
Human Papilloma Virus: An Unraveled Enigma of Universal Burden of Malignancies



Abstract
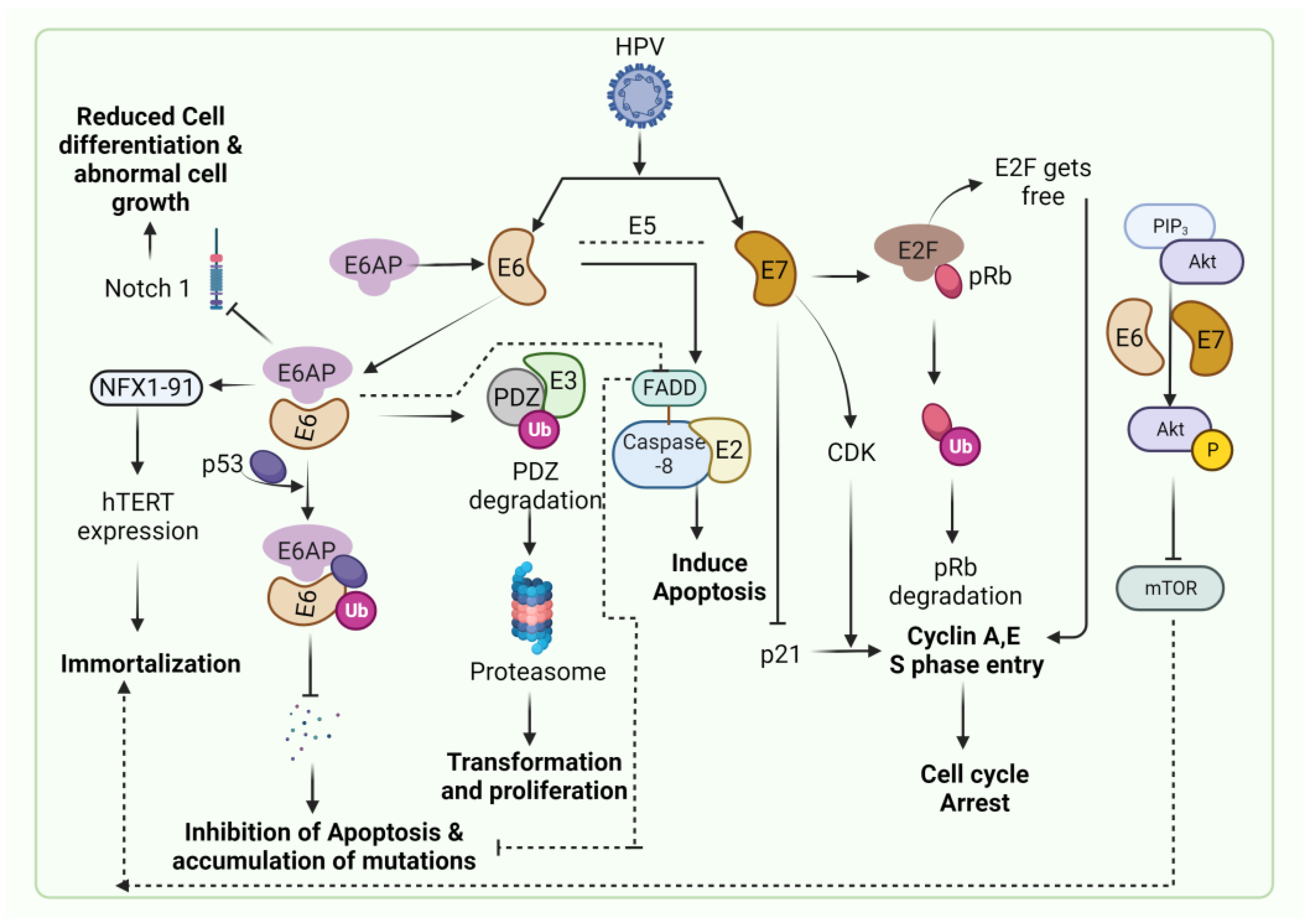
:1. Introduction
2. Burden of HPV Infection
3. HPV and Associated Malignancies
4. Cervical Cancer
5. Oropharyngeal Cancer
6. Vulvar and Vaginal Cancer
7. Anal Cancer
8. Colorectal Cancer
9. Other Cancer Types Associated with HPV
10. Vaccine Status

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine | Type | Target Strains | Manufacturer | Licensed Year | Target Population |
|---|---|---|---|---|---|
| Cervarix® | Bivalent | 16/18 | Glaxo SmihtKline | 2007 | Females & Males |
| Gardasil® | Quadrivalent | 6/11/16/18 | Merck | 2006 | Females & Males |
| Gardasil® 9 | Nonavalent | 6/11/16/18/31/33/45/52/58 | Merck | 2014 | Females & Males |
| Cecolin® | Bivalent | 16/18 | Xiamen Innovax Biotech | 2020 | Females |
| Cervavac® | Quadrivalent | 6/11/16/18 | Serum Institute of India | 2022 | Females & Males |
| Walrinvax® | Bivalent | 16/18 | Walvax/Zerun | 2022 | Females |
11. Alternate Therapies and Novel Therapeutics
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global burden of cancers attributable to infections in 2008: A review and synthetic analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.; de Martel, C.; Lacey, C.J.; Soerjomataram, I.; Lortet-Tieulent, J.; Bruni, L.; Vignat, J.; Ferlay, J.; Bray, F.; Plummer, M.; et al. Global Burden of Human Papillomavirus and Related Diseases. Vaccine 2012, 30, F12–F23. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, T.R.; Graybill, W.S.; Pierce, J.Y. Morbidity and mortality of vulvar and vaginal cancers: Impact of 2-, 4-, and 9-valent HPV vaccines. Hum. Vaccines Immunother. 2016, 12, 1352–1356. [Google Scholar] [CrossRef] [Green Version]
- Asiaf, A.; Ahmad, S.T.; Mohammad, S.O.; Zargar, M.A. Review of the current knowledge on the epidemiology, pathogenesis, and prevention of human papillomavirus infection. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. ECP 2014, 23, 206–224. [Google Scholar] [CrossRef]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- NCDIR-NCRP ICMR. National Cancer Registry Programme: Three-Year Report of Population-Based Cancer Registries 2012–2014; NCDIR-NCRP (ICMR): Bengaluru, India, 2016. [Google Scholar]
- Sankaranarayanan, R.; Bhatla, N.; Basu, P. Current global status & impact of human papillomavirus vaccination: Implications for India. Indian J. Med. Res. 2016, 144, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Jit, M.; Brisson, M.; Portnoy, A.; Hutubessy, R. Cost-effectiveness of female human papillomavirus vaccination in 179 countries: A PRIME modelling study. Lancet Glob. Health 2014, 2, e406–e414. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, R.; Hariprasad, R.; Rajaraman, P.; Mahajan, V.; Grover, R.; Kaur, P.; Swaminathan, S. Stemming the Wave of Cervical Cancer: Human Papillomavirus Vaccine Introduction in India. J. Glob. Oncol. 2018, 4, JGO.17.00030. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.; Andrews, K.S.; Brooks, D.; Fedewa, S.A.; Manassaram-Baptiste, D.; Saslow, D.; Brawley, O.W.; Wender, R.C. Cancer screening in the United States, 2017: A review of current American Cancer Society guidelines and current issues in cancer screening. CA Cancer J. Clin. 2017, 67, 100–121. [Google Scholar] [CrossRef]
- American Cancer Society American Cancer Society Guidelines for the Early Detection of Cancer 2022. Available online: https://www.cancer.org/healthy/find-cancer-early/american-cancer-society-guidelines-for-the-early-detection-of-cancer.html (accessed on 11 February 2023).
