
Spherical Body Protein 4 from Babesia bigemina: A Novel Gene That Contains Conserved B-Cell Epitopes and Induces Cross-Reactive Neutralizing Antibodies in Babesia ovata

 ,
,  , , ,
, , ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioethical Declarations
2.2. B. bigemina DNA Isolation
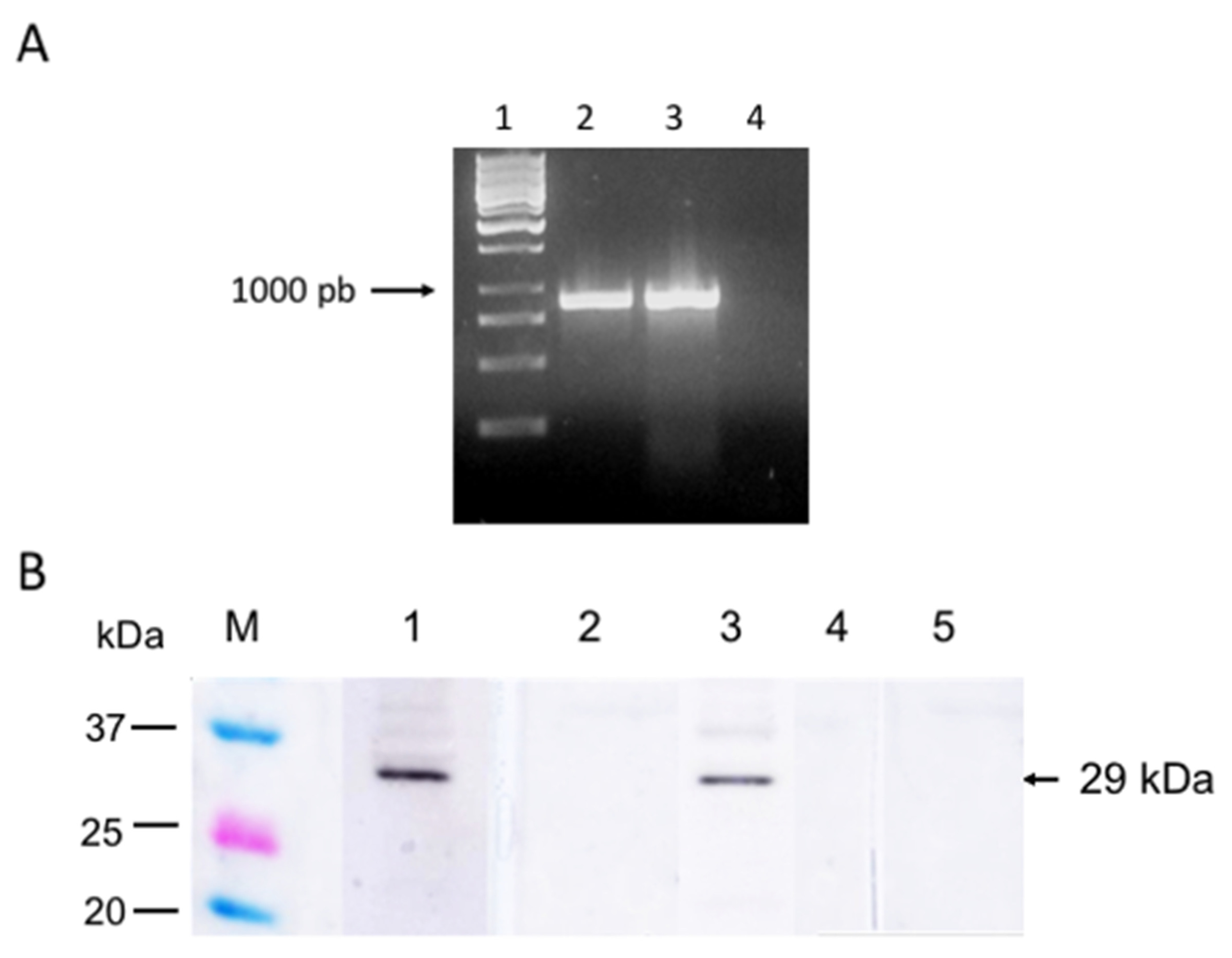
2.3. sbp4 Gene Amplification
2.4. Transcription Analysis
2.5. Cloning and Expression of sbp4 Gene
2.6. In Silico Analysis of Predicted SBP4 Protein Sequences
2.7. Search and Selection of Conserved B-Cell Epitopes in SBP4
2.8. SBP4 Peptide and Anti-rSBP4 Antisera Production
2.9. Expression Analysis of SBP4 in Merozoites
2.10. In Vitro Neutralization Assays with Antibodies against rSBP4
2.11. Recognition of SBP4 Peptides by Antibodies from Cattle Naturally Infected with B. bigemina
2.12. Neutralization Assays Using SBP4 Specific Peptides in B. bigemina Cultures In Vitro
3. Results
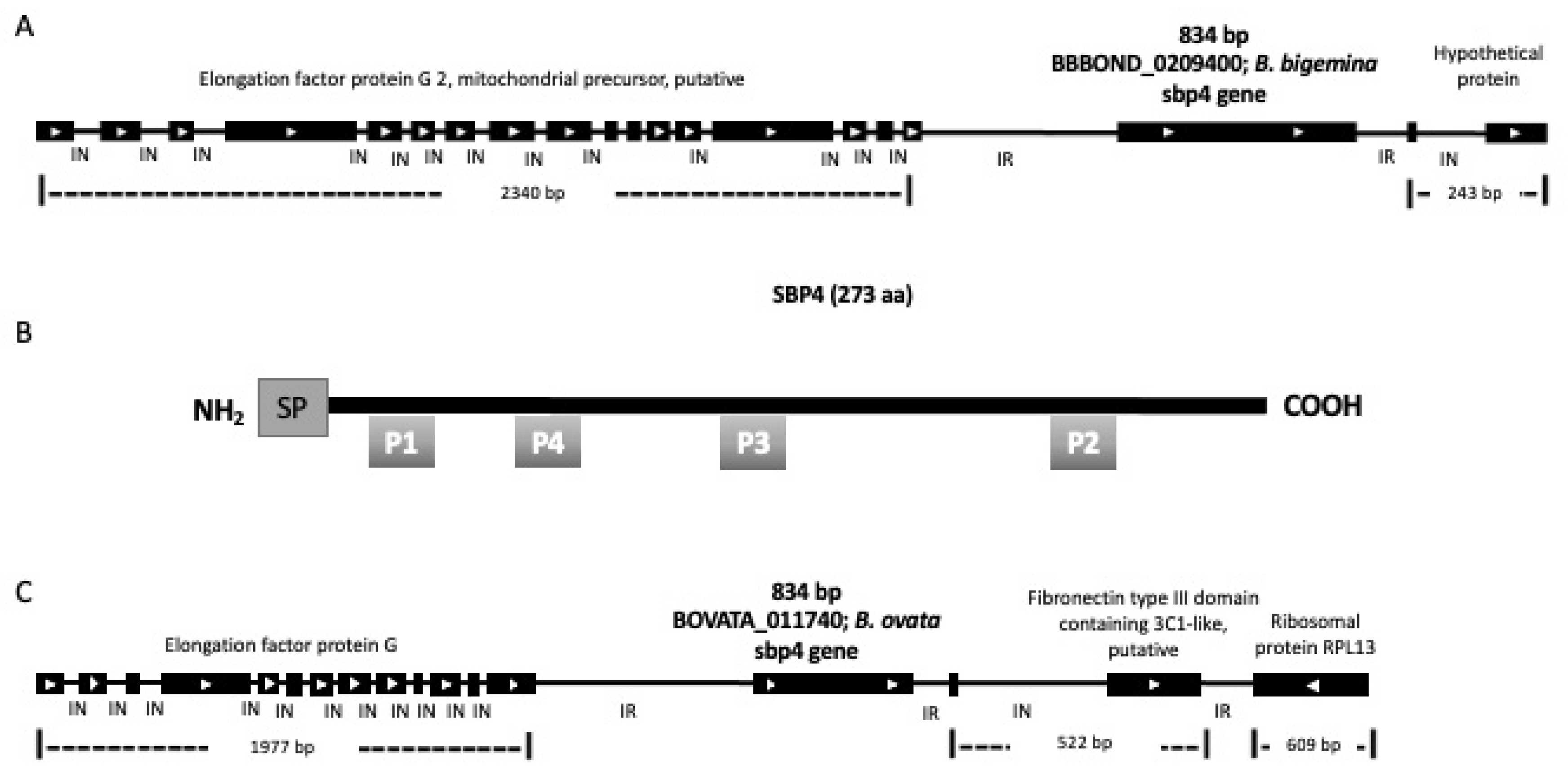
3.1. B. bigemina sbp4 Gene Identification and Characterization
3.2. SBP4 Is Transcribed and Expressed in B. bigemina Erythrocyte Stages
3.3. SBP4 Localizes to Round Organelles of B. bigemina and B. Ovata Merozoites and in the Cytoplasm of Infected Erythrocytes
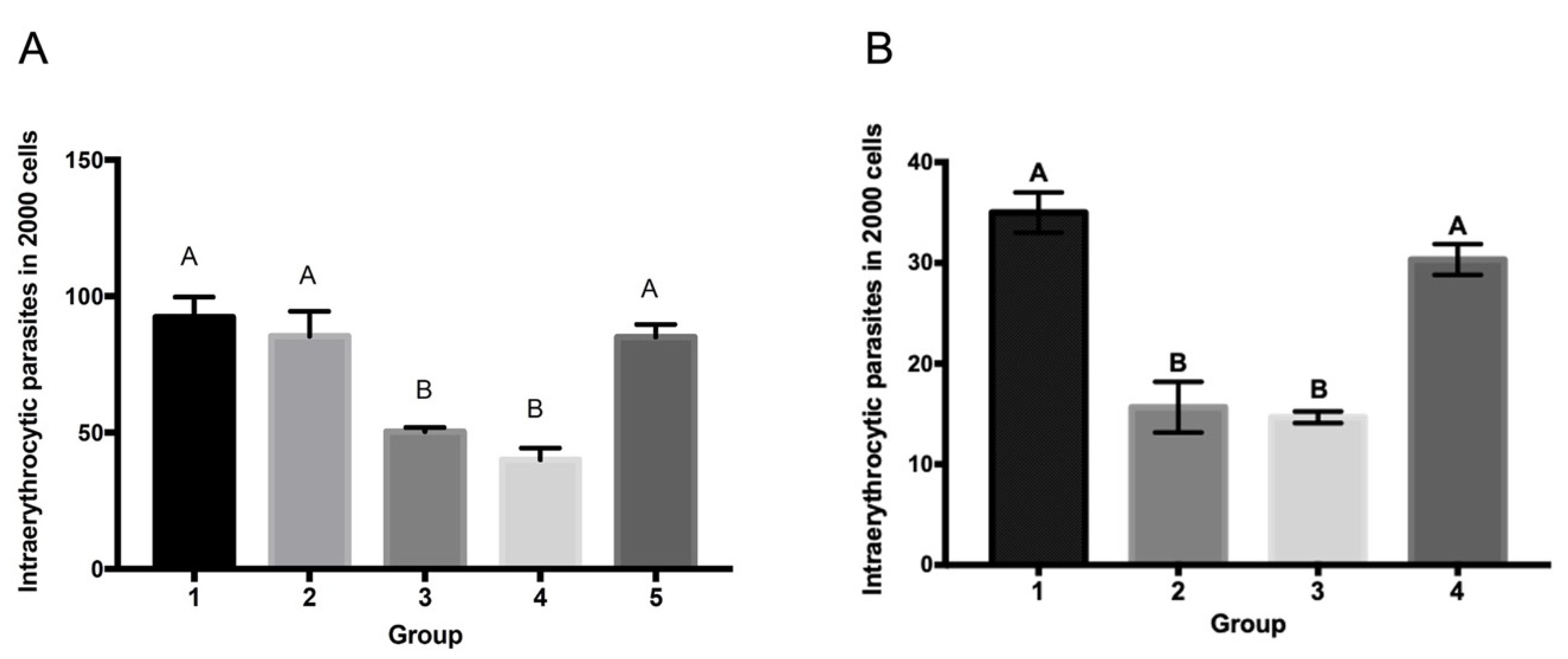
3.4. Anti-SBP4 Antibodies Are Capable of Neutralizing B. bigemina and B. ovata’s Parasite Growth In Vitro
3.5. SBP4 Peptides Containing Conserved Epitopes Are Recognized by Antibodies from Cattle Naturally Infected with B. bigemina
3.6. Antibodies to SBP4 Peptides Containing Conserved Epitopes Neutralize B. bigemina Parasite Growth In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ristic, M. Current Topics in Veterinary Medicine and Animal Science. In Diseases of Cattle in the Tropics; Ristic, M., McIntyre, I., Eds.; Martinus Nijhoff Publishers: Hague, The Netherlands, 1981; Volume 6, pp. 443–468. [Google Scholar]
