
The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Chemicals
2.2. Bacterial Culture and Identifications
2.3. Survival of O. nipae Larvae Infected with P. luminescens H06 and X. nematophila All
2.4. Phenoloxidase (PO) Enzyme Activity Assay
2.5. Recognitions of P. luminescens H06 and X. nematophila All by O. nipae’s Immune System
2.6. Effects of P. luminescens H06 and X. nematophila All Infections on O. nipae’s Prophenoloxidase Activation System
2.7. Influence of Eicosanoid Inhibitors and Precursor on Phenoloxidase Activity in O. nipae
2.8. Effect of Eicosanoid Biosynthesis Inhibitor and Precursor on O. nipae PPO Gene Expression
2.9. Statistical Analysis
3. Results
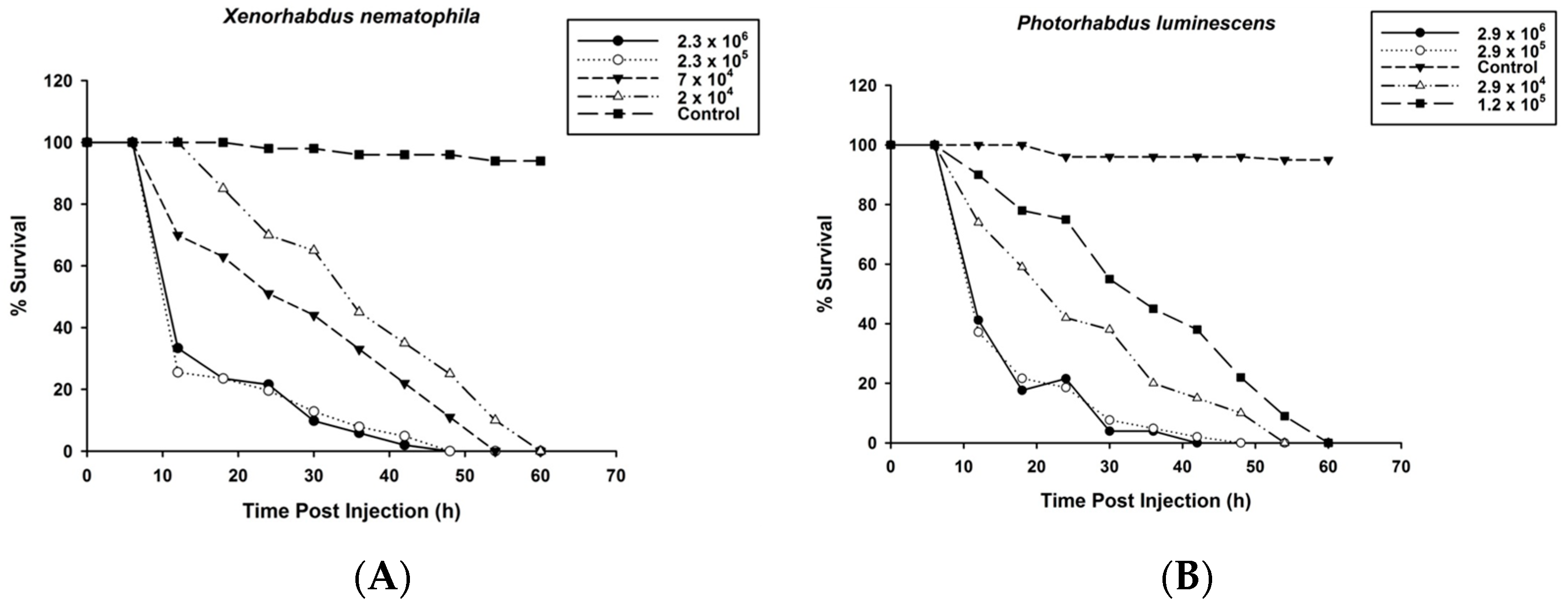
3.1. Survival of O. nipae Larvae Infected with P. luminescens H06 and X. nematophila All
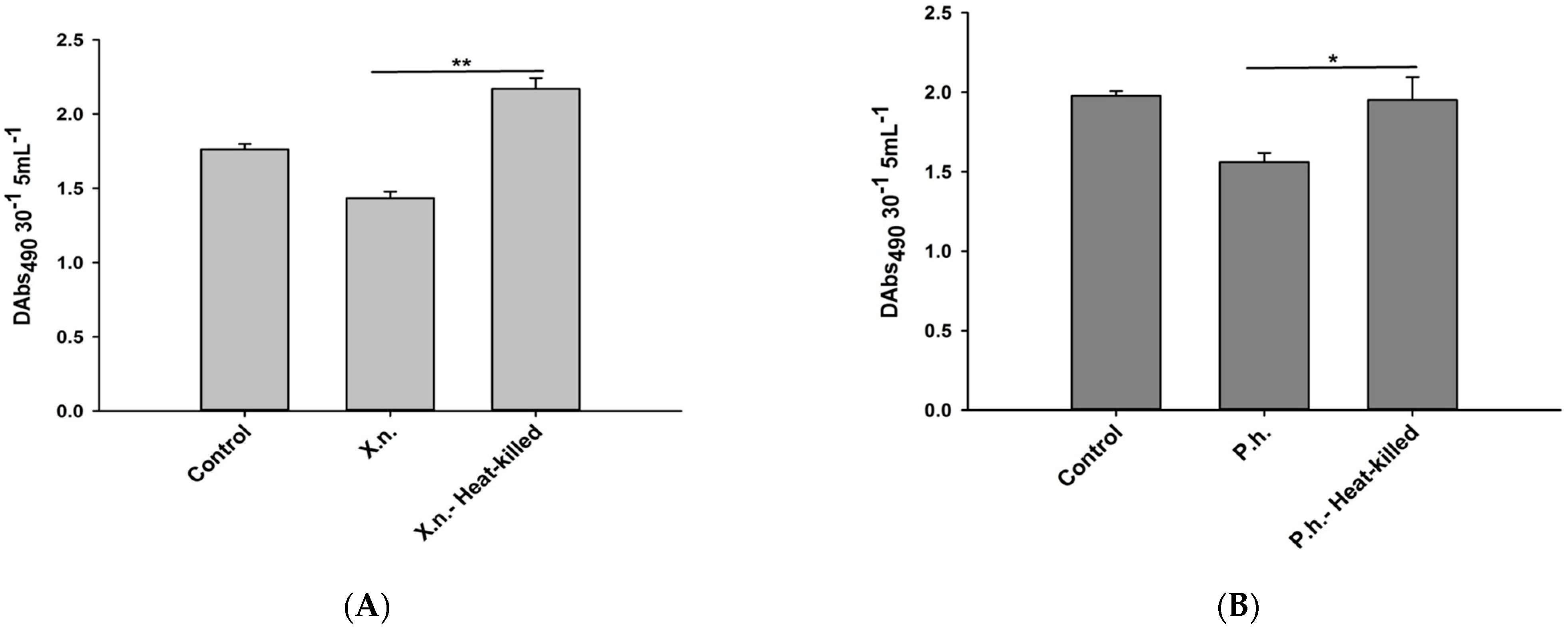
3.2. Inhibitory Effects of P. luminescens H06 and X. nematophila All against the Phenoloxidase Activity in O. nipae
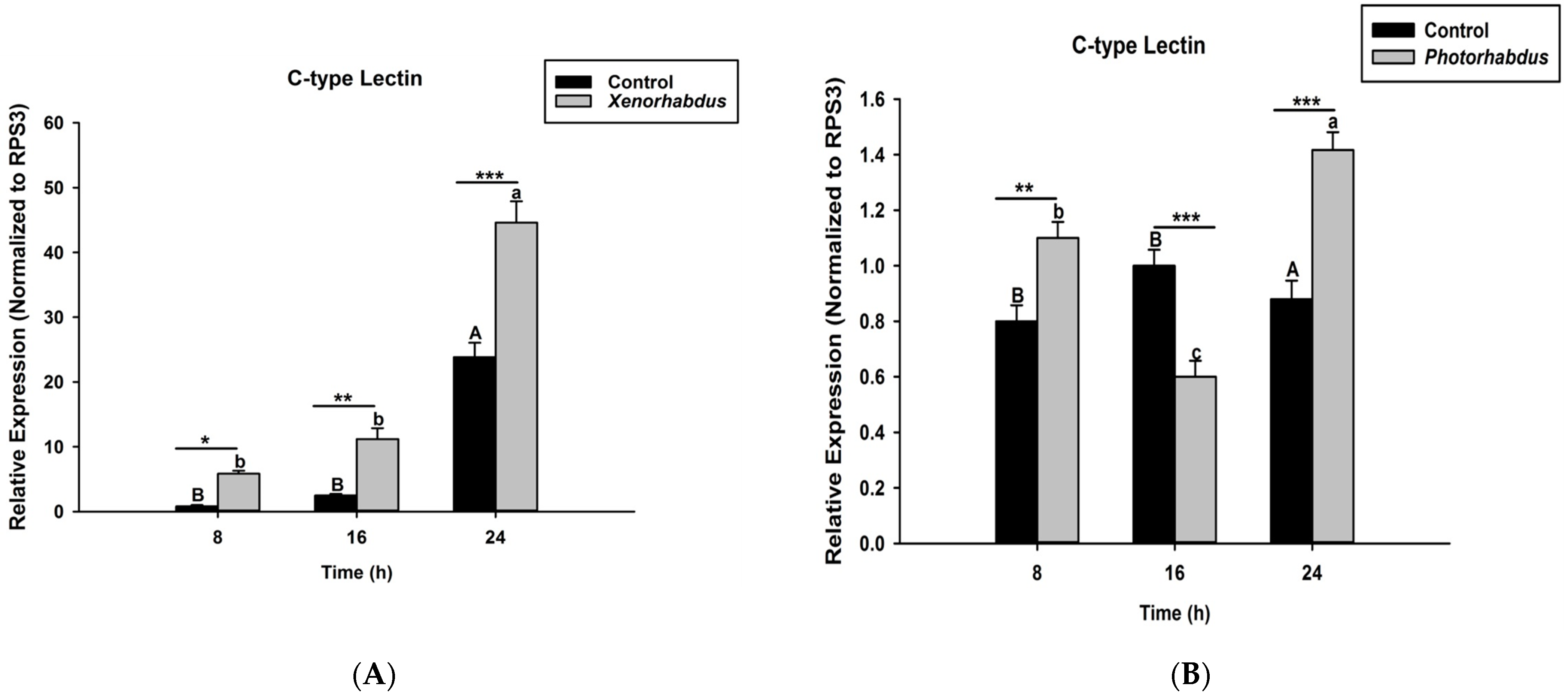
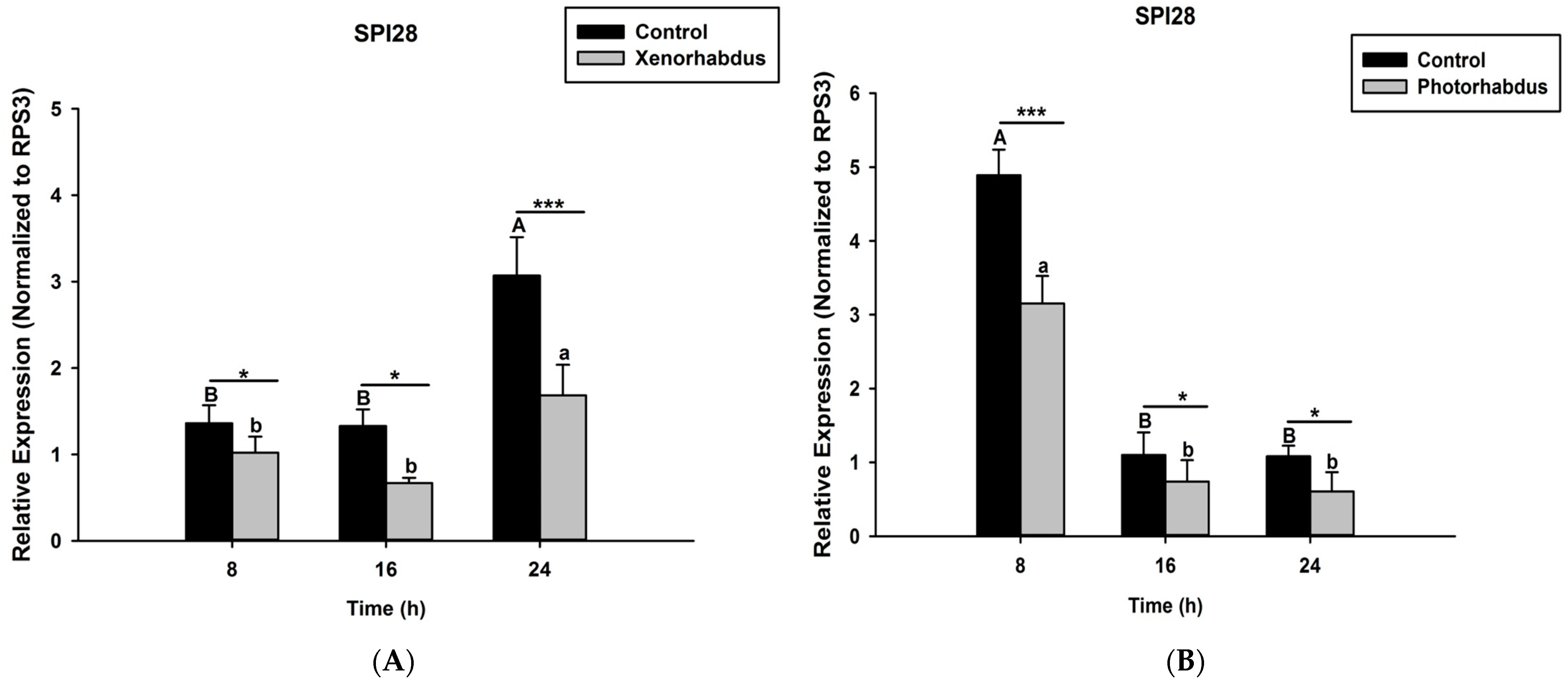
3.3. P. luminescens H06 and X. nematophila All Induce the Expression of Recognition Gene by O. nipae’s Immune System
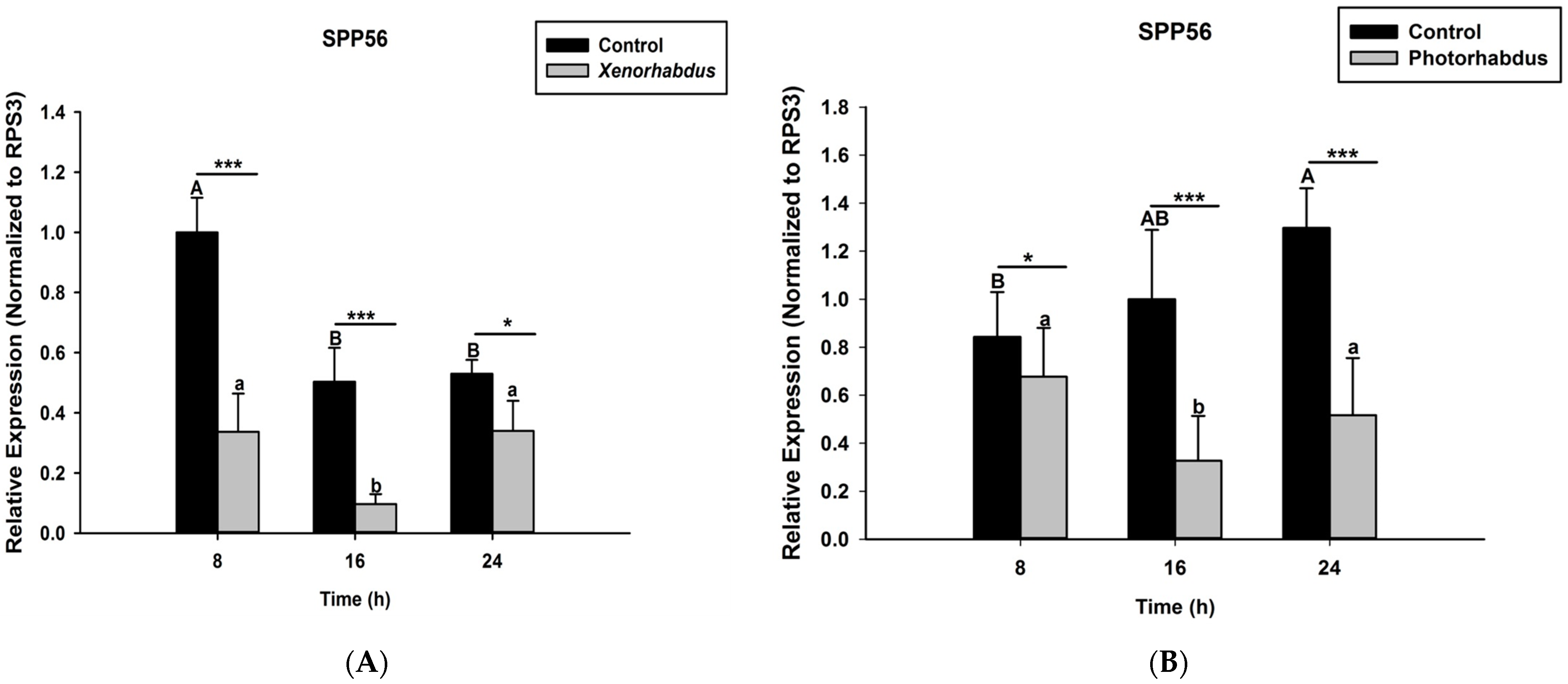
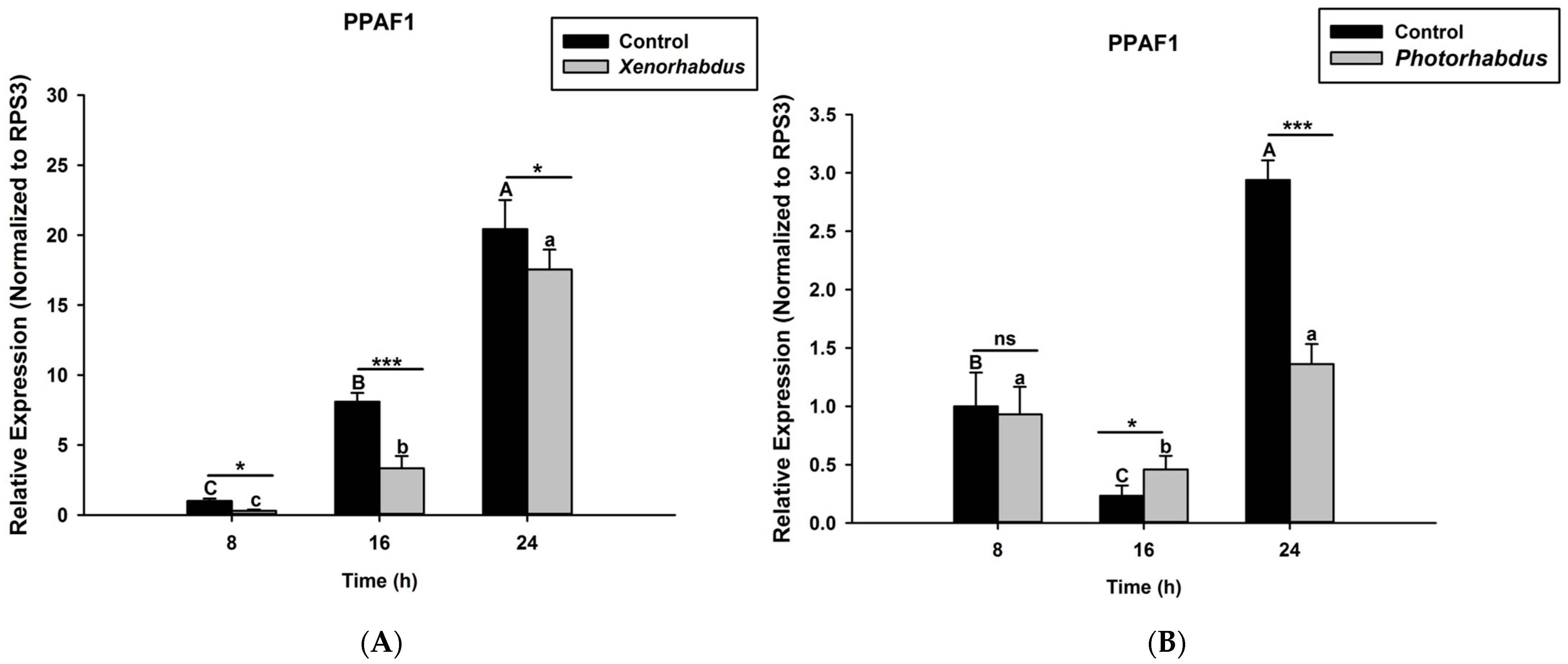
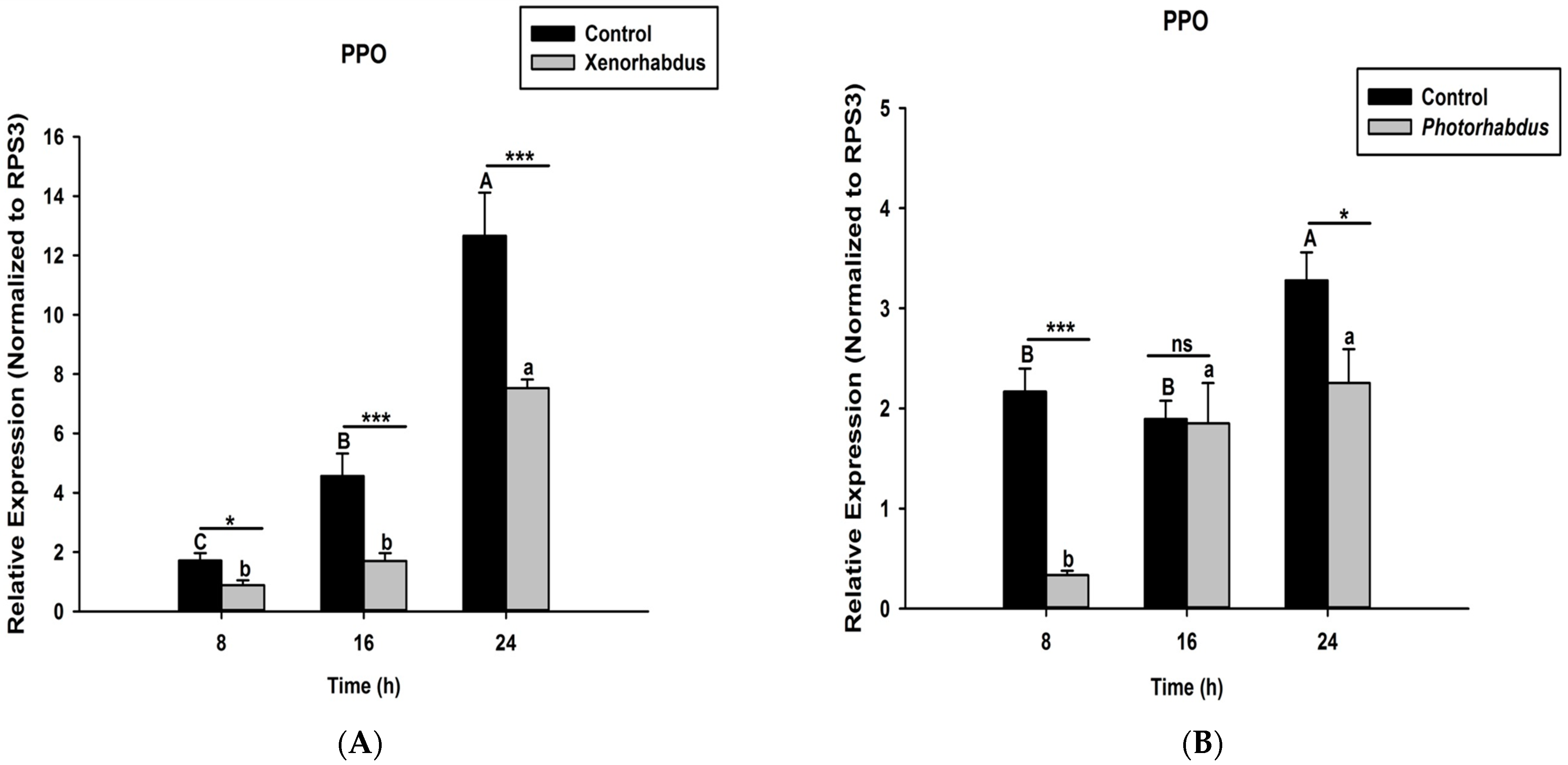
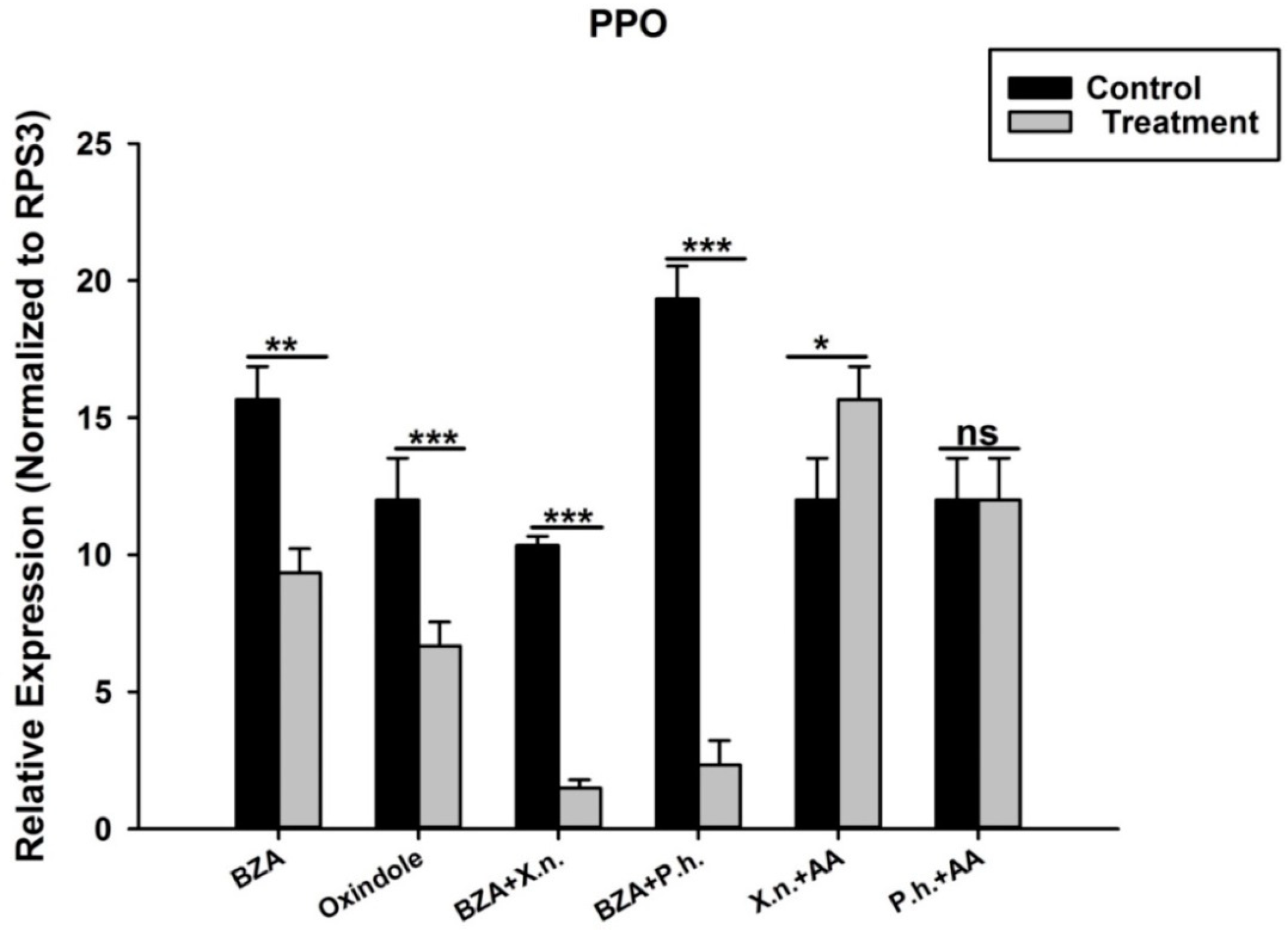
3.4. P. luminescens H06 and X. nematophila All Down-Regulate the Expression of Prophenoloxidase Genes in O. nipae
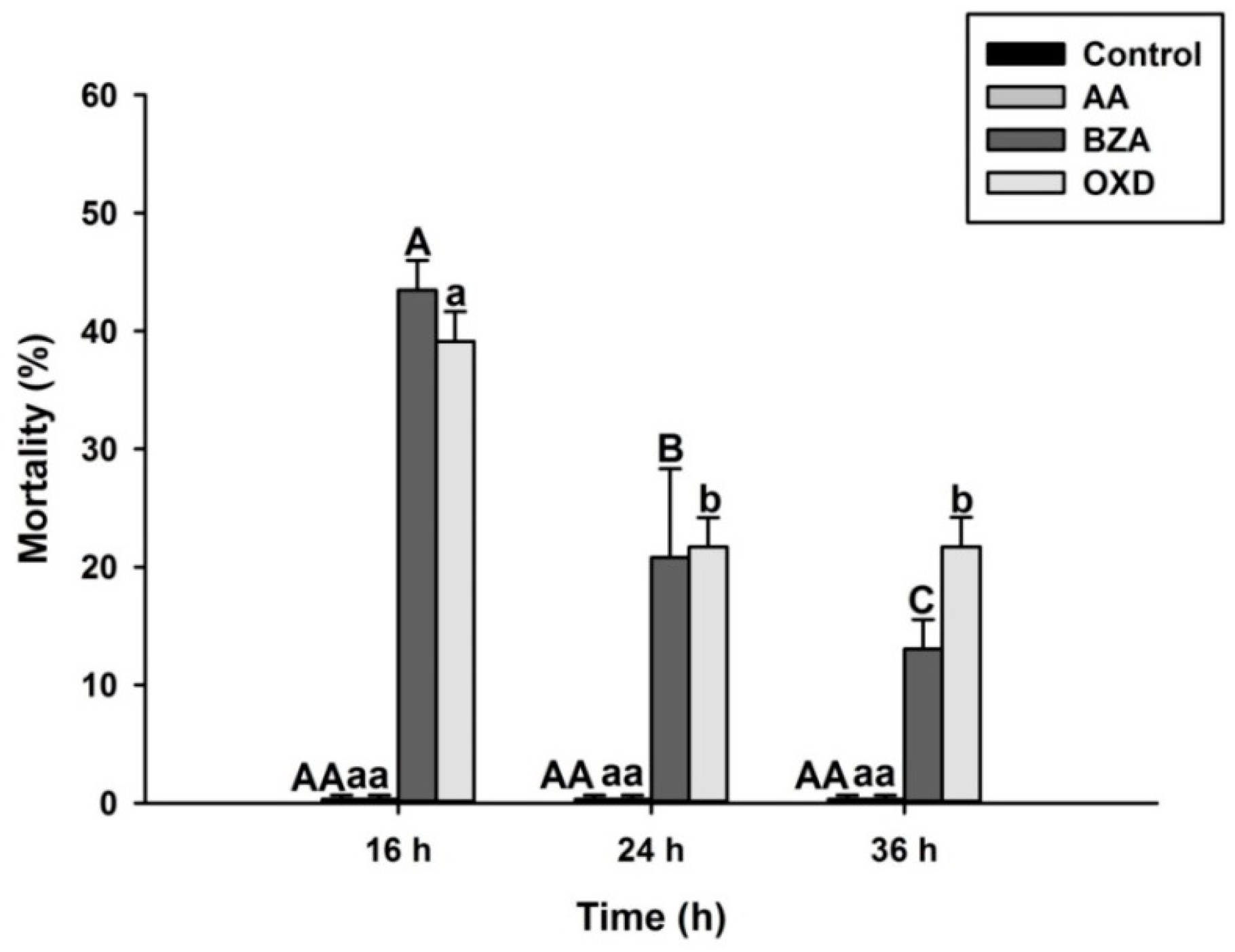
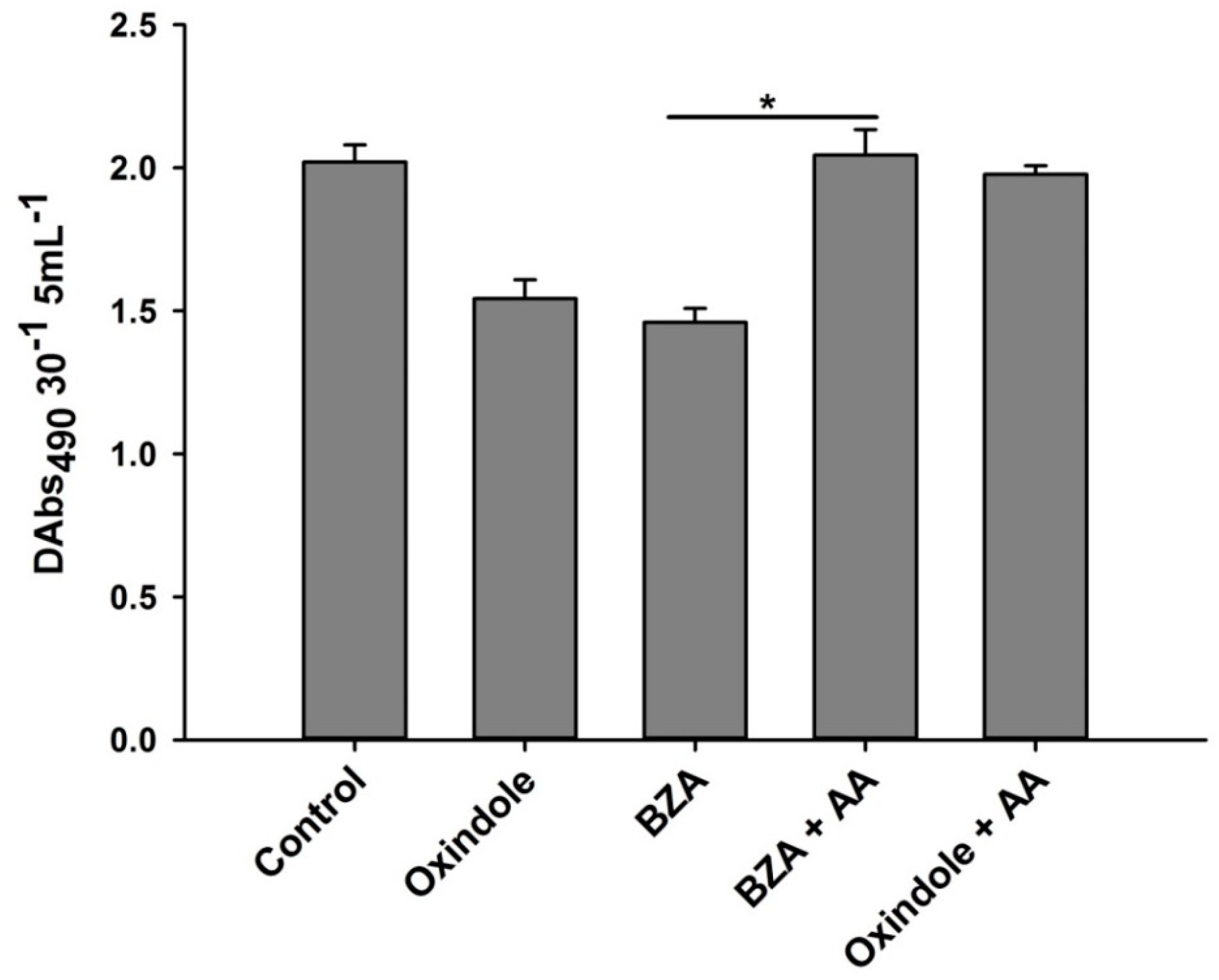
3.5. Addition of Arachidonic Acid Restores the Inhibitory Effects of the Eicosanoid Inhibitors on Phenoloxidase Enzyme Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanda, N.B.; Hou, B.; Muhammad, A.; Ali, H.; Hou, Y. Exploring the role of Relish on antimicrobial peptide expressions (AMPs) upon nematode-bacteria complex challenge in the Nipa Palm Hispid Beetle, Octodonta nipae Maulik (Coleoptera: Chrysomelidae). Front. Microbiol. 2019, 10, 2466. [Google Scholar] [CrossRef] [PubMed]
