
Field Resistance to Rose Rosette Disease as Determined by Multi-Year Evaluations in Tennessee and Delaware

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Plot Design
2.3. RRD Augmentation Procedures
2.4. RRD Evaluation in the Field
2.5. Data Tabulation and Analysis
3. Results and Discussion
3.1. Disease Development over Years
3.2. Cultivar × Location Interaction
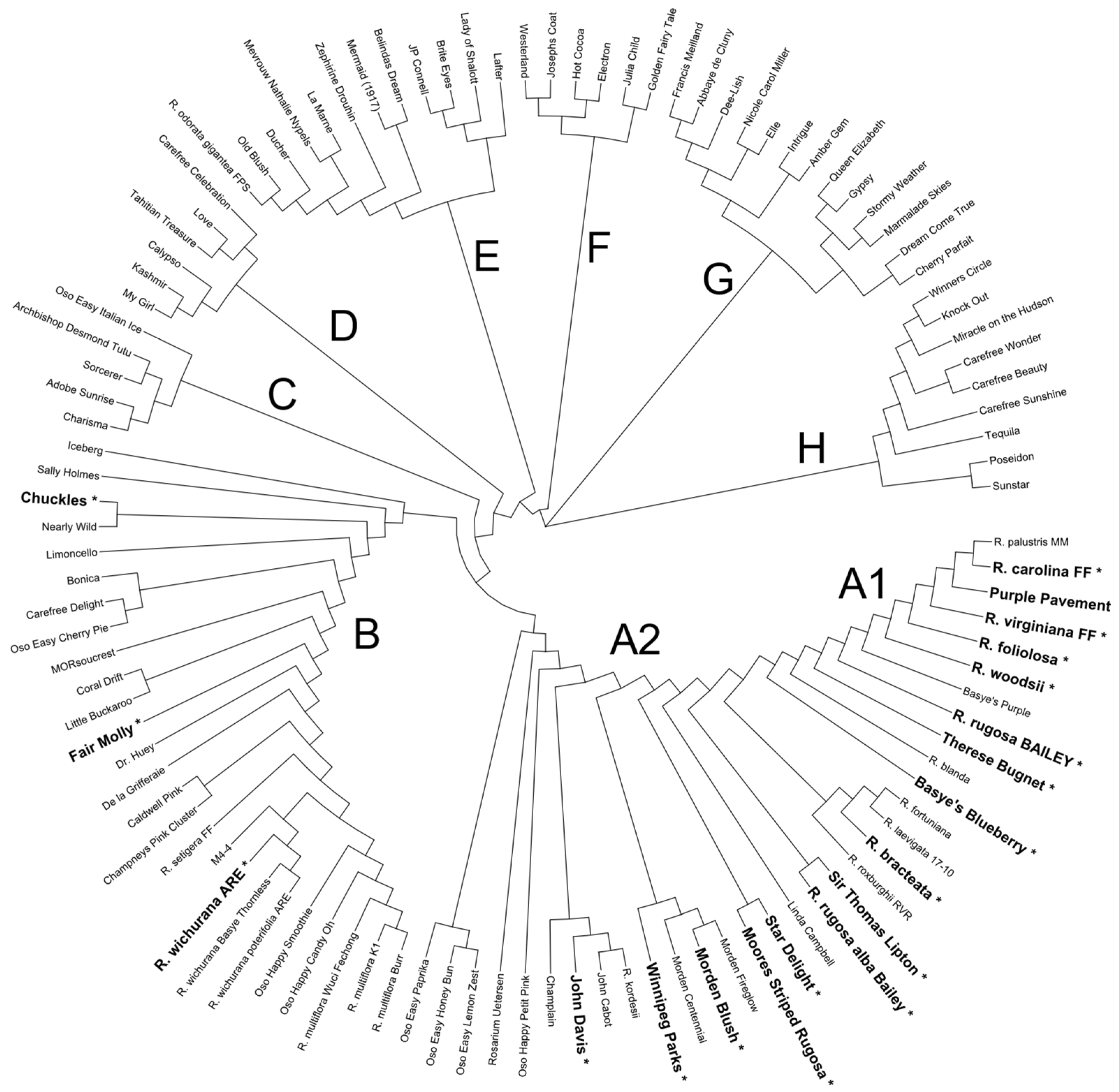
3.3. Genetic Relationships of the Roses
3.4. RRD Resistance among Roses
3.5. Mechanism of Resistance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pemberton, H.B.; Ong, K.; Windham, M.; Olsen, J.; Byrne, D.H. What is Rose Rosette Disease? Hort. Sci. 2018, 53, 592–595. [Google Scholar] [CrossRef] [Green Version]
- Debener, T.; Byrne, D.H. Disease resistance breeding in rose: Current status and potential of biotechnological tools. Plant Sci. 2014, 228, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.; Rebek, E.; Schnelle, M. Rose Rosette Disease. Oklahoma Cooperative Extension Service, EPP-7329. Available online: https://extension.okstate.edu/fact-sheets/print-publications/epp-entomology-and-plant-pathologhy/rose-rosette-disease-epp-7329.pdf (accessed on 1 March 2023).
