

Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification of Virus-Related Sequences from Yellow Leaf Samples of Wheat Plants via RNA-Seq
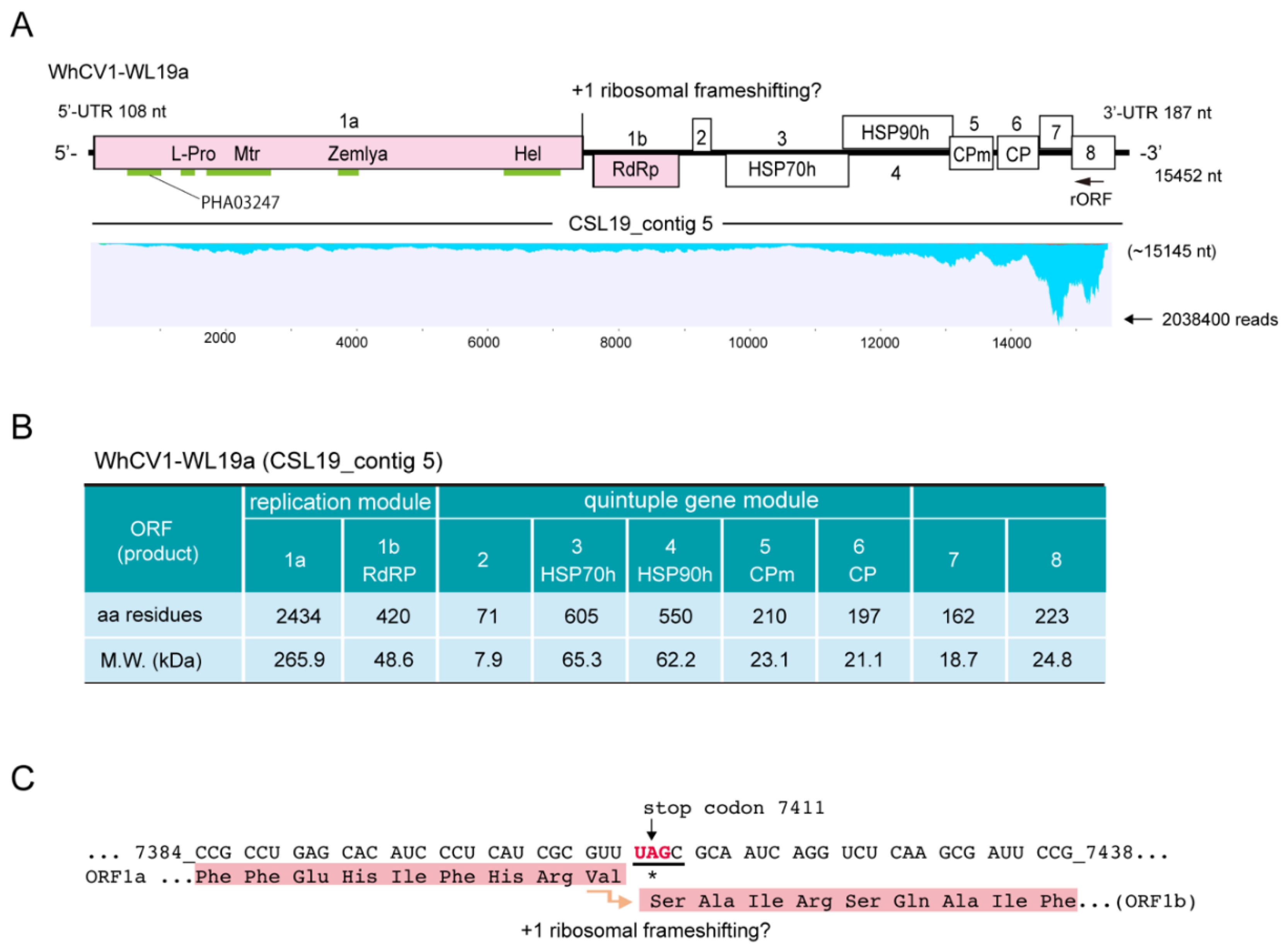
2.2. Complete Genome Sequence of Closterovirus from the Wheat Yellow Leaf Sample
2.3. Sequence Characteristics of the Predicted WhCV1-Encoded Proteins
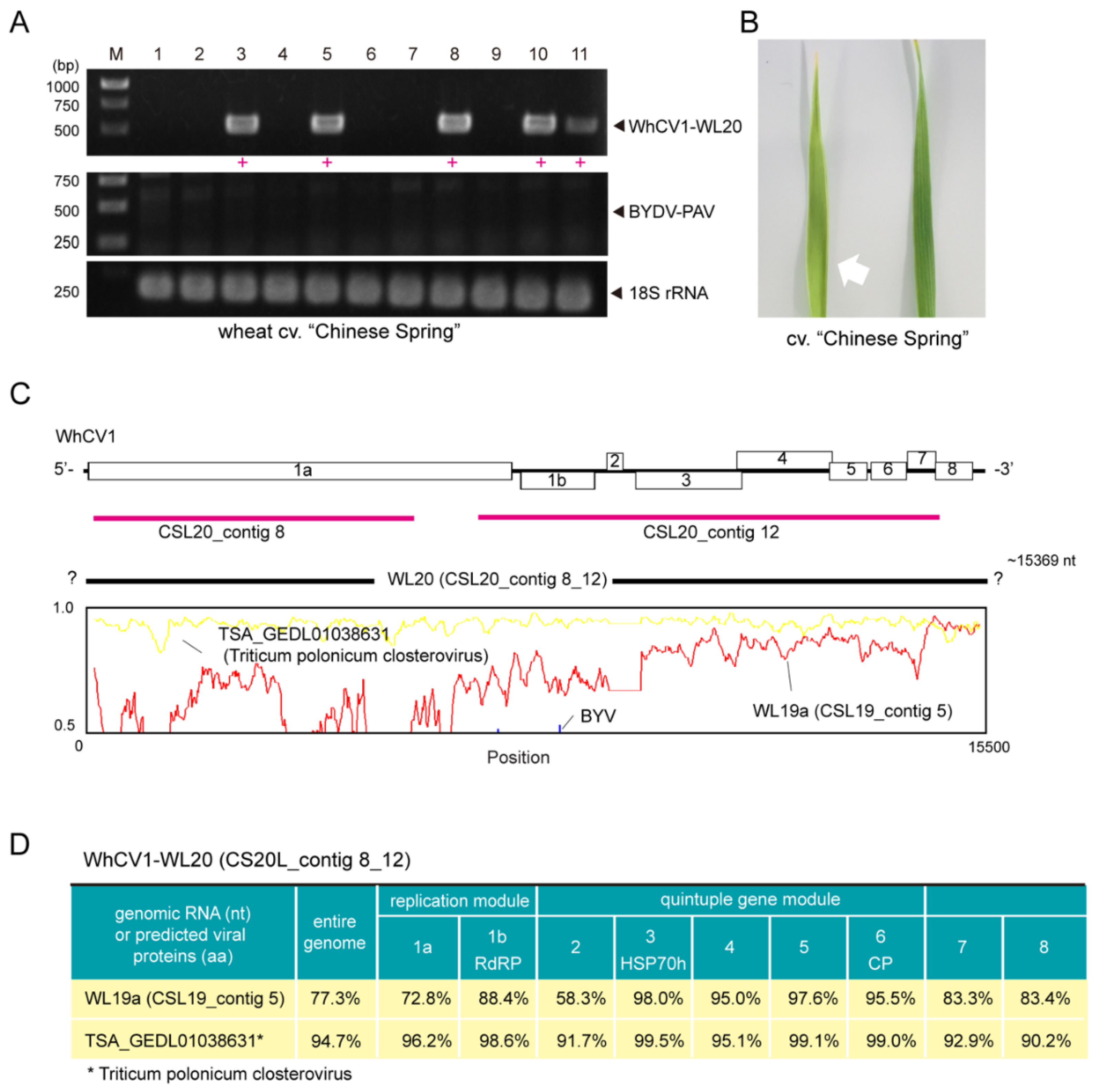
2.4. Identification of a WhCV1 Variant and Evaluation of Its Aphid Transmission
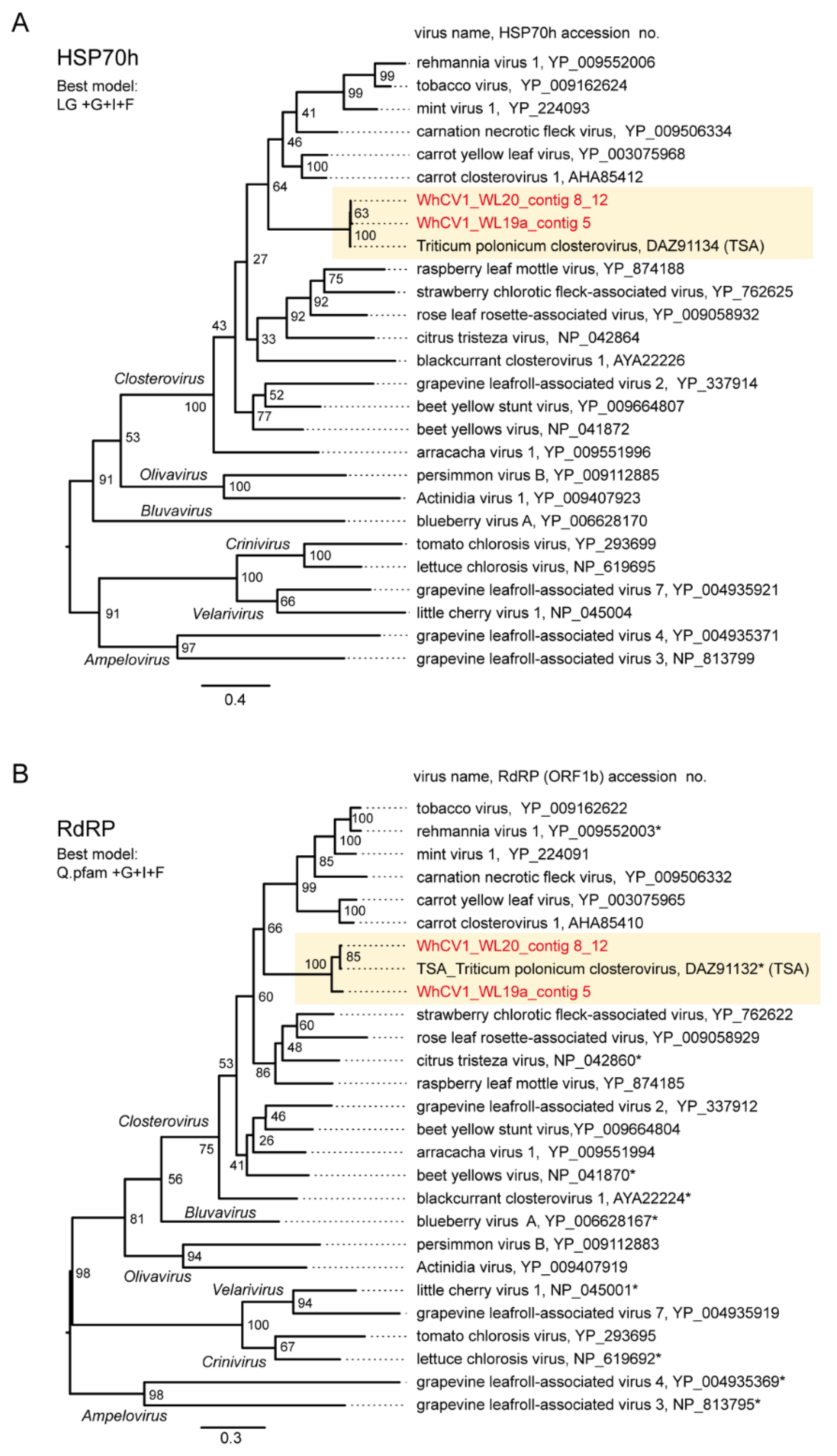
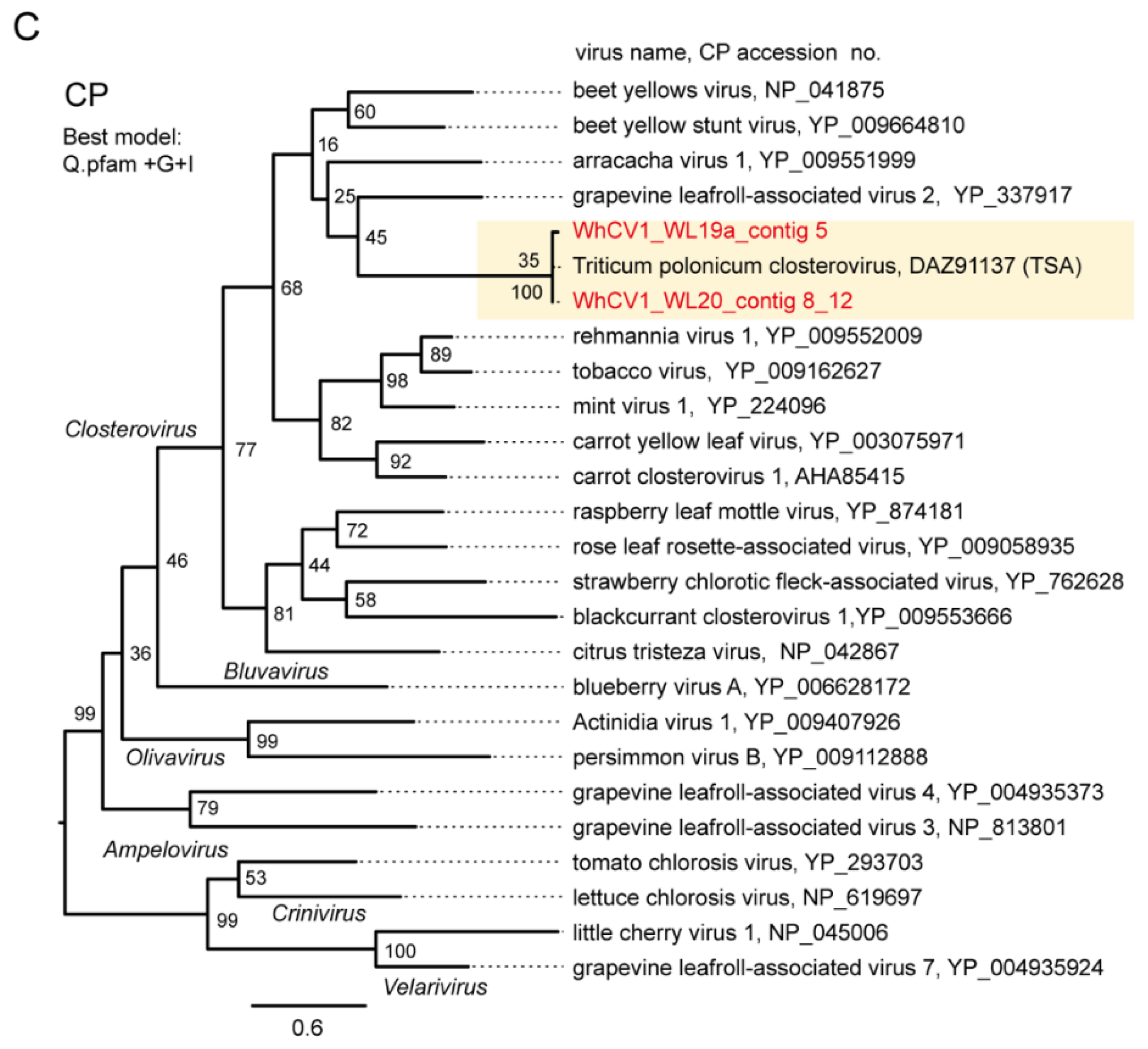
2.5. Similarity and Phylogenetic Relationship of WhCV1 and Other Closteroviruses
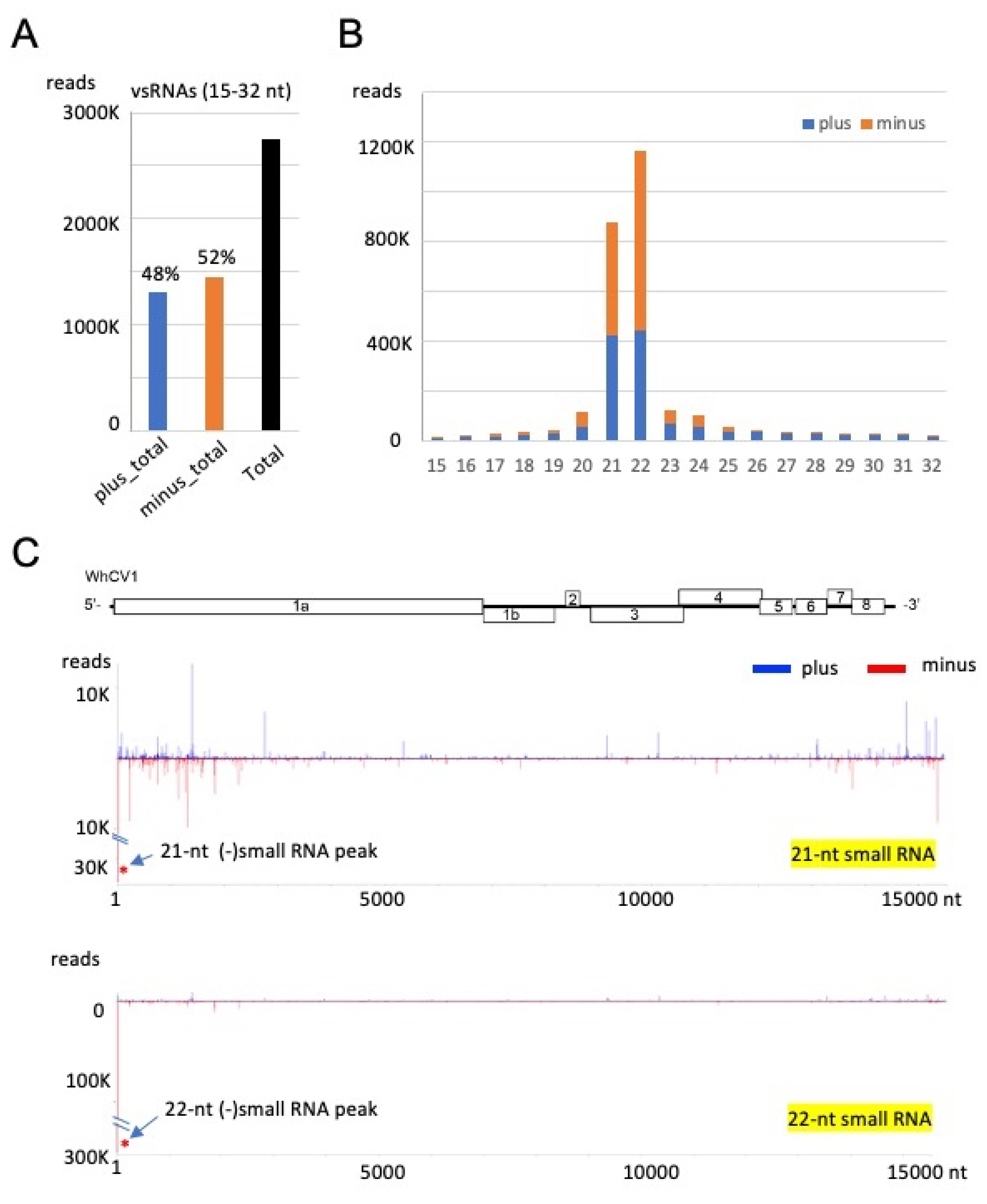
2.6. Small RNA Profiles of WhCV1
3. Discussion
4. Materials and Methods
4.1. Plant Sampling and RNA Extraction
4.2. Next-Generation Sequencing and De Novo Read Assembly