- Sanda, N.B.; Hou, B.; Hou, Y. The Entomopathogenic Nematodes H. bacteriophora and S. carpocapsae Inhibit the Activation of proPO System of the Nipa Palm Hispid Octodonta nipae (Coleoptera: Chrysomelidae). Life 2022, 12, 1019. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.-F.; Li, J.-L.; Hou, Y.-M.; Zhang, X. Descriptions of immature stages of Octodonta nipae (Maulik) (Coleoptera, Chrysomelidae, Cassidinae, Cryptonychini). ZooKeys 2018, 764, 91–109. [Google Scholar] [CrossRef]
- Sanda, N.; Wudil, B.; Hou, Y. Effect of Temperatures on the Pathogenicity of Entomopathogenic Nematodes (Steinernema carpocapsae and Heterorhabditis bacteriophora) for Biocontrol of Octodonta nipae (Maulik). Niger. J. Entomol. 2020, 36, 88–95. [Google Scholar] [CrossRef]
- Sanda, N.B.; Muhammad, A.; Ali, H.; Hou, Y. Entomopathogenic nematode Steinernema carpocapsae surpasses the cellular immune responses of the hispid beetle, Octodonta nipae (Coleoptera: Chrysomelidae). Microb. Pathog. 2018, 124, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Muhammad, A.; Islam, S.U.; Islam, W.; Hou, Y. A novel bacterial symbiont association in the hispid beetle, Octodonta nipae (Coleoptera: Chrysomelidae), their dynamics and phylogeny. Microb. Pathog. 2018, 118, 378–386. [Google Scholar] [CrossRef]
- Meng, E.; Qiao, T.; Tang, B.; Hou, Y.; Yu, W.; Chen, Z. Effects of ovarian fluid, venom and egg surface characteristics of Tetrastichus brontispae (Hymenoptera: Eulophidae) on the immune response of Octodonta nipae (Coleoptera: Chrysomelidae). J. Insect Physiol. 2018, 109, 125–137. [Google Scholar] [CrossRef]
- Tang, B.; Chen, J.; Hou, Y.; Meng, E. Transcriptome immune analysis of the invasive beetle Octodonta nipae (Maulik) (Coleoptera: Chrysomelidae) parasitized by Tetrastichus brontispae Ferrière (Hymenoptera: Eulophidae). PLoS ONE 2014, 9, e91482. [Google Scholar] [CrossRef]
- Lacey, L.; Grzywacz, D.; Shapiro-Ilan, D.; Frutos, R.; Brownbridge, M.; Goettel, M. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Gutiérrez, J.D.; Castelblanco, A.; Rodríguez-Bocanegra, M.X.; Teran, W.; Sáenz-Aponte, A. Photorhabdus luminescens subsp. akhurstii SL0708 pathogenicity in Spodoptera frugiperda (Lepidoptera: Noctuidae) and Galleria mellonella (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2017, 20, 1112–1121. [Google Scholar] [CrossRef]