- Sivakumar, T.; Igarashi, I.; Yokoyama, N. Babesia ovata: Taxonomy, phylogeny and epidemiology. Vet. Parasitol. 2016, 229, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.A.; Rodriguez, M.; Cursino-Santos, J.R. Babesia and red cell invasion. Curr. Opin. Hematol. 2012, 19, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, N.; Okamura, M.; Igarashi, I. Erythrocyte invasion by Babesia parasites: Current advances in the elucidation of the molecular interactions between the protozoan ligands and host receptors in the invasion stage. Vet. Parasitol. 2006, 138, 22–32. [Google Scholar] [CrossRef]
- Potgieter, F.T.; Els, H.J. The fine structure of intra-erythrocytic stages of Babesia bigemina. Onderstepoort J. Vet. Res. 1977, 44, 157–168. [Google Scholar]
- Hines, S.A.; Palmer, G.H.; Brown, W.C.; McElwain, T.F.; Suarez, C.E.; Vidotto, O.; Rice-Ficht, A.C. Genetic and antigenic characterization of Babesia bovis merozoite spherical body protein Bb-1. Mol. Biochem. Parasitol. 1995, 69, 149–159. [Google Scholar] [CrossRef]
- Dowling, S.C.; Perryman, L.E.; Jasmer, D.P. A Babesia bovis 225-kilodalton spherical-body protein: Localization to the cytoplasmic face of infected erythrocytes after merozoite invasion. Infect. Immun. 1996, 64, 2618–2626. [Google Scholar] [CrossRef] [Green Version]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. A unique Babesia bovis spherical body protein is conserved among geographic isolates and localizes to the infected erythrocyte membrane. Mol. Biochem. Parasitol. 2000, 105, 1–12. [Google Scholar]
- Masatani, T.; Matsuo, T.; Tanaka, T.; Terkawi, M.A.; Lee, E.-G.; Goo, Y.-K.; Aboge, G.O.; Yamagishi, J.; Hayashi, K.; Kameyama, K.; et al. TgGRA23, a novel Toxoplasma gondii dense granule protein associated with the parasitophorous vacuole membrane and intravacuolar network. Parasitol. Int. 2013, 62, 372–379. [Google Scholar] [CrossRef]