- Ong, K.; Giesbrecht, M.; Woodson, D.; Miller, L. Rose Rosette Demystified. Texas A&M AgriLife Ext. Bul. 2015. EPLP-010. Available online: https://agrilifelearn.tamu.edu/s/product/rose-rosette-disease-demystified/01t4x000004OfrrAAC (accessed on 1 March 2023).
- Windham, M.T.; Hale, F.A.; Windham, A.S. Managing Rose Rosette in the Landscape—Ideas based on experimental data. Rose Mag. 2016, 44, 36–38. [Google Scholar]
- Di Bello, P.L.; Thekke-Veetil, T.; Druciarek, T.; Tzanetakis, I.E. Transmission attributes and resistance to rose rosette virus. Plant Pathol. 2018, 67, 499–504. [Google Scholar] [CrossRef]
- Shires, M.K. Study of Resistance to Rose Rosette Disease Utilizing Field Research, Molecular Tools, and Transmission Method. Ph.D. Dissertation, Texas A&M University, College Station, TX, USA, December 2020. [Google Scholar]
- Mielke-Ehret, N.; Mühlbach, H.P. Emaravirus: A novel genus of multipartite, negative strand RNA plant viruses. Viruses 2012, 4, 1515–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laney, A.G.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A discovery 70 years in the making: Characterization of the Rose rosette virus. J. Gen. Virol. 2011, 92, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Di Bello, P.L.; Ho, T.; Tzanetakis, I.E. Evolution of emaraviruses is becoming more complex: Seven segments identified in the causal agent of Rose rosette disease. Virus Res. 2015, 210, 241–244. [Google Scholar] [CrossRef]
- Babu, B.; Jeyaprakash, A.; Jones, D.; Schubert, T.S.; Baker, C.; Washburn, B.K.; Miller, S.H.; Poduch, K.; Knox, G.W.; Ochoa-Corona, F.M.; et al. Development of a rapid, sensitive TaqMan real-time RT-PCR assay for the detection of Rose rosette virus using multiple gene targets. J. Virol. Methods 2016, 235, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Katsiani, A.; Stainton, D.; Lamour, K.; Tzanetakis, I.E. The population structure of Rose rosette virus in the USA. J. Gen. Virol. 2020, 101, 676–684. [Google Scholar] [CrossRef]
- Amrine, J.W. What happens to Phyllocoptes fructiphilus the vector of Rose rosette Virus in winter? Am. Rose 2014, 42, 118–121. [Google Scholar]
- Bauchan, G.; Otero-Colina, G.; Hammond, J.; Jordan, R.; Ochoa, R. Rose rosette disease: It all started with a small mite. Acta Hortic. 2019, 1232, 227–232. [Google Scholar] [CrossRef]
- Monterrosa, A.; Paret, M.L.; Ochoa, R.; Ulsamer, A.; Joseph, S.V. Temporal Incidence of Eriophyid Mites on Rose Rosette Disease-Symptomatic and -Asymptomatic Roses in Central Georgia, USA. Pathogens 2022, 11, 228. [Google Scholar] [CrossRef] [PubMed]
- Allington, W.B.; Staples, R.; Viehmeyer, G. Transmission of Rose Rosette Virus by the Eriophyid Mite Phyllocoptes fructiphilus1. J. Econ. Èntomol. 1968, 61, 1137–1140. [Google Scholar] [CrossRef]
- Amrine, J.W.; Hindal, D.F.; Stasny, T.A.; Williams, R.L.; Coffman, C.C. Transmission of the rose rosette disease agent to Rosa multiflora by Phyllocoptes fructiphilus (Acari: Eriophyidae). Entomol. News 1988, 99, 239–252. [Google Scholar]