- Froy, O. Convergent evolution of invertebrate defensins and nematode antibacterial factors. Trends Microbiol. 2005, 13, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Binda-Rossetti, S.; Mastore, M.; Protasoni, M.; Brivio, M.F. Effects of an entomopathogen nematode on the immune response of the insect pest red palm weevil: Focus on the host antimicrobial response. J. Invertebr. Pathol. 2016, 133, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Abdolmaleki, A.; Maafi, Z.; Dastjerdi, H.; Naseri, B.; Ghasemi, A. Immune defense of Pieris brassicae larvae in challenged with Heterorhabditis bacteriophora, its symbiotic bacteria and metabolites. Invertebr. Surviv. J. 2017, 14, 73–84. [Google Scholar]
- Herbert, E.E.; Goodrich-Blair, H. Friend and foe: The two faces of Xenorhabdus nematophila. Nat. Rev. Microbiol. 2007, 5, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Mastore, M.; Brivio, M.F. Cuticular surface lipids are responsible for disguise properties of an entomoparasite against host cellular responses. Dev. Comp. Immunol. 2008, 32, 1050–1062. [Google Scholar] [CrossRef]
- Dillman, A.R.; Chaston, J.M.; Adams, B.J.; Ciche, T.A.; Goodrich-Blair, H.; Stock, S.P.; Sternberg, P.W. An entomopathogenic nematode by any other name. PLoS Pathog. 2012, 8, e1002527. [Google Scholar] [CrossRef] [Green Version]
- Goodrich-Blair, H. They’ve got a ticket to ride: Xenorhabdus nematophila–Steinernema carpocapsae symbiosis. Curr. Opin. Microbiol. 2007, 10, 225–230. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.; Waterfield, N.; Daborn, P.; Joyce, S.; Bennett, H.; Au, C.; Dowling, A.; Boundy, S.; Reynolds, S.; Clarke, D. Photorhabdus: Towards a functional genomic analysis of a symbiont and pathogen. FEMS Microbiol. Rev. 2003, 26, 433–456. [Google Scholar] [CrossRef]
- Blackburn, M.B.; Domek, J.M.; Gelman, D.B.; Hu, J.S. The broadly insecticidal Photorhabdus luminescens toxin complex a (Tca): Activity against the Colorado potato beetle, Leptinotarsa decemlineata, and sweet potato whitefly, Bemisia tabaci. J. Insect Sci. 2005, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, E.; Upadhyay, D.; Mandjiny, S.; Bullard-Dillard, R.; Storms, M.; Menefee, M.; Holmes, L.D. Photorhabdus luminescens: Virulent properties and agricultural applications. Am. J. Agric. For. 2015, 3, 171–177. [Google Scholar] [CrossRef]