- Sharma, S.J.; Wagner, S.; Reder, H.S.F.; Kroll, T.; Wuerdemann, N.; Klussmann, J.P.; Wittekindt, C. The 8th edition AJCC/UICC TNM staging for p16-positive oropharyngeal carcinoma: Is there space for improvement? Eur. Arch. Oto-Rhino-Laryngol. 2018, 275, 3087–3091. [Google Scholar] [CrossRef]
- Westra, W.H. Detection of human papillomavirus (HPV) in clinical samples: Evolving methods and strategies for the accurate determination of HPV status of head and neck carcinomas. Oral Oncol. 2014, 50, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, I.; Vincent-Salomon, A.; Validire, P.; Zafrani, B.; de la Rochefordière, A.; Clough, K.; Favre, M.; Pouillart, P.; Sastre-Garau, X. Human papillomavirus genotype as a major determinant of the course of cervical cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1998, 16, 2613–2619. [Google Scholar] [CrossRef] [PubMed]
- Kombe Kombe, A.J.; Li, B.; Zahid, A.; Mengist, H.M.; Bounda, G.-A.; Zhou, Y.; Jin, T. Epidemiology and Burden of Human Papillomavirus and Related Diseases, Molecular Pathogenesis, and Vaccine Evaluation. Front. Public Health 2021, 8, 552028. [Google Scholar] [CrossRef] [PubMed]
- Bruni, L.; Saura-Lázaro, A.; Montoliu, A.; Brotons, M.; Alemany, L.; Diallo, M.S.; Afsar, O.Z.; LaMontagne, D.S.; Mosina, L.; Contreras, M.; et al. HPV vaccination introduction worldwide and WHO and UNICEF estimates of national HPV immunization coverage 2010–2019. Prev. Med. 2021, 144, 106399. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Diaz, M.; Castellsagué, X.; Clifford, G.; Bruni, L.; Muñoz, N.; Bosch, F.X. Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology: A meta-analysis. Lancet Infect. Dis. 2007, 7, 453–459. [Google Scholar] [CrossRef]
- Clifford, G.M.; Gallus, S.; Herrero, R.; Muñoz, N.; Snijders, P.J.F.; Vaccarella, S.; Anh, P.T.H.; Ferreccio, C.; Hieu, N.T.; Matos, E.; et al. Worldwide distribution of human papillomavirus types in cytologically normal women in the International Agency for Research on Cancer HPV prevalence surveys: A pooled analysis. Lancet 2005, 366, 991–998. [Google Scholar] [CrossRef]
- Bruni, L.; Diaz, M.; Castellsagué, X.; Ferrer, E.; Bosch, F.X.; de Sanjosé, S. Cervical human papillomavirus prevalence in 5 continents: Meta-analysis of 1 million women with normal cytological findings. J. Infect. Dis. 2010, 202, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Senapati, R.; Nayak, B.; Kar, S.K.; Dwibedi, B. HPV Genotypes distribution in Indian women with and without cervical carcinoma: Implication for HPV vaccination program in Odisha, Eastern India. BMC Infect. Dis. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, R.; Das, S.S.; Biswal, S.S.; Nath, A.; Das, D.; Basu, A.; Malik, S.; Kumar, L.; Kar, S.; Singh, S.K.; et al. Mechanistic role of HPV-associated early proteins in cervical cancer: Molecular pathways and targeted therapeutic strategies. Crit. Rev. Oncol. Hematol. 2022, 174, 103675. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Liu, J.; Masterson, L.; Fenton, T.R. HPV-associated oropharyngeal cancer: Epidemiology, molecular biology and clinical management. Nat. Rev. Clin. Oncol. 2022, 19, 306–327. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ramírez, I.; Carrillo-García, A.; Contreras-Paredes, A.; Ortiz-Sánchez, E.; Cruz-Gregorio, A.; Lizano, M. Regulation of Cellular Metabolism by High-Risk Human Papillomaviruses. Int. J. Mol. Sci. 2018, 19, 1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuninghame, S.; Jackson, R.; Zehbe, I. Hypoxia-inducible factor 1 and its role in viral carcinogenesis. Virology 2014, 456–457, 370–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narisawa-Saito, M.; Kiyono, T. Basic mechanisms of high-risk human papillomavirus-induced carcinogenesis: Roles of E6 and E7 proteins. Cancer Sci. 2007, 98, 1505–1511. [Google Scholar] [CrossRef]
- Vieira, G.V.; Somera Dos Santos, F.; Lepique, A.P.; da Fonseca, C.K.; Innocentini, L.M.A.R.; Braz-Silva, P.H.; Quintana, S.M.; Sales, K.U. Proteases and HPV-Induced Carcinogenesis. Cancers 2022, 14, 3038. [Google Scholar] [CrossRef] [PubMed]
- Otter, S.; Whitaker, S.; Chatterjee, J.; Stewart, A. The Human Papillomavirus as a Common Pathogen in Oropharyngeal, Anal and Cervical Cancers. Clin. Oncol. 2019, 31, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.; Viveros-Carreño, D.; Hoegl, J.; Ávila, M.; Pareja, R. Human papillomavirus-independent cervical cancer. Int. J. Gynecol. Cancer 2022, 32, 1–7. [Google Scholar] [CrossRef]
- Dufour, X.; Beby-Defaux, A.; Agius, G.; Lacau St Guily, J. HPV and head and neck cancer. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2012, 129, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Sartor, M.A.; Dolinoy, D.C.; Jones, T.R.; Colacino, J.A.; Prince, M.E.P.; Carey, T.E.; Rozek, L.S. Genome-wide methylation and expression differences in HPV(+) and HPV(-) squamous cell carcinoma cell lines are consistent with divergent mechanisms of carcinogenesis. Epigenetics 2011, 6, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.A.; Werth, A.J.; Sharaf, R.; Montesion, M.; Sokol, E.S.; Pavlick, D.C.; McLaughlin-Drubin, M.; Erlich, R.; Toma, H.; Williams, K.J.; et al. Vulvar Squamous Cell Carcinoma: Comprehensive Genomic Profiling of HPV+ Versus HPV- Forms Reveals Distinct Sets of Potentially Actionable Molecular Targets. JCO Precis. Oncol. 