- Amrine, J.W. Phyllocoptes fructiphilus and biological control of multiflora rose. In Eriophyid Mites—Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruins, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; pp. 741–749. [Google Scholar]
- Amrine, J.W. Multiflora rose. In Biological control of Invasive Plants in the Eastern United States; Van Driesche, R., Blossey, B., Huddle, M., Lyon, S., Reardon, R., Eds.; USDA: Washington, DC, USA, 2002; pp. 272–309. [Google Scholar]
- Solo, K.M.; Collins, S.B.; Shires, M.K.; Ochoa, R.; Bauchan, G.R.; Schneider, L.; Henn, A.; Jacobi, J.C.; Williams-Woodward, J.L.; Hajimorad, M.; et al. A Survey of Rose rosette virus and Eriophyid Mites Associated with Roses in the Southeastern United States. Hortscience 2020, 55, 1288–1294. [Google Scholar] [CrossRef]
- Tipping, P.W.; Sindermann, A.B. Natural and Augmented Spread of Rose Rosette Disease of Multiflora Rose in Maryland. Plant Dis. 2000, 84, 1344. [Google Scholar] [CrossRef]
- Babu, B.; Dankers, H.; Newberry, E.; Baker, C.; Schubert, T.; Knox, G.; Paret, M. First Report of Rose rosette virus Associated with Rose Rosette Disease Infecting Knockout Roses in Florida. Plant Dis. 2014, 98, 1449. [Google Scholar] [CrossRef]
- Singh, R.; Valverde, R.; Cook, M.; Owings, A. First Report of Rose Rosette Disease Caused by Rose rosette virus on Knock Out Roses in Louisiana. Plant Health Prog. 2016, 17, 195–196. [Google Scholar] [CrossRef] [Green Version]
- Bratsch, S.; Zlesak, D.; Mollov, D.; Lockhart, B. First report of Rose Rosette virus associated with rose rosette disease in Rosa hybrid in Minnesota. Plant Health Prog. 2017, 18, 102–103. [Google Scholar] [CrossRef] [Green Version]
- Al Rwahnih, M.; Karlik, J.; Diaz-Lara, A.; Ong, K.; Mollov, D.; Haviland, D.; Golino, D. First report of Rose rosette virus associated with rose rosette disease affecting rose in California. Plant Dis. 2019, 103, 380. [Google Scholar] [CrossRef]
- Vazquez-Iglesias, I.; Ochoa-Corona, F.M.; Tang, J.; Robinson, R.; Clover, G.R.; Fox, A.; Boonham, N. Facing Rose rosette virus: A risk to European rose cultivation. Plant Pathol. 2020, 69, 1603–1617. [Google Scholar] [CrossRef]
- Amrine, J.W.; Kassar, A.; Stasny, T.A. Phyllocoptes fructiphilus K. (Acari: Eriophyoi- dea), the vector of Rose Rosette disease, taxonomy, biology and distribution. In Proceedings of the Rose Rosette and Other Eriophyid Mite- transmitted Plant Disease Agents of Uncertain Etiology, Iowa State University, Ames, IA, USA, 19–21 May 1994; pp. 61–66. [Google Scholar]
- Young, E.L.; Lau, J.; Bentley, N.B.; Rawadoozi, Z.; Collins, S.; Windham, M.T.; Klein, P.E.; Byrne, D.H.; Riera-Lizarazu, O. Identification of QTLs for reduced susceptibility to rose rosette disease in diploid roses. Pathogens 2022, 11, 660. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.; Young, E.L.; Collins, S.; Windham, M.T.; Klein, P.E.; Byrne, D.H.; Riera-Lizarazu, O. Rose rosette disease resistance loci detected in two interconnected tetraploid garden rose populations. Front. Plant Sci. 2022, 13, 916231. [Google Scholar] [CrossRef] [PubMed]