- Ramakrishnan, C.; Walker, R.A.; Eichenberger, R.M.; Hehl, A.; Smith, N. nThe merozoite-specific protein, TgGRA11B, identified as a component of the Toxoplasma gondii parasitophorous vacuole in a tachyzoite expression model. Int. J. Parasitol. 2017, 47, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Okada, T.; Marmansari, D.; Li, Z.-M.; Adilbish, A.; Canko, S.; Ueno, A.; Shono, H.; Furuoka, H.; Igarashi, M. A novel dense granule protein, GRA22, is involved in regulating parasite egress in Toxoplasma gondii. Mol. Biochem. Parasitol. 2013, 189, 5–13. [Google Scholar] [CrossRef]
- Gold, D.A.; Kaplan, A.D.; Lis, A.; Bett, G.C.; Rosowski, E.E.; Cirelli, K.M.; Bougdour, A.; Sidik, S.M.; Beck, J.R.; Lourido, S.; et al. The Toxoplasma Dense Granule Proteins GRA17 and GRA23 Mediate the Movement of Small Molecules between the Host and the Parasitophorous Vacuole. Cell Host Microbe 2015, 17, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosqueda, J.; Hidalgo-Ruiz, M.; Calvo-Olvera, D.A.; Hernandez-Silva, D.J.; Ueti, M.W.; Mercado-Uriostegui, M.A.; Rodriguez, A.; Ramos-Aragon, J.A.; Hernandez-Ortiz, R.; Kawazu, S.-I.; et al. RON2, a novel gene in Babesia bigemina, contains conserved, immunodominant B-cell epitopes that induce antibodies that block merozoite invasion. Parasitology 2019, 146, 1646–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Owczarzy, R.; Tataurov, A.V.; Wu, Y.; Manthey, J.A.; McQuisten, K.A.; Almabrazi, H.G.; Pedersen, K.F.; Lin, Y.; Garretson, J.; McEntaggart, N.O.; et al. IDT SciTools: A suite for analysis and design of nucleic acid oligomers. Nucleic Acids Res. 2008, 36, W163–W169. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Raghava, G.P.S. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.E.P.; Lund, O.; Nielsen, M. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosqueda, J.; McElwain, T.F.; Palmer, G.H. Babesia bovis merozoite surface antigen 2 proteins are expressed on the merozoite and sporozoite surface, and specific antibodies inhibit attachment and invasion of erythrocytes. Infect. Immun. 2002, 70, 6448–6455. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Dang-Trinh, M.A.; Higuchi, L.; Mosqueda, J.; Hakimi, H.; Asada, M.; Yamagishi, J.; Umemiya-Shirafuji, R.; Kawazu, S.I. Initiated Babesia ovata Sexual Stages under In Vitro Conditions Were Recognized by Anti-CCp2 Antibodies, Showing Changes in the DNA Content by Imaging Flow Cytometry. Pathogens 2019, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado-Uriostegui, M.A.; Castro-Sánchez, L.A.; Batiha, G.E.-S.; Valdez-Espinoza, U.M.; Falcón-Neri, A.; Ramos-Aragon, J.A.; Hernández-Ortiz, R.; Kawazu, S.-I.; Igarashi, I.; Mosqueda, J. The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates. Pathogens 2022, 11, 591. [Google Scholar] [CrossRef]
- Hernández-Silva, D.J.; Valdez-Espinoza, U.M.; Mercado-Uriostegui, M.A.; Aguilar-Tipacamú, G.; Ramos-Aragón, J.A.; Hernández-Ortiz, R.; Ueti, M.; Mosqueda, J. Immunomolecular Characterization of MIC-1, a Novel Antigen in Babesia bigemina, Which Contains Conserved and Immunodominant B-Cell Epitopes that Induce Neutralizing Antibodies. Vet. Sci. 2018, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Weber, G. Ultrastructure and cytochemistry of the pellicle and apical complexes of the kinete of Babesia bigemina and Babesia ovis in the hemolymph and oavry of the tick. J. Protozool. 1980, 27, 59–71. [Google Scholar] [CrossRef]
- Vichido, R.; Falcon, A.; Ramos, J.A.; Alvarez, A.; Figueroa, J.V.; Norimine, J.; Brown, W.C.; Castro, L.A.; Mosqueda, J. Expression analysis of heat shock protein 20 and rhoptry-associated protein 1a in sexual stages and kinetes of Babesia bigemina. Ann. N. Y. Acad. Sci. 2008, 1149, 136–140. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Huyen, N.X.; Wibowo, P.E.; Seuseu, F.J.; Aboulaila, M.; Ueno, A.; Goo, Y.-K.; Yokoyama, N.; Xuan, X.; Igarashi, I. Spherical body protein 4 is a new serological antigen for global detection of Babesia bovis infection in cattle. Clin. Vaccine Immunol. 