- Shi, M.; Chen, X.-Y.; Zhu, N.; Chen, X.-X. Molecular identification of two prophenoloxidase-activating proteases from the hemocytes of Plutella xylostella (Lepidoptera: Plutellidae) and their transcript abundance changes in response to microbial challenges. J. Insect Sci. 2014, 14, 179. [Google Scholar] [CrossRef] [Green Version]
- Bisch, G.; Pagès, S.; McMullen II, J.G.; Stock, S.P.; Duvic, B.; Givaudan, A.; Gaudriault, S. Xenorhabdus bovienii CS03, the bacterial symbiont of the entomopathogenic nematode Steinernema weiseri, is a non-virulent strain against lepidopteran insects. J. Invertebr. Pathol. 2015, 124, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Song, C.J.; Seo, S.; Shrestha, S.; Kim, Y. Bacterial metabolites of an 694 entomopathogenic bacterium, Xenorhabdus nematophila, inhibit a catalytic 695 activity of phenoloxidase of the diamondback moth, Plutella xylostella. J 2011, 696, 317–322. [Google Scholar]
- Ullah, I.; Khan, A.L.; Ali, L.; Khan, A.R.; Waqas, M.; Lee, I.-J.; Shin, J.-H. An insecticidal compound produced by an insect-pathogenic bacterium suppresses host defenses through phenoloxidase inhibition. Molecules 2014, 19, 20913–20928. [Google Scholar] [CrossRef] [Green Version]
- Eleftherianos, I.; Reynolds, S.E. A nematode symbiont sheds light on invertebrate immunity. Trends Parasitol. 2007, 23, 514–517. [Google Scholar]
- Kim, Y.; Ji, D.; Cho, S.; Park, Y. Two groups of entomopathogenic bacteria, Photorhabdus and Xenorhabdus, share an inhibitory action against phospholipase A2 to induce host immunodepression. J. Invertebr. Pathol. 2005, 89, 258–264. [Google Scholar] [CrossRef]
- Stanley, D.; Miller, J.; Tunaz, H. Eicosanoid actions in insect immunity. J. Innate Immun. 2009, 1, 282–290. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y. Xenorhabdus nematophilus inhibits p-bromophenacyl bromide (BPB)-sensitive PLA2 of Spodoptera exigua. Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2003, 54, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.M.; Portmann, C.; Zhang, X.; Roeffaers, M.B.; Clardy, J. Small molecule perimeter defense in entomopathogenic bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 10821–10826. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Miao, Y.; Zhang, Z. Leaf consumption capacity and damage projection of Octodonta nipae (Coleoptera: Chrysomelidae) on three palm species. Ann. Entomol. Soc. Am. 2014, 107, 1010–1017. [Google Scholar] [CrossRef]