- Solo, K.M.; Collins, S.B.; Schneider, L.; Hajimorad, M.R.; Hale, F.A.; Wilkerson, J.B.; Windham, A.S.; Byrne, D.H.; Windham, M.T. Evaluation of Rosa Species Accessions for Resistance to Eriophyid Mites1. J. Environ. Hortic. 2019, 37, 108–112. [Google Scholar] [CrossRef]
- Hall, C.R.; Dickson, M.W. Economic, Environmental, and Health/Well-Being Benefits Associated with Green Industry Products and Services: A Review. J. Environ. Hortic. 2011, 29, 96–103. [Google Scholar] [CrossRef]
- Chavez, D.E.; Palma, M.A.; Byrne, D.H.; Hall, C.R.; Ribera, L.A. Willingness to Pay for Rose Attributes: Helping Provide Consumer Orientation to Breeding Programs. J. Agric. Appl. Econ. 2019, 52, 1–15. [Google Scholar] [CrossRef] [Green Version]
- USDA. Census of Agriculture. Census of Horticultural Specialities. 2015, 3, Special Studies, Part 3. AC-12-SS-3. Available online: https://omb.report/icr/201906-0535-002/doc/92748901 (accessed on 1 March 2023).
- USDA. Census of Agriculture. Census of Horticultural Specialities. 2017, 2020, 3, Part 3. AC-17-SS-3. Available online: https://www.nass.usda.gov/Publications/AgCensus/2017/Online_Resources/Census_of_Horticulture_Specialties/HORTIC.pdf (accessed on 1 March 2023).
- Byrne, D.H.; Klein, P.; Yan, M.; Young, E.; Lau, J.; Ong, K.; Shires, M.; Olson, J.; Windham, M.; Evans, T.; et al. Challenges of Breeding Rose Rosette–resistant Roses. Hortscience 2018, 53, 604–608. [Google Scholar] [CrossRef]
- Waliczek, T.M.; Byrne, D.; Holeman, D. Opinions of Landscape Roses Available for Purchase and Preferences for the Future Market. Horttechnology 2018, 28, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Prevosti, A.; Alonso, G. Distances between populations of Drosophila subobscura, based on chromosome arrangement frequencies. Theor. Appl. Genet. 1975, 45, 231–241. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2014, e281. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Dobhal, S.; Olson, J.D.; Arif, M.; Suarez, J.A.G.; Ochoa-Corona, F.M. A simplified strategy for sensitive detection of Rose rosette virus compatible with three RT-PCR chemistries. J. Virol. Methods 2016, 232, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, B.; Washburn, B.K.; Ertek, T.S.; Miller, S.H.; Riddle, C.B.; Knox, G.W.; Ochoa-Corona, F.M.; Olson, J.; Katircioglu, Y.Z.; Paret, M.L. A field based detection method for Rose rosette virus using isothermal probe based Reverse transcription-recombinase polymerase amplification assay. J. Virol. Methods 2017, 247, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Zlesak, D. Rose. Rosa x hybrida. In Flower Breeding and Genetics. Issues, Challenges, and Opportunities for the 21st Century; Anderson, N.O., Ed.; Springer: New York, NY, USA, 2006; pp. 695–738. [Google Scholar]