4.3. RT-PCR and RLM-RACE Analyses of Viral RNA Sequences
4.4. Database Search and Sequence Analysis
4.5. Phylogenetic Analysis
4.6. Small RNA Analysis
4.7. Aphid Transmission Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, C.C.; Kan, J.H.; Ordon, F.; Perovic, D.; Yang, P. Bymovirus-induced yellow mosaic diseases in barley and wheat: Viruses, genetic resistances and functional aspects. Theor. Appl. Genet. 2020, 133, 1623–1640. [Google Scholar] [CrossRef]
- Lapierre, H.D.; Hariri, D. Cereal viruses: Wheat and barley. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., Regenmortel, M.H.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 490–497. [Google Scholar] [CrossRef]
- Jones, R.A.C. Global l plant virus disease pandemics and epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef]
- Miller, W.A.; Lozier, Z. Yellow dwarf viruses of cereals: Taxonomy and molecular mechanisms. Annu. Rev. Phytopathol. 2022, 60, 121–141. [Google Scholar] [CrossRef]
- Aradottir, G.I.; Crespo-Herrera, L. Host plant resistance in wheat to barley yellow dwarf viruses and their aphid vectors: A review. Curr. Opin. Insect Sci. 2021, 45, 59–68. [Google Scholar] [CrossRef]
- Svanella-Dumas, L.; Candresse, T.; Hulle, M.; Marais, A. Distribution of barley yellow dwarf virus-PAV in the sub-Antarctic Kerguelen islands and characterization of two new luteovirus species. PLoS ONE 2013, 8, e67231. [Google Scholar] [CrossRef] [Green Version]
- Minato, N.; Hatori, S.; Okawa, A.; Nakagawa, K.; Hironaka, M. Manipulation of insect vectors’ host selection behavior by barley yellow dwarf virus is dependent on the host plant species and viral co-infection. Life 2022, 12, 644. [Google Scholar] [CrossRef]
- Sano, Y.; Tanno, F.; Kato, Y.; Matsubara, A.; Kojima, M. Complete nucleotide sequence of the Japanese isolate of barley yellow dwarf virus-PAV serotype. Jpn. J. Phytopathol. 1996, 62, 566–571. [Google Scholar] [CrossRef]
- Fuji, S.; Mochizuki, T.; Okuda, M.; Tsuda, S.; Kagiwada, S.; Sekine, K.T.; Ugaki, M.; Natsuaki, K.T.; Isogai, M.; Maoka, T.; et al. Plant viruses and viroids in Japan. J. Gen. Plant Pathol. 2022, 88, 105–127. [Google Scholar] [CrossRef]
- Inouye, T.; Mitsuhata, K.; Heta, H.; Hiura, U. A Novel Virus, Wheat Yellow Leaf Virus, Occuring Wheat, Barley and Several other Plants in Gramineae. Bull. Res. Inst. Bioresour. Okayama Univ. 1973, 55, 1–6. Available online: https://ousar.lib.okayama-u.ac.jp/51023 (accessed on 15 February 2023).