- Hou, Y.; Miao, Y.; Zhang, Z. Study on life parameters of the invasive species Octodonta nipae (Coleoptera: Chrysomelidae) on different palm species, under laboratory conditions. J. Econ. Entomol. 2014, 107, 1486–1495. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Weng, Z. Temperature-dependent development and life table parameters of Octodonta nipae (Coleoptera: Chrysomelidae). Environ. Entomol. 2010, 39, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wu, Z.; Wang, C.; Xie, L. The status and harm of invasive insects in Fujian, China. In Biological Invasions: Problems and Countermeasures; Science Press: Beijing, China, 2011; pp. 111–114. [Google Scholar]
- Ali, H.; Muhammad, A.; Sanda Bala, N.; Hou, Y. The endosymbiotic Wolbachia and host COI gene enables to distinguish between two invasive palm pests; coconut leaf beetle, Brontispa longissima and hispid leaf beetle, Octodonta nipae. J. Econ. Entomol. 2018, 111, 2894–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukruksa, C.; Yimthin, T.; Suwannaroj, M.; Muangpat, P.; Tandhavanant, S.; Thanwisai, A.; Vitta, A. Isolation and identification of Xenorhabdus and Photorhabdus bacteria associated with entomopathogenic nematodes and their larvicidal activity against Aedes aegypti. Parasites Vectors 2017, 10, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahoo, A.M.; Mukhtar, T.; Gowen, S.R.; Pembroke, B. Virulence of entomopathogenic bacteria Xenorhabdus bovienii and Photorhabdus luminescens against Galleria mellonella larvae. Pak. J. Zool. 2011, 43, 543–548. [Google Scholar]
- Sadekuzzaman, M.; Park, Y.; Lee, S.; Kim, K.; Jung, J.K.; Kim, Y. An entomopathogenic bacterium, Xenorhabdus hominickii ANU101, produces oxindole and suppresses host insect immune response by inhibiting eicosanoid biosynthesis. J. Invertebr. Pathol. 2017, 145, 13–22. [Google Scholar] [CrossRef]
- Abdel-Razek, A. Pathogenic effects of Xenorhabdus nematophilus and Photorhabdus luminescens (Enterobacteriaceae) against pupae of the Diamondback Moth, Plutella xylostella (L.). Anz. Schädlingskunde J. Pest Sci. 2003, 76, 108–111. [Google Scholar] [CrossRef]