- Shepherd, R.E. History of the Rose; MacMillan Publishing Co.: New York, NY, USA, 1954; p. 203. [Google Scholar]
- Harness, J. Roses; J. M. Dent & Sons Ltd.: London, UK, 1978; p. 290. [Google Scholar]
- Wisseman, V.; Ritz, C.M. The genus Rosa (Rosoideae, Rosaceae) revisited: Molecular analysis of nrITS-1 and atpB-rbcL intergenic spacer (IGS) versus conventional taxonomy. Bot. J. Linn. Soc. 2005, 147, 275–290. [Google Scholar] [CrossRef]
- Byrne, D.; Anderson, N. Legacies of Robert Basye and Ralph Moore: Historical perspective. Acta Hortic. 2019, 171–176. [Google Scholar] [CrossRef]


{kind=link}
| Climber Brite Eyes, RADbrite Winner’s Circle™, RADwin Rosarium Uetersen, KORtersen Stormy Weather, ORAfantanov Westerland, KORwest Floribunda Adobe Sunrise, MEIpluvia Bonica, MEIdomonac Charisma, JELroganor Chuckles (1957) Eyeconic Melon Lemonade, SPRomel Hot Cocoa, WEKpaltlez Iceberg, KORbin Intrigue, JACum Joseph’s Coat Julia Child, WEKvossutono Marmalade Skies, MEIlmonblan Nearly Wild Oso Easy Cherry Pie, MEIboulka Oso Easy Italian Ice, CHEwnicebell Poseidon, KORfriedhar Sevillana, MEIgekanu Sunstar, KORsteimm Tequila, MEIpomolo Cherry Parfait, MEIsponge Mevrouw Nathalie Nypels Hybrid Tea Abbaye de Cluny, MEIbrinpay Dee-lish, MEIclusif Electron Elle, MEIbderos Francis Meilland, MEItroni Golden Fairy Tale, KORquelda Gypsy (1972) Michelangelo, MEItelov Queen Elizabeth Grandiflora Dream Come True, WEKdocpot Love, JACtwin Nicole Carol Miller, MEIskimov Tahitian Treasure, RADtreasure | Miniature Little Buckaroo Oso Happy Petit Pink, Zlemariannayoshida Sorcerer, SAVasorc Fair Molly, MORfairpo Polyantha Caldwell Pink La Marne Oso Happy Smoothie, Zlecharlie Shrub Basye’s Blueberry Belinda’s Dream Carefree Beauty, BUCbi Carefree Celebration, RADral Carefree Delight, MEIpotal Carefree Sunshine, RADsun Carefree Wonder, MEIpitac Champlain Coral Drift, MEIdrifora Desmond Tutu, KORtutu Easy Elegance Calypso, BAIypso Easy Elegance Kashmir, BAImir Easy Elegance My Girl, BAIgirl J. P. Connell Knock Out ®, RADrazz Lafter Limoncello, MEIjecycka Miracle on the Hudson Morden Blush Morden Centennial Morden Fireglow Oso Easy Honey Bun, Scrivjean Oso Easy Lemon Zest, CHEwhocan Oso Easy Paprika, CHEmaytime Oso Happy Candy OH!, Zlemartincipr Red Drift, Meigalpio Winnipeg Parks | Bourbon Zéphirine Drouhin China Ducher Old Blush Noisette Champney’s Pink Cluster Species R. arkansana FF * R. bracteata RM R. carolina FF R. folialosa ARE R. odorata FPS R. roxburghii ARE R. rugosa alba Bailey R. rugosa Bailey R. soulieana-RM R. virginiana FF R. wichurana poterifolia ARE R. wichurana thornless ARE R. woodsii RVR R. x fortuniana Species hybrids Amber Gem Mermaid John Cabot John Davis Sally Holmes Basye’s Purple Fru Dagmar Hastrup Fuzzy Wuzzy Red Linda Campbell Moore’s Striped Rugosa Purple Pavement Sir Thomas Lipton Star Delight Therese Bugnet MORsoucrest Rootstocks De La Grifferaie Dr. Huey Manetti |
| Rosa arkansana FF | Chuckles |
| Rosa bracteata-RM | Fair Molly |
| Rosa carolina FF | Fuzzy Wuzzy Red |