2020, 4, PO.19.00406. [Google Scholar] [CrossRef]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M.; International Agency for Research on Cancer Multicenter Cervical Cancer Study Group. Epidemiologic classification of human papillomavirus types associated with cervical cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghebre, R.; Berry-Lawhorn, J.M.; D’Souza, G. State of the Science: Screening, Surveillance, and Epidemiology of HPV-Related Malignancies. Am. Soc. Clin. Oncol. Educ. Book 2021, 41, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Fontham, E.T.H.; Wolf, A.M.D.; Church, T.R.; Etzioni, R.; Flowers, C.R.; Herzig, A.; Guerra, C.E.; Oeffinger, K.C.; Shih, Y.-C.T.; Walter, L.C.; et al. Cervical cancer screening for individuals at average risk: 2020 guideline update from the American Cancer Society. CA Cancer J. Clin. 2020, 70, 321–346. [Google Scholar] [CrossRef]
- Obermair, H.M.; Bennett, K.F.; Brotherton, J.M.L.; Smith, M.A.; McCaffery, K.J.; Dodd, R.H. Australian National Cervical Screening Program renewal: Attitudes and experiences of general practitioners, and obstetricians and gynaecologists. Aust. N. Z. J. Obstet. Gynaecol. 2021, 61, 416–423. [Google Scholar] [CrossRef]
- Brown, R.F.; Muller, T.R.; Olsen, A. Australian women’s cervical cancer screening attendance as a function of screening barriers and facilitators. Soc. Sci. Med. 2019, 220, 396–402. [Google Scholar] [CrossRef]
- Gu, C.; Chan, C.W.H.; Chow, K.M.; Yang, S.; Luo, Y.; Cheng, H.; Wang, H. Understanding the cervical screening behaviour of Chinese women: The role of health care system and health professions. Appl. Nurs. Res. ANR 2018, 39, 58–64. [Google Scholar] [CrossRef]
- Schiffman, M.; Wentzensen, N. A Suggested Approach to Simplify and Improve Cervical Screening in the United States. J. Low. Genit. Tract Dis. 2016, 20, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Karsa, L.; Arbyn, M.; De Vuyst, H.; Dillner, J.; Dillner, L.; Franceschi, S.; Patnick, J.; Ronco, G.; Segnan, N.; Suonio, E.; et al. European guidelines for quality assurance in cervical cancer screening. Summary of the supplements on HPV screening and vaccination. Papillomavirus Res. 2015, 1, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Ronco, G.; Giorgi-Rossi, P.; Carozzi, F.; Confortini, M.; Dalla Palma, P.; Del Mistro, A.; Ghiringhello, B.; Girlando, S.; Gillio-Tos, A.; De Marco, L.; et al. Efficacy of human papillomavirus testing for the detection of invasive cervical cancers and cervical intraepithelial neoplasia: A randomised controlled trial. Lancet Oncol. 2010, 11, 249–257. [Google Scholar] [CrossRef]
- Ghebre, R.G.; Grover, S.; Xu, M.J.; Chuang, L.T.; Simonds, H. Cervical cancer control in HIV-infected women: Past, present and future. Gynecol. Oncol. Rep. 2017, 21, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Roura, E.; Castellsagué, X.; Pawlita, M.; Travier, N.; Waterboer, T.; Margall, N.; Bosch, F.X.; de Sanjosé, S.; Dillner, J.; Gram, I.T.; et al. Smoking as a major risk factor for cervical cancer and pre-cancer: Results from the EPIC cohort. Int. J. Cancer 2014, 135, 453–466. [Google Scholar] [CrossRef]
- Stelzle, D.; Tanaka, L.F.; Lee, K.K.; Ibrahim Khalil, A.; Baussano, I.; Shah, A.S.V.; McAllister, D.A.; Gottlieb, S.L.; Klug, S.J.; Winkler, A.S.; et al. Estimates of the global burden of cervical cancer associated with HIV. Lancet Glob. Health 2021, 9, e161–e169. [Google Scholar] [CrossRef]
- Yuan, Y.; Cai, X.; Shen, F.; Ma, F. HPV post-infection microenvironment and cervical cancer. Cancer Lett. 2021, 497, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.; Franceschi, S.; Bosetti, C.; Moreno, V.; Herrero, R.; Smith, J.S.; Shah, K.V.; Meijer, C.J.L.M.; Bosch, F.X.; International Agency for Research on Cancer. Multicentric Cervical Cancer Study Group Role of parity and human papillomavirus in cervical cancer: The IARC multicentric case-control study. Lancet 2002, 359, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, H.; Zhang, L.; Qiao, Y. Cervical cancer: Epidemiology, risk factors and screening. Chin. J. Cancer Res. Chung-Kuo Yen Cheng Yen Chiu 2020, 32, 720–728. [Google Scholar] [CrossRef]
- International Collaboration of Epidemiological Studies of Cervical Cancer; Appleby, P.; Beral, V.; de González, A.B.; Colin, D.; Franceschi, S.; Goodhill, A.; Green, J.; Peto, J.; Plummer, M.; et al. Cervical cancer and hormonal contraceptives: Collaborative reanalysis of individual data for 16,573 women with cervical cancer and 35,509 women without cervical cancer from 24 epidemiological studies. Lancet 2007, 370, 1609–1621. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Zhang, Y. Involvement of Human Papillomaviruses in Cervical Cancer. Front. Microbiol. 2018, 9, 2896. [Google Scholar] [CrossRef]
- Nakamura, M.; Obata, T.; Daikoku, T.; Fujiwara, H. The Association and Significance of p53 in Gynecologic Cancers: The Potential of Targeted Therapy. Int. J. Mol. Sci. 2019, 20, 5482. [Google Scholar] [CrossRef] [Green Version]
- Yugawa, T.; Handa, K.; Narisawa-Saito, M.; Ohno, S.; Fujita, M.; Kiyono, T. Regulation of Notch1 gene expression by p53 in epithelial cells. Mol. Cell. Biol. 2007, 27, 3732–3742. [Google Scholar] [CrossRef] [Green Version]