- da Silva, O.S.; Prado, G.R.; da Silva, J.L.R.; Silva, C.E.; da Costa, M.; Heermann, R. Oral toxicity of Photorhabdus luminescens and Xenorhabdus nematophila (Enterobacteriaceae) against Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2013, 112, 2891–2896. [Google Scholar] [CrossRef]
- Bussaman, P.; Sobanboa, S.; Grewal, P.S.; Chandrapatya, A. Pathogenicity of additional strains of Photorhabdus and Xenorhabdus (Enterobacteriaceae) to the mushroom mite Luciaphorus perniciosus (Acari: Pygmephoridae). Appl. Entomol. Zool. 2009, 44, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Gökçen, F.; Felföldi, G.; Millichap, P.J.; Trenczek, T.E.; Ffrench-Constant, R.H.; Reynolds, S.E. The immunoglobulin family protein Hemolin mediates cellular immune responses to bacteria in the insect Manduca sexta. Cell. Microbiol. 2007, 9, 1137–1147. [Google Scholar] [CrossRef]
- Aliota, M.T.; Fuchs, J.F.; Rocheleau, T.A.; Clark, A.K.; Hillyer, J.F.; Chen, C.-C.; Christensen, B.M. Mosquito transcriptome profiles and filarial worm susceptibility in Armigeres subalbatus. PLoS Negl. Trop. Dis. 2010, 4, e666. [Google Scholar] [CrossRef] [Green Version]
- Kariuki, M.M.; Hearne, L.B.; Beerntsen, B.T. Differential transcript expression between the microfilariae of the filarial nematodes, Brugia malayi and B. pahangi. BMC Genom. 2010, 11, 225. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Daugherty, S.; Shetty, A.C.; Eleftherianos, I. RNAseq analysis of the Drosophila response to the entomopathogenic nematode Steinernema. G3 Genes Genomes Genet. 2017, 7, 1955–1967. [Google Scholar]
- Shi, H.; Zeng, H.; Yang, X.; Zhao, J.; Chen, M.; Qiu, D. An insecticidal protein from Xenorhabdus ehlersii triggers prophenoloxidase activation and hemocyte decrease in Galleria mellonella. Curr. Microbiol. 2012, 64, 604–610. [Google Scholar] [CrossRef]
- Yokoo, S.; Tojo, S.; Ishibashi, N. Suppression of the prophenoloxidase cascade in the larval haemolymph of the turnip moth, Agrotis segetum by an entomopathogenic nematode, Steinernema carpocapsae and its symbiotic bacterium. J. Insect Physiol. 1992, 38, 915–924. [Google Scholar] [CrossRef]