| Rosa folialosa-ARE | John Davis |
| Rosa rugosa alba Bailey | Moore’s Striped Rugosa |
| Rosa rugosa Bailey | Morden Blush |
| Rosa virginiana FF | Purple Pavement |
| Rosa wichurana thornless ARE | Sir Thomas Lipton |
| Rosa woodsii RVR | Star Delight |
| Basye’s Blueberry | Therese Bugnet |
| Winnipeg Parks |
| Abbaye de Cluny | Elle | Mevrouw Nathalie Nypels |
| Adobe Sunrise | Eyeconic Melon Lemonade | Michelangelo |
| Amber Gem | Francis Meilland | Miracle on the Hudson |
| Basye’s Purple | Fru Dagmar Hastrup | Morden Centennial |
| Belinda’s Dream | Golden Fairy Tale | Morden Fireglow |
| Bonica | Gypsy | MORsoucrest |
| Brite Eyes | Hot Cocoa | Nicole Carol Miller |
| Caldwell Pink | Iceberg | Old Blush |
| Carefree Celebration | Intrigue | Oso Easy Cherry Pie |
| Carefree Delight | John Cabot | Oso Easy Lemon Zest |
| Carefree Sunshine | Joseph’s Coat | Oso Easy Paprika |
| Carefree Wonder | J.P. Connell | Oso Happy Petit Pink |
| Champlain | Julia Child | Oso Happy Smoothie |
| Champney’s Pink Cluster | Knock Out | Poseidon |
| Charisma | La Marne | Queen Elizabeth |
| Cherry Parfait | Lafter | Red Drift |
| De La Grifferaie | Limoncello | Rosa roxburghii ARE |
| Dr. Huey | Linda Campbell | Rosa wichurana poterfolia ARE |
| Dream Come True | Little Buckaroo | Rosarium Uetersen |
| Easy Elegance Kashmir | Love | Sorcerer |
| Easy Elegance My Girl | Manetti | Westerland |
| Electron | Marmalade Skies | Winner’s Circle |
| Mermaid |
| Carefree Beauty | Rosa odorata FPS |
| Coral Drift | Rosa soulieana-RM |
| Dee-lish | Rosa x fortuniana |
| Desmond Tutu | Sally Holmes |
| Ducher | Sevillana |
| Easy Elegance Calypso | Stormy Weather |
| Nearly Wild | Sunstar |
| Oso Easy Honey Bun | Tahitian Treasure |
| Oso Easy Italian Ice | Tequila |
| Oso Happy Candy OH! | Zephirine Drouhin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windham, M.T.; Evans, T.; Collins, S.; Lake, J.A.; Lau, J.; Riera-Lizarazu, O.; Byrne, D.H. Field Resistance to Rose Rosette Disease as Determined by Multi-Year Evaluations in Tennessee and Delaware. Pathogens 2023, 12, 439. https://doi.org/10.3390/pathogens12030439
Windham MT, Evans T, Collins S, Lake JA, Lau J, Riera-Lizarazu O, Byrne DH. Field Resistance to Rose Rosette Disease as Determined by Multi-Year Evaluations in Tennessee and Delaware. Pathogens. 2023; 12(3):439. https://doi.org/10.3390/pathogens12030439
Chicago/Turabian StyleWindham, Mark T., Thomas Evans, Sara Collins, Juniper A. Lake, Jeekin Lau, Oscar Riera-Lizarazu, and David H. Byrne. 2023. "Field Resistance to Rose Rosette Disease as Determined by Multi-Year Evaluations in Tennessee and Delaware" Pathogens 12, no. 3: 439. https://doi.org/10.3390/pathogens12030439
APA StyleWindham, M. T., Evans, T., Collins, S., Lake, J. A., Lau, J., Riera-Lizarazu, O., & Byrne, D. H. (2023). Field Resistance to Rose Rosette Disease as Determined by Multi-Year Evaluations in Tennessee and Delaware. Pathogens, 12(3), 439. https://doi.org/10.3390/pathogens12030439