- Narisawa-Saito, M.; Handa, K.; Yugawa, T.; Ohno, S.; Fujita, M.; Kiyono, T. HPV16 E6-mediated stabilization of ErbB2 in neoplastic transformation of human cervical keratinocytes. Oncogene 2007, 26, 2988–2996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, V.M.B.; Shalini, J.V.; Lekha, T.; Devaraj, S.N.; Devaraj, H. Co-overexpression of p53 and bcl-2 proteins in HPV-induced squamous cell carcinoma of the uterine cervix. Gynecol. Oncol. 2003, 91, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Mitildzans, A.; Arechvo, A.; Rezeberga, D.; Isajevs, S. Expression of p63, p53 and Ki-67 in Patients with Cervical Intraepithelial Neoplasia. Turk Patoloji Derg. 2017, 33, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Ntuli, L.; Mtshali, A.; Mzobe, G.; Liebenberg, L.J.; Ngcapu, S. Role of Immunity and Vaginal Microbiome in Clearance and Persistence of Human Papillomavirus Infection. Front. Cell. Infect. Microbiol. 2022, 12, 927131. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef] [Green Version]
- Cascardi, E.; Cazzato, G.; Daniele, A.; Silvestris, E.; Cormio, G.; Di Vagno, G.; Malvasi, A.; Loizzi, V.; Scacco, S.; Pinto, V.; et al. Association between Cervical Microbiota and HPV: Could This Be the Key to Complete Cervical Cancer Eradication? Biology 2022, 11, 1114. [Google Scholar] [CrossRef]
- Mitra, A.; MacIntyre, D.A.; Lee, Y.S.; Smith, A.; Marchesi, J.R.; Lehne, B.; Bhatia, R.; Lyons, D.; Paraskevaidis, E.; Li, J.V.; et al. Cervical intraepithelial neoplasia disease progression is associated with increased vaginal microbiome diversity. Sci. Rep. 2015, 5, 16865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, A.; MacIntyre, D.A.; Marchesi, J.R.; Lee, Y.S.; Bennett, P.R.; Kyrgiou, M. The vaginal microbiota, human papillomavirus infection and cervical intraepithelial neoplasia: What do we know and where are we going next? Microbiome 2016, 4, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Norenhag, J.; Hu, Y.O.O.; Brusselaers, N.; Fransson, E.; Ährlund-Richter, A.; Guðnadóttir, U.; Angelidou, P.; Zha, Y.; Hamsten, M.; et al. Vaginal microbiota and human papillomavirus infection among young Swedish women. Npj Biofilms Microbiomes 2020, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Santella, B.; Schettino, M.T.; Franci, G.; De Franciscis, P.; Colacurci, N.; Schiattarella, A.; Galdiero, M. Microbiota and HPV: The role of viral infection on vaginal microbiota. J. Med. Virol. 2022, 94, 4478–4484. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Lee, S.; Lee, H.; Song, Y.-M.; Lee, K.; Han, M.J.; Sung, J.; Ko, G. Association of the vaginal microbiota with human papillomavirus infection in a Korean twin cohort. PLoS ONE 2013, 8, e63514. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, Z.; Chen, T. Role of Vaginal Microbiota Dysbiosis in Gynecological Diseases and the Potential Interventions. Front. Microbiol. 2021, 12, 643422. [Google Scholar] [CrossRef]
- Mitra, T.; Elangovan, S. Cervical cancer development, chemoresistance, and therapy: A snapshot of involvement of microRNA. Mol. Cell. Biochem. 2021, 476, 4363–4385. [Google Scholar] [CrossRef]
- Banno, K.; Iida, M.; Yanokura, M.; Kisu, I.; Iwata, T.; Tominaga, E.; Tanaka, K.; Aoki, D. MicroRNA in cervical cancer: OncomiRs and tumor suppressor miRs in diagnosis and treatment. Sci. World J. 2014, 2014, 178075. [Google Scholar] [CrossRef]
- Yao, Q.; Xu, H.; Zhang, Q.-Q.; Zhou, H.; Qu, L.-H. MicroRNA-21 promotes cell proliferation and down-regulates the expression of programmed cell death 4 (PDCD4) in HeLa cervical carcinoma cells. Biochem. Biophys. Res. Commun. 2009, 388, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Hemmat, N.; Bannazadeh Baghi, H. Association of human papillomavirus infection and inflammation in cervical cancer. Pathog. Dis. 2019, 77, ftz048. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Zhen, Y.; Chen, Y.; Zou, L.; Zhang, Y.; Hu, F.; Feng, J.; Shen, J.; Wei, B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-κB/COX-2-caspase-1 pathway in HeLa cervical cancer cells. Int. J. Oncol. 2014, 45, 1929–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampias, T.; Sasaki, C.; Psyrri, A. Molecular mechanisms of HPV induced carcinogenesis in head and neck. Oral Oncol. 2014, 50, 356–363. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human papillomavirus and rising oropharyngeal cancer incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef]
- Rampias, T.; Sasaki, C.; Weinberger, P.; Psyrri, A. E6 and e7 gene silencing and transformed phenotype of human papillomavirus 16-positive oropharyngeal cancer cells. J. Natl. Cancer Inst. 2009, 101, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Saada-Bouzid, E.; Peyrade, F.; Guigay, J. Molecular genetics of head and neck squamous cell carcinoma. Curr. Opin. Oncol. 2019, 31, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, A.K.; Sojourner, E.J.; Vitzthum, L.K.; Zakeri, K.; Shen, H.; Nguyen, C.; Murphy, J.D.; Califano, J.A.; Cohen, E.E.W.; Mell, L.K. Prognostic Role of p16 in Nonoropharyngeal Head and Neck Cancer. J. Natl. Cancer Inst. 2018, 110, 1393–1399. [Google Scholar] [CrossRef]
- Dayyani, F.; Etzel, C.J.; Liu, M.; Ho, C.-H.; Lippman, S.M.; Tsao, A.S. Meta-analysis of the impact of human papillomavirus (HPV) on cancer risk and overall survival in head and neck squamous cell carcinomas (HNSCC). Head Neck Oncol. 2010, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Lechner, M.; Jones, O.S.; Breeze, C.E.; Gilson, R. Gender-neutral HPV vaccination in the UK, rising male oropharyngeal cancer rates, and lack of HPV awareness. Lancet Infect. Dis. 2019, 19, 131–132. [Google Scholar] [CrossRef] [Green Version]