- Inouye, T. Wheat Yellow Leaf Virus. CMI/AAB Descr. Plant Viruses 1976. Available online: https://www.dpvweb.net/dpv/showdpv/?dpvno=157 (accessed on 15 February 2023).
- Fuchs, M.; Bar-Joseph, M.; Candresse, T.; Maree, H.J.; Martelli, G.P.; Melzer, M.J.; Menzel, W.; Minafra, A.; Sabanadzovic, S.; Consortium, I.R. ICTV virus taxonomy profile: Closteroviridae. J. Gen. Virol. 2020, 101, 364–365. [Google Scholar] [CrossRef]
- Dolja, V.V.; Kreuze, J.F.; Valkonen, J.P. Comparative and functional genomics of closteroviruses. Virus Res. 2006, 117, 38–51. [Google Scholar] [CrossRef]
- Shin, C.; Choi, D.; Shirasu, K.; Ichihashi, Y.; Hahn, Y. A novel RNA virus, Thesium chinense closterovirus 1, identified by high-throughput RNA-sequencing of the parasitic plant Thesium chinense. Acta Virol. 2022, 66, 206–215. [Google Scholar] [CrossRef]
- Wang, D.F.; Cui, L.Y.; Yu, P.X.; Qi, Y.H.; Wang, B.X.; Niu, Y.B. A new closterovirus in Alcea rosea identified by small RNA deep sequencing. J. Plant Pathol. 2020, 102, 29–37. [Google Scholar] [CrossRef]
- Park, D.; Goh, C.J.; Hahn, Y. Two novel closteroviruses, fig virus A and fig virus B, identified by the analysis of the high-throughput RNA-sequencing data of fig (Ficus carica) latex. Acta Virol. 2021, 65, 42–48. [Google Scholar] [CrossRef]
- Orilio, A.F.; Blawid, R.; Costa, G.A.; Gomes, S.; Nagata, T.; Madeira, N.R.; Inoue-Nagata, A.K.; Resende, R.O. High-throughput sequencing reveals a novel closterovirus in arracacha (Arracacia xanthorrhiza). Arch. Virol. 2018, 163, 2547–2550. [Google Scholar] [CrossRef]
- He, Y.; Yang, Z.K.; Hong, N.; Wang, G.P.; Ning, G.G.; Xu, W.X. Deep sequencing reveals a novel closterovirus associated with wild rose leaf rosette disease. Mol. Plant Pathol. 2015, 16, 449–458. [Google Scholar] [CrossRef]
- Kwon, S.J.; Jin, M.; Cho, I.S.; Yoon, J.Y.; Choi, G.S. Identification of rehmannia virus 1, a novel putative member of the genus Closterovirus, from Rehmannia glutinosa. Arch. Virol. 2018, 163, 3383–3388. [Google Scholar] [CrossRef]
- Zheng, L.P.; Wu, L.P.; Postman, J.; Liu, H.W.; Li, R.H. Molecular characterization and detection of a new closterovirus identified from blackcurrant by high-throughput sequencing. Virus Genes 2018, 54, 828–832. [Google Scholar] [CrossRef]
- Gudeta, W.F.; Belete, M.T.; Igori, D.; Kim, S.E.; Moon, J.S. Complete genome sequence of cnidium closterovirus 1, a novel member of the genus Closterovirus infecting Cnidium officinale. Arch. Virol. 2022, 167, 1491–1494. [Google Scholar] [CrossRef]
- Igori, D.; Lim, S.; Cho, H.S.; Kim, H.S.; Park, J.M.; Lee, H.J.; Kwon, S.Y.; Moon, J.S. Complete genome sequence of platycodon closterovirus 1, a novel putative member of the genus Closterovirus. Arch. Virol. 2021, 166, 2051–2054. [Google Scholar] [CrossRef]
- Koloniuk, I.; Thekke-Veetil, T.; Reynard, J.S.; Plesko, I.M.; Pribylova, J.; Brodard, J.; Kellenberger, I.; Sarkisova, T.; Spak, J.; Lamovsek, J.; et al. Molecular characterization of divergent closterovirus isolates infecting Ribes species. Viruses 2018, 10, 369. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.Y.; Zhang, S.; Mei, S.Q.; Zhou, Y.; Wang, J.H.; Han, G.Z.; Chen, L.; Zhou, C.Y.; Cao, M.J. Viromics unveils extraordinary genetic diversity of the family Closteroviridae in wild citrus. PLoS Pathog. 2021, 17, e1009751. [Google Scholar] [CrossRef]
- Satyanarayana, T.; Gowda, S.; Ayllon, M.A.; Dawson, W.O. Closterovirus bipolar virion: Evidence for initiation of assembly by minor coat protein and its restriction to the genomic RNA 5′ region. Proc. Natl. Acad. Sci. USA 2004, 101, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Gowda, S.; Satyanarayana, T.; Ayllon, M.A.; Moreno, P.; Flores, R.; Dawson, W.O. The conserved structures of the 5’ nontranslated region of Citrus tristeza virus are involved in replication and virion assembly. Virology 2003, 317, 50–64. [Google Scholar] [CrossRef] [Green Version]
- Satyanarayana, T.; Gowda, S.; Ayllon, M.A.; Albiach-Marti, M.R.; Dawson, W.O. Mutational analysis of the replication signals in the 3’-nontranslated region of Citrus tristeza virus. Virology 2002, 300, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Morozov, S.Y.; Agranovsky, A.A. Alphaflexiviruses (Alphaflexiviridae). In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2021; pp. 140–148. [Google Scholar]
