, J.S.; Dumler, J.S. Human granulocytic anaplasmosis. Infect. Dis. Clin. N. Am. 2015, 29, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Dumler, J.S.; Dotevall, L.; Gustafson, R.; Granström, M. A population-based seroepidemiologic study of human granulocytic ehrlichiosis and Lyme borreliosis on the west coast of Sweden. J. Infect. Dis. 1997, 175, 720–722. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ecotype | Cluster | Haplotypes (n) | Hosts | Vectors | Regions |
|---|---|---|---|---|---|
| Ecotype I | 1 | 1095 | Artiodactyla, Perisodactyla, Aves, Carnivora, Rodentia, Erinaceomorpha, Primates, Lagomorpha | Ixodidae Lipoptena cervi | Europe, Central America, and North America |
| 2 | 1 | Artiodactyla | Unknown | Europe | |
| Ecotype II | 3 | 253 | Artiodactyla, Carnivora, Rodentia | Ixodidae Lipoptena cervi | Asia, Europe, and South America |
| 4 | 31 | Artiodactyla, Carnivora, non-human Primates, Rodentia | Ixodidae | Asia, Europe | |
| Ecotype III | 5 | 117 | Rodentia, Soricomorpha | Ixodidae | Europe |
| 6 | 15 | Rodentia, | Ixodidae | Europe | |
| Ecotype IV | 7 | 2 | Aves, Erinaceomorpha, Rodentia | Ixodidae | Europe |
| 8 | 5 | unknown | I. ventalloi | Europe |
| Species | Matrix | Total Number | Date Range |
|---|---|---|---|
| Ovine | Tissue | 43 | May 2020–Aug 2022 |
| Ovine | EDTA blood | 19 | Jan 2022–Aug 2022 |
| Bovine | Tissues | 3 | Jul 2020–Jun 2022 |
| Bovine | EDTA blood | 2 | Jun 2022–Jul 2022 |
| Cervine | EDTA blood | 3 | Feb 2022–Jun 2022 |
| Caprine | Heparinised blood | 1 | Aug 2022 |
| Originally isolated from caprine | Infected tick cell line APFG | 1 | May 2015 |
| Primer Name | Sequence | Specificity | Reference |
|---|---|---|---|
| groEL 569 F | ATGGTATGCAGTTTGATCGC | A. platys, A. bovis, Ehrlichia sp. | [26] |
| groEL nd643 F | ACTGATGGTATGCAGTTTGATCG | A. phagocytophilum | New |
| groEL 1236 R | TCTTTRCGTTCYTTMACYTCAACTTC | A. platys, A. bovis, Ehrlichia sp., Lentilitoribacter sp., Neorickettsia findlayensis, Rhodobiaceae bacterium, Erythrobacter sp., Cohaesibacter sp. | [15] |
| groEl nd1236 R | TCTTTGCGTTCCTTCACCTCAACTTC | A. phagocytophilum | New |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchessi, L.; Rocchi, M.S.; Maley, M.; Allen, K.; Ballingall, K.; Turin, L. Presence of Anaplasma phagocytophilum Ecotype I in UK Ruminants and Associated Zoonotic Risk. Pathogens 2023, 12, 216. https://doi.org/10.3390/pathogens12020216
Bianchessi L, Rocchi MS, Maley M, Allen K, Ballingall K, Turin L. Presence of Anaplasma phagocytophilum Ecotype I in UK Ruminants and Associated Zoonotic Risk. Pathogens. 2023; 12(2):216. https://doi.org/10.3390/pathogens12020216
Chicago/Turabian StyleBianchessi, Laura, Mara Silvia Rocchi, Madeleine Maley, Kayleigh Allen, Keith Ballingall, and Lauretta Turin. 2023. "Presence of Anaplasma phagocytophilum Ecotype I in UK Ruminants and Associated Zoonotic Risk" Pathogens 12, no. 2: 216. https://doi.org/10.3390/pathogens12020216
APA StyleBianchessi, L., Rocchi, M. S., Maley, M., Allen, K., Ballingall, K., & Turin, L. (2023). Presence of Anaplasma phagocytophilum Ecotype I in UK Ruminants and Associated Zoonotic Risk. Pathogens, 12(2), 216. https://doi.org/10.3390/pathogens12020216