- Faraji, F.; Rettig, E.M.; Tsai, H.-L.; El Asmar, M.; Fung, N.; Eisele, D.W.; Fakhry, C. The prevalence of human papillomavirus in oropharyngeal cancer is increasing regardless of sex or race, and the influence of sex and race on survival is modified by human papillomavirus tumor status. Cancer 2019, 125, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Carlander, A.F.; Jakobsen, K.K.; Bendtsen, S.K.; Garset-Zamani, M.; Lynggaard, C.D.; Jensen, J.S.; Grønhøj, C.; Buchwald, C. von A Contemporary Systematic Review on Repartition of HPV-Positivity in Oropharyngeal Cancer Worldwide. Viruses 2021, 13, 1326. [Google Scholar] [CrossRef]
- Mehanna, H.; Evans, M.; Beasley, M.; Chatterjee, S.; Dilkes, M.; Homer, J.; O’Hara, J.; Robinson, M.; Shaw, R.; Sloan, P. Oropharyngeal cancer: United Kingdom National Multidisciplinary Guidelines. J. Laryngol. Otol. 2016, 130, S90–S96. [Google Scholar] [CrossRef]
- Gage, K.L.; Thomas, K.; Jeong, D.; Stallworth, D.G.; Arrington, J.A. Multimodal Imaging of Head and Neck Squamous Cell Carcinoma. Cancer Control. J. Moffitt Cancer Cent. 2017, 24, 172–179. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Zhang, Z. Prevalence of human papillomavirus and its prognostic value in vulvar cancer: A systematic review and meta-analysis. PLoS ONE 2018, 13, e0204162. [Google Scholar] [CrossRef]
- Halec, G.; Alemany, L.; Quiros, B.; Clavero, O.; Höfler, D.; Alejo, M.; Quint, W.; Pawlita, M.; Bosch, F.X.; de Sanjose, S. Biological relevance of human papillomaviruses in vulvar cancer. Mod. Pathol. 2017, 30, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortekaas, K.E.; Bastiaannet, E.; van Doorn, H.C.; de Vos van Steenwijk, P.J.; Ewing-Graham, P.C.; Creutzberg, C.L.; Akdeniz, K.; Nooij, L.S.; van der Burg, S.H.; Bosse, T.; et al. Vulvar cancer subclassification by HPV and p53 status results in three clinically distinct subtypes. Gynecol. Oncol. 2020, 159, 649–656. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, H.; Clifford, G.M.; Nascimento, M.C.; Madeleine, M.M.; Franceschi, S. Prevalence and type distribution of human papillomavirus in carcinoma and intraepithelial neoplasia of the vulva, vagina and anus: A meta-analysis. Int. J. Cancer 2009, 124, 1626–1636. [Google Scholar] [CrossRef]
- Hay, C.M.; Lachance, J.A.; Lucas, F.L.; Smith, K.A.; Jones, M.A. Biomarkers p16, Human Papillomavirus and p53 Predict Recurrence and Survival in Early Stage Squamous Cell Carcinoma of the Vulva. J. Low. Genit. Tract Dis. 2016, 20, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.J.; Burger, R.A.; Lin, F.; Parham, G.; Vasilev, S.A.; Wilczynski, S.P. Prognostic significance of human papillomavirus DNA in vulvar carcinoma. Obstet. Gynecol. 1995, 85, 709–715. [Google Scholar] [CrossRef]
- Wakeham, K.; Kavanagh, K.; Cuschieri, K.; Millan, D.; Pollock, K.G.; Bell, S.; Burton, K.; Reed, N.S.; Graham, S.V. HPV status and favourable outcome in vulvar squamous cancer. Int. J. Cancer 2017, 140, 1134–1146. [Google Scholar] [CrossRef]
- ACOG, C.O. ACOG Committee Opinion No. 509: Management of vulvar intraepithelial neoplasia. Obstet. Gynecol. 2011, 118, 1192–1194. [Google Scholar] [CrossRef]
- Judson, P.L.; Habermann, E.B.; Baxter, N.N.; Durham, S.B.; Virnig, B.A. Trends in the incidence of invasive and in situ vulvar carcinoma. Obstet. Gynecol. 2006, 107, 1018–1022. [Google Scholar] [CrossRef]
- Adams, T.S.; Rogers, L.J.; Cuello, M.A. Cancer of the vagina: 2021 update. Int. J. Gynaecol. Obstet. 2021, 155 (Suppl. S1), 19–27. [Google Scholar] [CrossRef]
- Adhikari, P.; Vietje, P.; Mount, S. Premalignant and malignant lesions of the vagina. Diagn. Histopathol. 2017, 23, 28–34. [Google Scholar] [CrossRef]
- Hellman, K.; Silfverswärd, C.; Nilsson, B.; Hellström, A.C.; Frankendal, B.; Pettersson, F. Primary carcinoma of the vagina: Factors influencing the age at diagnosis. The Radiumhemmet series 1956-96. Int. J. Gynecol. Cancer 2004, 14, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Hampl, M.; Sarajuuri, H.; Wentzensen, N.; Bender, H.G.; Kueppers, V. Effect of human papillomavirus vaccines on vulvar, vaginal, and anal intraepithelial lesions and vulvar cancer. Obstet. Gynecol. 2006, 108, 1361–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamos, C.; Mihaljevic, C.; Aulmann, S.; Bruckner, T.; Domschke, C.; Wallwiener, M.; Paringer, C.; Fluhr, H.; Schott, S.; Dinkic, C.; et al. Detection of Human Papillomavirus Infection in Patients with Vaginal Intraepithelial Neoplasia. PLoS ONE 2016, 11, e0167386. [Google Scholar] [CrossRef]
- Serrano, B.; de Sanjosé, S.; Tous, S.; Quiros, B.; Muñoz, N.; Bosch, X.; Alemany, L. Human papillomavirus genotype attribution for HPVs 6, 11, 16, 18, 31, 33, 45, 52 and 58 in female anogenital lesions. Eur. J. Cancer 2015, 51, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, H.K.; Rasmussen, C.L.; Sand, F.L.; Albieri, V.; Norrild, B.; Verdoodt, F.; Kjaer, S.K. Human papillomavirus and p16 in squamous cell carcinoma and intraepithelial neoplasia of the vagina. Int. J. Cancer 2019, 145, 78–86. [Google Scholar] [CrossRef]
- Rasmussen, C.L.; Bertoli, H.K.; Sand, F.L.; Kjaer, A.K.; Thomsen, L.T.; Kjaer, S.K. The prognostic significance of HPV, p16, and p53 protein expression in vaginal cancer: A systematic review. Acta Obstet. Gynecol. Scand. 2021, 100, 2144–2156. [Google Scholar] [CrossRef]