- Dolja, V.V.; Koonin, E.V. The closterovirus-derived gene expression and RNA interference vectors as tools for research and plant biotechnology. Front. Microbiol. 2013, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Firth, A.E.; Brierley, I. Non-canonical translation in RNA viruses. J. Gen. Virol. 2012, 93, 1385–1409. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V. Evolution and taxonomy of positive-strand RNA viruses—Implications of comparative-analysis of amino-acid-sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef]
- Kondo, H.; Chiba, S.; Maruyama, K.; Andika, I.B.; Suzuki, N. A novel insect-infecting virga/nege-like virus group and its pervasive endogenization into insect genomes. Virus Res. 2019, 262, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, C.; Wang, X.L.; Peng, F.; Wang, R.J.; Jiang, Y.L.; Zeng, J.; Fan, X.; Kang, H.Y.; Sha, L.N.; et al. De novo sequencing and characterization of the transcriptome of dwarf Polish wheat (Triticum polonicum L.). Int. J. Genom. 2016, 2016, 5781412. [Google Scholar] [CrossRef] [Green Version]
- Sidharthan, V.K.; Baranwal, V.K. Dwarf polish wheat hosts a novel closterovirus: Revelation by transcriptome data-mining. Acta Virol. 2022, 66, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Yoshida, N.; Fujita, M.; Maruyama, K.; Hyodo, K.; Hisano, H.; Tamada, T.; Andika, I.B.; Suzuki, N. Identification of a novel quinvirus in the family Betaflexiviridae that infects winter wheat. Front. Microbiol. 2021, 12, 715545. [Google Scholar] [CrossRef]
- Poretti, M.; Praz, C.R.; Meile, L.; Kälin, C.; Schaefer, L.K.; Schläfli, M.; Widrig, W.; Sanchez-Vallet, A.; Wicker, T.; Bourras, S. Domestication of high-copy transposons underlays the wheat small RNA response to an obligate pathogen. Mol. Biol. Evol. 2020, 37, 839–848. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Garnsey, S.M.; Gonsalves, D. The closteroviruses: A distinct group of elongated plant viruses. Adv. Virus Res. 1979, 25, 93–168. [Google Scholar] [CrossRef]
- Usugi, T.; Nakano, M.; Shinkai, A. Purification and serological diagnosis of barley yellow dwarf virus. Proc. Assoc. Pl. Prot. Kyushu 1988, 34, 10–12. [Google Scholar] [CrossRef]
- Dupont, P.Y.; Eaton, C.J.; Wargent, J.J.; Fechtner, S.; Solomon, P.; Schmid, J.; Day, R.C.; Scott, B.; Cox, M.P. Fungal endophyte infection of ryegrass reprograms host metabolism and alters development. New Phytol. 2015, 208, 1227–1240. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.W.; Peremyslov, V.V.; Mushegian, A.R.; Dawson, W.O.; Dolja, V.V. Functional specialization and evolution of leader proteinases in the family Closteroviridae. J. Virol. 2001, 75, 12153–12160. [Google Scholar] [CrossRef] [Green Version]
- Gushchin, V.A.; Karlin, D.G.; Makhotenko, A.V.; Khromov, A.V.; Erokhina, T.N.; Solovyev, A.G.; Morozov, S.Y.; Agranovsky, A.A. A conserved region in the Closterovirus la polyprotein drives extensive remodeling of endoplasmic reticulum membranes and induces motile globules in Nicotiana bentharniana cells. Virology 2017, 502, 106–113. [Google Scholar] [CrossRef]
- Bartholomaus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Identification of a novel mycovirus isolated from Rhizoctonia solani (AG 2-2 IV) provides further information about genome plasticity within the order Tymovirales. Arch. Virol. 2017, 162, 555–559. [Google Scholar] [CrossRef]
- Alzhanova, D.V.; Napuli, A.J.; Creamer, R.; Dolja, V.V. Cell-to-cell movement and assembly of a plant closterovirus: Roles for the capsid proteins and Hsp70 homolog. Embo J. 2001, 20, 6997–7007. [Google Scholar] [CrossRef] [Green Version]
- Prokhnevsky, A.I.; Peremyslov, V.V.; Napuli, A.J.; Dolja, V.V. Interaction between long-distance transport factor and Hsp70-related movement protein of beet yellows virus. J. Virol. 2002, 76, 11003–11011. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.C.; Kasschau, K.D.; Prokhnevsky, A.I.; Gopinath, K.; Pogue, G.P.; Carrington, J.C.; Dolja, V.V. Suppressor of RNA silencing encoded by Beet yellows virus. Virology 2003, 306, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Folimonov, A.; Shintaku, M.; Li, W.X.; Falk, B.W.; Dawson, W.O.; Ding, S.W. Three distinct suppressors of RNA silencing encoded by a 20-kb viral RNA genome. Proc. Natl. Acad. Sci. USA 2004, 101, 15742–15747. [Google Scholar] [CrossRef] [Green Version]