2011, 18, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Li, M.; Sun, Y.; Yu, L.; He, P.; Nie, Z.; Zhan, X.; Zhao, Y.; Luo, X.; Wang, S.; et al. Characterization of a novel secretory spherical body protein in Babesia orientalis and Babesia orientalis-infected erythrocytes. Parasit. Vectors 2018, 11, 433. [Google Scholar] [CrossRef] [PubMed]
- Terkawi, M.A.; Seuseu, F.J.; Eko-Wibowo, P.; Huyen, N.X.; Minoda, Y.; AbouLaila, M.; Kawai, S.; Yokoyama, N.; Xuan, X.; Igarashi, I. Secretion of a new spherical body protein of Babesia bovis into the cytoplasm of infected erythrocytes. Mol. Biochem. Parasitol. 2011, 178, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Guillemi, E.; Ruybal, P.; Lia, V.; González, S.; Farber, M.; Wilkowsky, S.E. Multi-locus typing scheme for Babesia bovis and Babesia bigemina reveals high levels of genetic variability in strains from Northern Argentina. Infect. Genet. Evol. 2013, 14, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Ruiz, M.; Suarez, C.E.; Mercado-Uriostegui, M.A.; Hernandez-Ortiz, R.; Ramos, J.A.; Galindo-Velasco, E.; Leon-Avila, G.; Hernandez, J.M.; Mosqueda, J. Babesia bovis RON2 contains conserved B-cell epitopes that induce an invasion-blocking humoral immune response in immunized cattle. Parasit. Vectors 2018, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Barreda, D.; Hidalgo-Ruiz, M.; Hernandez-Ortiz, R.; Ramos, J.A.; Galindo-Velasco, E.; Mosqueda, J. Identification of conserved peptides containing B-cell epitopes of Babesia bovis AMA-1 and their potential as diagnostics candidates. Transbound. Emerg. Dis. 2020, 67, 60–68. [Google Scholar] [CrossRef]
- Hidalgo-Ruiz, M.; Mejia-López, S.; Pérez-Serrano, R.M.; de Larrea, G.Z.-L.; Ganzinelli, S.; Florin-Christensen, M.; Suarez, C.E.; Hernández-Ortiz, R.; Mercado-Uriostegui, M.A.; Rodríguez-Torres, A.; et al. Babesia bovis AMA-1, MSA-2c and RAP-1 contain conserved B and T-cell epitopes, which generate neutralizing antibodies and a long-lasting Th1 immune response in vaccinated cattle. Vaccine 2022, 40, 1108–1115. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Position in aa | Sequence |