- Palma Dos Reis, C.; Correia, L.; Jorge, A.F.; Félix, A. Primary vaginal cancer associated with HPV 68: A case report of an unusual presentation. Int. J. Gynaecol. Obstet. 2021, 152, 262–263. [Google Scholar] [CrossRef]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA Cancer J. Clin. 2012, 62, 10–29. [Google Scholar] [CrossRef] [Green Version]
- Arens, Y.; Gaisa, M.; Goldstone, S.E.; Liu, Y.; Wisnivesky, J.; Sigel, C.S.; Swartz, T.H.; Sigel, K. Risk of Invasive Anal Cancer in HIV-Infected Patients With High-Grade Anal Dysplasia: A Population-Based Cohort Study. Dis. Colon Rectum 2019, 62, 934–940. [Google Scholar] [CrossRef]
- Clifford, G.M.; Georges, D.; Shiels, M.S.; Engels, E.A.; Albuquerque, A.; Poynten, I.M.; de Pokomandy, A.; Easson, A.M.; Stier, E.A. A meta-analysis of anal cancer incidence by risk group: Toward a unified anal cancer risk scale. Int. J. Cancer 2021, 148, 38–47. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Serrano, B.; Tous, S.; Alejo, M.; Lloveras, B.; Quirós, B.; Clavero, O.; Vidal, A.; Ferrándiz-Pulido, C.; Pavón, M.Á.; et al. Burden of Human Papillomavirus (HPV)-Related Cancers Attributable to HPVs 6/11/16/18/31/33/45/52 and 58. JNCI Cancer Spectr. 2018, 2, pky045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Sharma, M.; Tan, N.; Barnabas, R.V. HIV-positive women have higher risk of human papilloma virus infection, precancerous lesions, and cervical cancer. Aids 2018, 32, 795–808. [Google Scholar] [CrossRef]
- Wang, C.-C.J.; Sparano, J.; Palefsky, J.M. Human Immunodeficiency Virus/AIDS, Human Papillomavirus, and Anal Cancer. Surg. Oncol. Clin. N. Am. 2017, 26, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Hernando, L.; Oliver-Pérez, R.; Bravo-Violeta, V.; Olloqui, A.; Parte-Izquierdo, B.; Almansa-González, C.; Bermejo-Martinez, R.; Bolivar-De Miguel, A.B.; Diez, A.; Seoane-Ruiz, J.M.; et al. Prevalence of Anal High-Risk Human Papilloma Virus Infection and Abnormal Anal Cytology among Women Living with HIV. J. Pers. Med. 2022, 12, 1778. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, W.; Paszat, L.; Kupets, R.; Wilton, A.; Tinmouth, J. Presumed previous human papillomavirus (HPV) related gynecological cancer in women diagnosed with anal cancer in the province of Ontario. Gynecol. Oncol. 2009, 114, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.M.; Paulus, J.K.; Shapter, A.P.; Baxter, N.N.; Roberts, P.L.; Ricciardi, R. Risk of anal cancer in a cohort with human papillomavirus-related gynecologic neoplasm. Obstet. Gynecol. 2011, 117, 643–649. [Google Scholar] [CrossRef]
- Sehnal, B.; Dusek, L.; Cibula, D.; Zima, T.; Halaska, M.; Driak, D.; Slama, J. The relationship between the cervical and anal HPV infection in women with cervical intraepithelial neoplasia. J. Clin. Virol. 2014, 59, 18–23. [Google Scholar] [CrossRef]
- Gonzalez-Losa, M.D.R.; Puerto-Solís, M.; Ayora-Talavera, G.; Gómez-Carvallo, J.; Euán-López, A.; Cisneros-Cutz, J.I.; Rosado-López, A.; Echeverría Salazar, J.; Conde-Ferráez, L. Prevalence of anal infection due to high-risk human papillomavirus and analysis of E2 gene integrity among women with cervical abnormalities. Enferm. Infecc. Microbiol. Clin. Engl. Ed. 2018, 36, 209–213. [Google Scholar] [CrossRef]
- Yu, H.-G.; Shun, L.-B.; Luo, H.-S.; Huang, H.; Yu, B.-P.; Yu, J.-P.; Meier, J.J.; Schrader, H.; Bastian, A.; Schmitz, F.; et al. Deletion of the FHIT gene in human colorectal cancer is independent of high-risk HPV infection. Int. J. Color. Dis. 2002, 17, 396–401. [Google Scholar] [CrossRef]
- Chen, A.C.-H.; Keleher, A.; Kedda, M.-A.; Spurdle, A.B.; McMillan, N.A.J.; Antonsson, A. Human papillomavirus DNA detected in peripheral blood samples from healthy Australian male blood donors. J. Med. Virol. 2009, 81, 1792–1796. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-H.; Huang, C.-C.; Yeh, K.-T.; Chang, S.-H.; Chang, S.-W.; Sung, W.-W.; Cheng, Y.-W.; Lee, H. Human papilloma virus 16 E6 oncoprotein associated with p53 inactivation in colorectal cancer. World J. Gastroenterol. 2012, 18, 4051–4058. [Google Scholar] [CrossRef] [PubMed]
- Lorenzon, L.; Ferri, M.; Pilozzi, E.; Torrisi, M.R.; Ziparo, V.; French, D. Human papillomavirus and colorectal cancer: Evidences and pitfalls of published literature. Int. J. Color. Dis. 2011, 26, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Soto, Y.; Limia, C.M.; González, L.; Grá, B.; Hano, O.M.; Martínez, P.A.; Kourí, V. Molecular evidence of high-risk human papillomavirus infection in colorectal tumours from Cuban patients. Mem. Inst. Oswaldo Cruz 2016, 111, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodaghi, S.; Yamanegi, K.; Xiao, S.-Y.; Da Costa, M.; Palefsky, J.M.; Zheng, Z.-M. Colorectal papillomavirus infection in patients with colorectal cancer. Clin. Cancer Res. 2005, 11, 2862–2867. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.Y.; Sheu, L.F.; Meng, C.L.; Lee, W.H.; Lin, J.C. Detection of human papillomavirus DNA in colorectal carcinomas by polymerase chain reaction. Gut 1995, 37, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Pérez, L.O.; Abba, M.C.; Laguens, R.M.; Golijow, C.D. Analysis of adenocarcinoma of the colon and rectum: Detection of human papillomavirus (HPV) DNA by polymerase chain reaction. Color. Dis. 2005, 7, 492–495. [Google Scholar] [CrossRef]