- Rahatkhah, Z.; Karimi, J.; Ghadamyari, M.; Brivio, M.F. Immune defenses of Agriotes lineatus larvae against entomopathogenic nematodes. BioControl 2015, 60, 641–653. [Google Scholar] [CrossRef]
- Darsouei, R.; Karimi, J.; Ghadamyari, M.; Hosseini, M. Differential change patterns of main antimicrobial peptide genes during infection of entomopathogenic nematodes and their symbiotic bacteria. J. Parasitol. 2017, 103, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Kim, Y. Various eicosanoids modulate the cellular and humoral immune responses of the beet armyworm, Spodoptera exigua. Biosci. Biotechnol. Biochem. 2009, 73, 2077–2084. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.-K.; Ullah, I.; Lim, J.-H.; Lee, I.-J.; Kim, J.-G.; Shin, J.-H. Physiological and molecular characterization of a newly identified entomopathogenic bacteria, Photorhabdus temperata M1021. J. Microbiol. Biotechnol. 2012, 22, 1605–1612. [Google Scholar] [CrossRef]
- Salvadori, J.D.M.; Defferrari, M.S.; Ligabue-Braun, R.; Lau, E.Y.; Salvadori, J.R.; Carlini, C.R. Characterization of entomopathogenic nematodes and symbiotic bacteria active against Spodoptera frugiperda (Lepidoptera: Noctuidae) and contribution of bacterial urease to the insecticidal effect. Biol. Control 2012, 63, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.S.; Kim, Y. Up-regulation of circulating hemocyte population in response to bacterial challenge is mediated by octopamine and 5-hydroxytryptamine via Rac1 signal in Spodoptera exigua. J. Insect Physiol. 2010, 56, 559–566. [Google Scholar] [CrossRef]
- Seo, S.; Lee, S.; Hong, Y.; Kim, Y. Phospholipase A2 inhibitors synthesized by two entomopathogenic bacteria, Xenorhabdus nematophila and Photorhabdus temperata subsp. temperata. Appl. Environ. Microbiol. 2012, 78, 3816–3823. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ke, L.; Xue, C.; Luo, W.; Chen, Q. Inhibitory Kinetics of p-Substituted Benzaldehydes on Polyphenol Oxidase from the Fifth Instar of Pieris Rapae L. Tsinghua Sci. Technol. 2007, 12, 400–404. [Google Scholar] [CrossRef]
- Sadekuzzaman, M.; Gautam, N.; Kim, Y. A novel calcium-independent phospholipase A2 and its physiological roles in development and immunity of a lepidopteran insect, Spodoptera exigua. Dev. Comp. Immunol. 2017, 77, 210–220. [Google Scholar] [CrossRef]