- Andika, I.B.; Kondo, H.; Sun, L.Y. Interplays between soil-borne plant viruses and RNA silencing-mediated antiviral defense in roots. Front. Microbiol. 2016, 7, 1458. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, A.F.; Barton, D.A.; Nakasugi, K.; Jackson, C.; Kalischuk, M.L.; Kawchuk, L.M.; Vaslin, M.F.S.; Correa, R.L.; Waterhouse, P.M. The luteovirus P4 movement protein is a suppressor of systemic RNA silencing. Viruses 2017, 9, 294. [Google Scholar] [CrossRef] [Green Version]
- Andika, I.B.; Maruyama, K.; Sun, L.; Kondo, H.; Tamada, T.; Suzuki, N. Differential contributions of plant Dicer-like proteins to antiviral defences against potato virus X in leaves and roots. Plant J. 2015, 81, 781–793. [Google Scholar] [CrossRef]
- Bouche, N.; Lauressergues, D.; Gasciolli, V.; Vaucheret, H. An antagonistic function for Arabidopsis DCL2 in development and a new function for DCL4 in generating viral siRNAs. Embo J. 2006, 25, 3347–3356. [Google Scholar] [CrossRef] [Green Version]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef]
- Golyaev, V.; Candresse, T.; Rabenstein, F.; Pooggin, M.M. Plant virome reconstruction and antiviral RNAi characterization by deep sequencing of small RNAs from dried leaves. Sci. Rep. 2019, 9, 19268. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ruiz, S.; Navarro, B.; Gisel, A.; Pena, L.; Navarro, L.; Moreno, P.; Di Serio, F.; Flores, R. Citrus tristeza virus infection induces the accumulation of viral small RNAs (21-24-nt) mapping preferentially at the 3’-terminal region of the genomic RNA and affects the host small RNA profile. Plant Mol. Biol. 2011, 75, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Licciardello, G.; Scuderi, G.; Ferraro, R.; Giampetruzzi, A.; Russo, M.; Lombardo, A.; Raspagliesi, D.; Bar-Joseph, M.; Catara, A. Deep sequencing and analysis of small RNAs in sweet orange grafted on sour orange infected with two citrus tristeza virus isolates prevalent in Sicily. Arch. Virol. 2015, 160, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Blevins, T.; Rajeswaran, R.; Aregger, M.; Borah, B.K.; Schepetilnikov, M.; Baerlocher, L.; Farinelli, L.; Meins, F.; Hohn, T.; Pooggin, M.M. Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense. Nucleic Acids Res. 2011, 39, 5003–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.A.; Shen, R.Z.; Staplin, W.; Kanodia, P. Noncoding RNAs of Plant Viruses and Viroids: Sponges of Host Translation and RNA Interference Machinery. Mol. Plant-Microbe Interact. 2016, 29, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollov, D.; Filloux, D.; Grinstead, S.; Bolus, S.; Claude, L.; Fernandez, E.; Julian, C.; Candresse, T.; Lockhart, B.; Roumagnac, P.; et al. Sugarcane mild mosaic virus: The rediscovery of an identified but unrecognized virus. Plant Pathol. 2022, 72, 312–321. [Google Scholar] [CrossRef]
- Peters, J.S.; Aguirre, B.A.; DiPaola, A.; Power, A.G. Ecology of yellow dwarf viruses in crops and grasslands: Interactions in the context of climate. Annu. Rev. Phytopathol. 2022, 60, 283–305. [Google Scholar] [CrossRef]
- Tsai, W.; Brosnan, C.A.; Mitter, N.; Dietzgen, R.G. Perspectives on plant virus diseases in a climate change scenario of elevated temperatures. Stress Biol. 2022, 2, 37. [Google Scholar] [CrossRef]
- Lin, Y.H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef]
- Jarosova, J.; Kundu, J.K. Validation of reference genes as internal control for studying viral infections in cereals by quantitative real-time RT-PCR. BMC Plant Biol. 2010, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Chiba, S.; Tani, A.; Kondo, H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel quadripartite dsRNA virus isolated from a phytopathogenic filamentous fungus, Rosellinia necatrix. Virology 2012, 426, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.N.; Wang, J.Y.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Chiba, S.; Suzuki, N. Detection and analysis of non-retroviral RNA virus-like elements in plant, fungal, and insect genomes. In Plant Virology Protocols—New Approaches to Detect Viruses and Host Responses, 3rd ed.; Uyeda, I., Masuta, C., Eds.; Springer Protocol; Humana Press Inc.: New York, NY, USA, 2015; pp. 73–88. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, R.; Serra, F.; Tarraga, J.; Medina, I.; Carbonell, J.; Pulido, L.; de Maria, A.; Capella-Gutierrez, S.; Huerta-Cepas, J.; Gabaldon, T.; et al. Phylemon 2.0: A suite of web-tools for molecular evolution, phylogenetics, phylogenomics and hypotheses testing. Nucleic Acids Res. 2011, 39, W470–W474. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Seguin, J.; Otten, P.; Baerlocher, L.; Farinelli, L.; Pooggin, M.M. MISIS-2: A bioinformatics tool for in-depth analysis of small RNAs and representation of consensus master genome in viral quasispecies. J. Virol. Methods 2016, 233, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Fujita, M.; Hisano, H.; Hyodo, K.; Andika, I.B.; Suzuki, N. Virome analysis of aphid populations that onfest the barley field: The discovery of two novel groups of nege/kita-like viruses and other novel RNA viruses. Front. Microbiol. 2020, 11, 509. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus-Isolate | Contig or Concatenate | Size (nt) | Total Reads | Accession No. |
|---|---|---|---|---|
| WhCV1-WL19a | CSL19_contig 5 | 15,452 (15,145) 1 | 1,995,569 | LC735716 |
| -WL20 | CSL20_contig 8_12 | 15,369 1 | 8,619,360 2 | LC735717 |
| BYDV-PAV-WL19b | CSL19_contig 1289 | 5699 | 37,336 | LC735718 |
| Top Hit Viruses | QC 1 | E-Value | Identity | Accession |
|---|---|---|---|---|
| ORF1a protein | ||||
| Rehmannia virus | 65% | 9e–160 | 34.4% | YP_009552004.1 |
| Tobacco virus | 64% | 2e–153 | 34.6% | YP_009162621.1 |
| Beet yellows virus | 47% | 2e–151 | 49.8% | AAF14300.1 |
| Cnidium closterovirus | 46% | 4e–150 | 45.6% | UNN55308.1 |
| Mint virus | 46% | 3e–148 | 44.6% | YP_224090.1 |
| ORF1b protein (RdRP) | ||||
| Soybean leaf crinkle mottle virus | 98% | 5e–175 | 61.3% | BCR37031.1 |
| Citrus tristeza virus | 95% | 6e–169 | 60.6% | ABW97532.1 |
| Tobacco virus | 97% | 9e–168 | 59.3% | YP_009162622.1 |
| Carnation yellow fleck virus | 98% | 1e–166 | 57.8% | BBK15488.1 |
| Alcea rosea virus | 97% | 2e–166 | 59.2% | QCW05658.1 |
| ORF3 protein (HSP70h) | ||||
| Cnidium closterovirus | 99% | 0.0 | 51.4% | UNN55311.1 |
| Carrot closterovirus | 99% | 0.0 | 51.3% | AHA85412.1 |
| Carrot yellow leaf virus | 99% | 0.0 | 50.5% | AHA85522.1 |
| Thesium chinense closterovirus | 99% | 0.0 | 47.2% | UQR78691.1 |
| Mint virus | 99% | 8e–177 | 46.0% | AAX98726.1 |
| ORF4 protein (HSP90h) | ||||
| Mint virus | 87% | 1e–140 | 43.9% | YP_224094.1 |
| Tobacco virus | 94% | 2e–133 | 41.7% | YP_009162625.1 |
| Rehmannia virus | 87% | 1e–128 | 41.9% | YP_009552007.1 |
| Alcea rosea virus | 87% | 4e–128 | 42.2% | QCW05661.1 |
| Beet yellows virus | 90% | 2e–126 | 38.4% | S28713 |
| ORF5 protein (CPm) | ||||
| Beet yellow stunt virus | 84% | 8e–38 | 38.0% | YP_009664809.1 |
| Carnation yellow fleck virus | 95% | 3e–37 | 33.3% | YP_008858535.1 |
| Beet yellows virus | 95% | 6e–35 | 31.3% | NP_041874.1 |
| Carrot yellow leaf virus | 95% | 2e–34 | 35.8% | AHA85464.1 |
| Mint virus | 91% | 1e–33 | 36.6% | YP_224095.1 |
| ORF6 protein (CP) | ||||
| Grapevine leafroll-associated virus 2 | 94% | 2e–16 | 33.5% | AHW79716.1 |
| Beet yellow stunt virus | 82% | 6e–13 | 32.4% | YP_009664810 |
| Carrot closterovirus | 91% | 9e–12 | 29.3% | AHA85415.1 |
| Carnation necrotic fleck virus | 93% | 1e–10 | 25.4% | BBK15484.1 |
| Carrot yellow leaf virus | 83% | 9e–08 | 29.1% | AHA85425.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, H.; Sugahara, H.; Fujita, M.; Hyodo, K.; Andika, I.B.; Hisano, H.; Suzuki, N. Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan. Pathogens 2023, 12, 358. https://doi.org/10.3390/pathogens12030358
Kondo H, Sugahara H, Fujita M, Hyodo K, Andika IB, Hisano H, Suzuki N. Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan. Pathogens. 2023; 12(3):358. https://doi.org/10.3390/pathogens12030358
Chicago/Turabian StyleKondo, Hideki, Hitomi Sugahara, Miki Fujita, Kiwamu Hyodo, Ida Bagus Andika, Hiroshi Hisano, and Nobuhiro Suzuki. 2023. "Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan" Pathogens 12, no. 3: 358. https://doi.org/10.3390/pathogens12030358
APA StyleKondo, H., Sugahara, H., Fujita, M., Hyodo, K., Andika, I. B., Hisano, H., & Suzuki, N. (2023). Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan. Pathogens, 12(3), 358. https://doi.org/10.3390/pathogens12030358