- Buyru, N.; Tezol, A.; Dalay, N. Coexistence of K-ras mutations and HPV infection in colon cancer. BMC Cancer 2006, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.V.; Daniel, R.W.; Simons, J.W.; Vogelstein, B. Investigation of colon cancers for human papillomavirus genomic sequences by polymerase chain reaction. J. Surg. Oncol. 1992, 51, 5–7. [Google Scholar] [CrossRef]
- Strickler, H.D.; Schiffman, M.H.; Shah, K.V.; Rabkin, C.S.; Schiller, J.T.; Wacholder, S.; Clayman, B.; Viscidi, R.P. A survey of human papillomavirus 16 antibodies in patients with epithelial cancers. Eur. J. Cancer Prev. 1998, 7, 305–313. [Google Scholar] [CrossRef]
- Audeau, A.; Han, H.W.; Johnston, M.J.; Whitehead, M.W.; Frizelle, F.A. Does human papilloma virus have a role in squamous cell carcinoma of the colon and upper rectum? Eur. J. Surg. Oncol. 2002, 28, 657–660. [Google Scholar] [CrossRef]
- Gornick, M.C.; Castellsague, X.; Sanchez, G.; Giordano, T.J.; Vinco, M.; Greenson, J.K.; Capella, G.; Raskin, L.; Rennert, G.; Gruber, S.B.; et al. Human papillomavirus is not associated with colorectal cancer in a large international study. Cancer Causes Control. CCC 2010, 21, 737–743. [Google Scholar] [CrossRef]
- Baandrup, L.; Thomsen, L.T.; Olesen, T.B.; Andersen, K.K.; Norrild, B.; Kjaer, S.K. The prevalence of human papillomavirus in colorectal adenomas and adenocarcinomas: A systematic review and meta-analysis. Eur. J. Cancer 2014, 50, 1446–1461. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Mou, X.; Zhao, N.; Lin, J.; Teng, L.; Xiang, C. Prevalence of human papillomavirus in Chinese patients with colorectal cancer. Color. Dis. 2011, 13, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Burnett-Hartman, A.N.; Newcomb, P.A.; Potter, J.D. Infectious agents and colorectal cancer: A review of Helicobacter pylori, Streptococcus bovis, JC virus, and human papillomavirus. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2970–2979. [Google Scholar] [CrossRef] [Green Version]
- Bello, J.O.M.; Nieva, L.O.; Paredes, A.C.; Gonzalez, A.M.F.; Zavaleta, L.R.; Lizano, M. Regulation of the Wnt/β-Catenin Signaling Pathway by Human Papillomavirus E6 and E7 Oncoproteins. Viruses 2015, 7, 4734–4755. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-M.; Cheng, Y.-W.; Wang, Y.-C.; Wu, T.-C.; Chen, C.-Y.; Lee, H. Up-regulation of FOXM1 by E6 oncoprotein through the MZF1/NKX2-1 axis is required for human papillomavirus-associated tumorigenesis. Neoplasia 2014, 16, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.O.; Waga, S.; Harry, J.B.; Espling, E.; Stillman, B.; Galloway, D.A. Inhibition of CDK activity and PCNA-dependent DNA replication by p21 is blocked by interaction with the HPV-16 E7 oncoprotein. Genes Dev. 1997, 11, 2090–2100. [Google Scholar] [CrossRef] [Green Version]
- Bodily, J.; Laimins, L.A. Persistence of human papillomavirus infection: Keys to malignant progression. Trends Microbiol. 2011, 19, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Kessis, T.D.; Connolly, D.C.; Hedrick, L.; Cho, K.R. Expression of HPV16 E6 or E7 increases integration of foreign DNA. Oncogene 1996, 13, 427–431. [Google Scholar]
- Duensing, S.; Münger, K. The human papillomavirus type 16 E6 and E7 oncoproteins independently induce numerical and structural chromosome instability. Cancer Res. 2002, 62, 7075–7082. [Google Scholar]
- Lim, Y.P.; Lim, T.T.; Chan, Y.L.; Song, A.C.M.; Yeo, B.H.; Vojtesek, B.; Coomber, D.; Rajagopal, G.; Lane, D. The p53 knowledgebase: An integrated information resource for p53 research. Oncogene 2007, 26, 1517–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffner, M.; Werness, B.A.; Huibregtse, J.M.; Levine, A.J.; Howley, P.M. The E6 oncoprotein encoded by human papillomavirus types 16 and 18 promotes the degradation of p53. Cell 1990, 63, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Scheffner, M.; Huibregtse, J.M.; Vierstra, R.D.; Howley, P.M. The HPV-16 E6 and E6-AP complex functions as a ubiquitin-protein ligase in the ubiquitination of p53. Cell 1993, 75, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Werness, B.A.; Levine, A.J.; Howley, P.M. Association of human papillomavirus types 16 and 18 E6 proteins with p53. Science 1990, 248, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, L.; Allgayer, H. Viruses in colorectal cancer. Mol. Oncol. 2022, 16, 1423–1450. [Google Scholar] [CrossRef]
- Santos, N.; Bezerra Tocantins, P.; Bernardes Leão-Cordeiro, J.; Ataides, F.; Rosa Marques, L.O.; Cordeiro Silva, A.T. The human papillomavirus in colorectal cancer. J. Med. Sci. 2022, 42, 1–7. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Li, J.; Ding, J.; Zhai, K. Detection of Human Papillomavirus DNA in Patients with Breast Tumor in China. PLoS ONE 2015, 10, e0136050. [Google Scholar] [CrossRef] [Green Version]
- de Villiers, E.-M.; Sandstrom, R.E.; zur Hausen, H.; Buck, C.E. Presence of papillomavirus sequences in condylomatous lesions of the mamillae and in invasive carcinoma of the breast. Breast Cancer Res. 2004, 7, R1. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Chakraborty, B.; Panda, C.K. Human papilloma virus (HPV) profiles in breast cancer: Future management. Ann. Transl. Med. 2020, 8, 650. [Google Scholar] [CrossRef]