- Hwang, J.; Park, Y.; Kim, Y.; Hwang, J.; Lee, D. An entomopathogenic bacterium, Xenorhabdus nematophila, suppresses expression of antimicrobial peptides controlled by Toll and Imd pathways by blocking eicosanoid biosynthesis. Arch. Insect Biochem. Physiol. 2013, 83, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Keum, S.; Hasan, A.; Kim, H.; Jung, Y.; Lee, D.; Kim, Y. Identification of an entomopathogenic bacterium, Xenorhabdus ehlersii KSY, from Steinernema longicaudum GNUS101 and its immunosuppressive activity against insect host by inhibiting eicosanoid biosynthesis. J. Invertebr. Pathol. 2018, 159, 6–17. [Google Scholar] [CrossRef]
- Ahmed, S.; Kim, Y. Differential immunosuppression by inhibiting PLA2 affects virulence of Xenorhabdus hominickii and Photorhabdus temperata temperata. J. Invertebr. Pathol. 2018, 157, 136–146. [Google Scholar] [CrossRef]
- Ullah, I.; Khan, A.L.; Ali, L.; Khan, A.R.; Waqas, M.; Hussain, J.; Lee, I.-J.; Shin, J.-H. Benzaldehyde as an insecticidal, antimicrobial, and antioxidant compound produced by Photorhabdus temperata M1021. J. Microbiol. 2015, 53, 127–133. [Google Scholar] [CrossRef]
- Shrestha, S.; Kim, Y. Activation of immune-associated phospholipase A2 is functionally linked to Toll/Imd signal pathways in the red flour beetle, Tribolium castaneum. Dev. Comp. Immunol. 2010, 34, 530–537. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanda, N.B.; Hou, Y. The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae. Pathogens 2023, 12, 506. https://doi.org/10.3390/pathogens12040506
Sanda NB, Hou Y. The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae. Pathogens. 2023; 12(4):506. https://doi.org/10.3390/pathogens12040506
Chicago/Turabian StyleSanda, Nafiu Bala, and Youming Hou. 2023. "The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae" Pathogens 12, no. 4: 506. https://doi.org/10.3390/pathogens12040506
APA StyleSanda, N. B., & Hou, Y. (2023). The Symbiotic Bacteria—Xenorhabdus nematophila All and Photorhabdus luminescens H06 Strongly Affected the Phenoloxidase Activation of Nipa Palm Hispid, Octodonta nipae (Coleoptera: Chrysomelidae) Larvae. Pathogens, 12(4), 506. https://doi.org/10.3390/pathogens12040506