|---|---|---|
| SBP4-1 | 51–70 | MIHRQTDGCAPRTPVVYTPV |
| SBP4-2 | 215–234 | KLVALIYHDVDGMKEALYHG |
| SBP4-3 | 145–163 | KTIVVDINDVNDNKYLSYE |
| SBP4-4 | 71–89 | RPNKLRHLVWSDTVIHGVG |
| Peptide 1 | Peptide 2 | Peptide 3 | Peptide 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Region | Farm | + | − | + | − | + | − | + | − |
| Aguascalientes | Villa Guadalupe | 9 | 1 | 7 | 3 | 10 | 0 | 10 | 0 |
| Las Palomas | 24 | 0 | 15 | 9 | 22 | 2 | 24 | 0 | |
| Granja María I | 4 | 0 | 2 | 2 | 3 | 1 | 4 | 0 | |
| Sinaloa | El Torito | 7 | 0 | 6 | 1 | 7 | 0 | 7 | 0 |
| El Moral | 10 | 0 | 8 | 2 | 10 | 0 | 10 | 0 | |
| Herradura | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | |
| El Barón | 2 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | |
| Veracruz | La Esperanza | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 |
| San Faudila | 2 | 2 | 4 | 0 | 4 | 0 | 4 | 0 | |
| Las Torres | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | |
| Irineo Murillo | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | |
| Playa Vicente | 2 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | |
| El Arbolito | 2 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | |
| Manuel Antonio | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | |
| Buenos Aires | 2 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | |
| El Orijuelo | 9 | 0 | 9 | 0 | 8 | 1 | 9 | 0 | |
| Queretaro | Granja Araceli | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 |
| Amazcala | 0 | 7 | 1 | 6 | 0 | 7 | 0 | 7 | |
| Los Moreno | 3 | 7 | 4 | 6 | 3 | 7 | 3 | 7 | |
| Durango | Arroyo Seco | 2 | 33 | 1 | 34 | 2 | 33 | 2 | 33 |
| Total | 86 | 50 | 69 | 67 | 85 | 51 | 89 | 47 | |
| True positive | 81 | 63 | 80 | 84 | |||||
| False positve | 5 | 6 | 5 | 5 | |||||
| False negative | 3 | 21 | 4 | 0 | |||||
| True negative | 47 | 46 | 47 | 47 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosqueda, J.; Hernandez-Silva, D.J.; Ueti, M.W.; Cruz-Reséndiz, A.; Marquez-Cervantez, R.; Valdez-Espinoza, U.M.; Dang-Trinh, M.-A.; Nguyen, T.-T.; Camacho-Nuez, M.; Mercado-Uriostegui, M.A.; et al. Spherical Body Protein 4 from Babesia bigemina: A Novel Gene That Contains Conserved B-Cell Epitopes and Induces Cross-Reactive Neutralizing Antibodies in Babesia ovata. Pathogens 2023, 12, 495. https://doi.org/10.3390/pathogens12030495
Mosqueda J, Hernandez-Silva DJ, Ueti MW, Cruz-Reséndiz A, Marquez-Cervantez R, Valdez-Espinoza UM, Dang-Trinh M-A, Nguyen T-T, Camacho-Nuez M, Mercado-Uriostegui MA, et al. Spherical Body Protein 4 from Babesia bigemina: A Novel Gene That Contains Conserved B-Cell Epitopes and Induces Cross-Reactive Neutralizing Antibodies in Babesia ovata. Pathogens. 2023; 12(3):495. https://doi.org/10.3390/pathogens12030495
Chicago/Turabian StyleMosqueda, Juan, Diego Josimar Hernandez-Silva, Massaro W. Ueti, Adolfo Cruz-Reséndiz, Ricardo Marquez-Cervantez, Uriel Mauricio Valdez-Espinoza, Minh-Anh Dang-Trinh, Thu-Thuy Nguyen, Minerva Camacho-Nuez, Miguel Angel Mercado-Uriostegui, and et al. 2023. "Spherical Body Protein 4 from Babesia bigemina: A Novel Gene That Contains Conserved B-Cell Epitopes and Induces Cross-Reactive Neutralizing Antibodies in Babesia ovata" Pathogens 12, no. 3: 495. https://doi.org/10.3390/pathogens12030495
APA StyleMosqueda, J., Hernandez-Silva, D. J., Ueti, M. W., Cruz-Reséndiz, A., Marquez-Cervantez, R., Valdez-Espinoza, U. M., Dang-Trinh, M. -A., Nguyen, T. -T., Camacho-Nuez, M., Mercado-Uriostegui, M. A., Aguilar-Tipacamú, G., Ramos-Aragon, J. A., Hernandez-Ortiz, R., Kawazu, S. -i., & Igarashi, I. (2023). Spherical Body Protein 4 from Babesia bigemina: A Novel Gene That Contains Conserved B-Cell Epitopes and Induces Cross-Reactive Neutralizing Antibodies in Babesia ovata. Pathogens, 12(3), 495. https://doi.org/10.3390/pathogens12030495