- Lawson, J.S.; Glenn, W.K.; Salyakina, D.; Clay, R.; Delprado, W.; Cheerala, B.; Tran, D.D.; Ngan, C.C.; Miyauchi, S.; Karim, M.; et al. Human Papilloma Virus Identification in Breast Cancer Patients with Previous Cervical Neoplasia. Front. Oncol. 2015, 5, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodaghi, S.; Wood, L.V.; Roby, G.; Ryder, C.; Steinberg, S.M.; Zheng, Z.-M. Could human papillomaviruses be spread through blood? J. Clin. Microbiol. 2005, 43, 5428–5434. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Goepfert, R.; Vela-Chávez, T.; Carrillo-García, A.; Lizano-Soberón, M.; Amador-Molina, A.; Oñate-Ocaña, L.F.; Hallmann, R.S.-R. High-risk human papillomavirus (HPV) DNA sequences in metaplastic breast carcinomas of Mexican women. BMC Cancer 2013, 13, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carolis, S.; Storci, G.; Ceccarelli, C.; Savini, C.; Gallucci, L.; Sansone, P.; Santini, D.; Seracchioli, R.; Taffurelli, M.; Fabbri, F.; et al. HPV DNA Associates With Breast Cancer Malignancy and It Is Transferred to Breast Cancer Stromal Cells by Extracellular Vesicles. Front. Oncol. 2019, 9, 860. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Bi, X.; Zhang, Y.; Zhao, P.; Zheng, T.; Dai, M. Human papillomavirus infection and sporadic breast carcinoma risk: A meta-analysis. Breast Cancer Res. Treat. 2011, 126, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Simões, P.W.; Medeiros, L.R.; Simões Pires, P.D.; Edelweiss, M.I.; Rosa, D.D.; Silva, F.R.; Silva, B.R.; Rosa, M.I. Prevalence of human papillomavirus in breast cancer: A systematic review. Int. J. Gynecol. Cancer 2012, 22, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Kudela, E.; Kudelova, E.; Kozubík, E.; Rokos, T.; Pribulova, T.; Holubekova, V.; Biringer, K. HPV-Associated Breast Cancer: Myth or Fact? Pathogens 2022, 11, 1510. [Google Scholar] [CrossRef]
- Blanco, R.; Carrillo-Beltrán, D.; Muñoz, J.P.; Corvalán, A.H.; Calaf, G.M.; Aguayo, F. Human Papillomavirus in Breast Carcinogenesis: A Passenger, a Cofactor, or a Causal Agent? Biology 2021, 10, 804. [Google Scholar] [CrossRef]
- Thomas, A.; Necchi, A.; Muneer, A.; Tobias-Machado, M.; Tran, A.T.H.; Van Rompuy, A.-S.; Spiess, P.E.; Albersen, M. Penile cancer. Nat. Rev. Dis. Primer 2021, 7, 11. [Google Scholar] [CrossRef]
- Bleeker, M.C.G.; Heideman, D.A.M.; Snijders, P.J.F.; Horenblas, S.; Dillner, J.; Meijer, C.J.L.M. Penile cancer: Epidemiology, pathogenesis and prevention. World J. Urol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Ribera-Cortada, I.; Guerrero-Pineda, J.; Trias, I.; Veloza, L.; Garcia, A.; Marimon, L.; Diaz-Mercedes, S.; Alamo, J.R.; Rodrigo-Calvo, M.T.; Vega, N.; et al. Pathogenesis of Penile Squamous Cell Carcinoma: Molecular Update and Systematic Review. Int. J. Mol. Sci. 2021, 23, 251. [Google Scholar] [CrossRef] [PubMed]
- Heidegger, I.; Borena, W.; Pichler, R. The role of human papilloma virus in urological malignancies. Anticancer Res. 2015, 35, 2513–2519. [Google Scholar] [PubMed]
- Cherif, S.; Amine, A.; Thies, S.; Taube, E.T.; Braicu, E.I.; Sehouli, J.; Kaufmann, A.M. Prevalence of human papillomavirus detection in ovarian cancer: A meta-analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1791–1802. [Google Scholar] [CrossRef]
- Ruzevick, J.; Olivi, A.; Westra, W.H. Metastatic squamous cell carcinoma to the brain: An unrecognized pattern of distant spread in patients with HPV-related head and neck cancer. J. Neurooncol. 2013, 112, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.-M.; Xu, Q.-P.; Li, X.; Xiao, R.-D.; Cai, L.; He, F. The association between human papillomavirus infection and lung cancer: A system review and meta-analysis. Oncotarget 2017, 8, 96419–96432. [Google Scholar] [CrossRef] [Green Version]
- Scinicariello, F.; Sato, T.; Lee, C.S.; Hsu, H.C.; Chan, T.S.; Tyring, S.K. Detection of human papillomavirus in primary hepatocellular carcinoma. Anticancer Res. 1992, 12, 763–766. [Google Scholar]
- St Laurent, J.; Luckett, R.; Feldman, S. HPV vaccination and the effects on rates of HPV-related cancers. Curr. Probl. Cancer 2018, 42, 493–506. [Google Scholar] [CrossRef]
- Bruni, L.; Diaz, M.; Barrionuevo-Rosas, L.; Herrero, R.; Bray, F.; Bosch, F.X.; de Sanjosé, S.; Castellsagué, X. Global estimates of human papillomavirus vaccination coverage by region and income level: A pooled analysis. Lancet Glob. Health 2016, 4, e453–e463. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Biswas, M.; Jose, T. HPV vaccine: Current status and future directions. Med. J. Armed Forces India 2015, 71, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajitani, N.; Satsuka, A.; Kawate, A.; Sakai, H. Productive Lifecycle of Human Papillomaviruses that Depends Upon Squamous Epithelial Differentiation. Front. Microbiol. 2012, 3, 152. [Google Scholar] [CrossRef] [Green Version]
- Wideroff, L.; Schiffman, M.H.; Nonnenmacher, B.; Hubbert, N.; Kirnbauer, R.; Greer, C.E.; Lowy, D.; Lorincz, A.T.; Manos, M.M.; Glass, A.G.; et al. Evaluation of Seroreactivity to Human Papillomavirus Type 16 Virus-like Particles in an Incident Case-Control Study of Cervical Neoplasia. J. Infect. Dis. 1995, 172, 1425–1430. [Google Scholar] [CrossRef]
- Carter, J.J.; Koutsky, L.A.; Wipf, G.C.; Christensen, N.D.; Lee, S.-K.; Kuypers, J.; Kiviat, N.; Galloway, D.A. The Natural History of Human Papillomavirus Type 16 Capsid Antibodies among a Cohort of University Women. J. Infect. Dis. 1996, 174, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.T.; Lowy, D.R. Papillomavirus-like particles and HPV vaccine development. Semin. Cancer Biol. 1996, 7, 373–382. [Google Scholar] [CrossRef]
- Mollers, M.; Vossen, J.M.; Scherpenisse, M.; van der Klis, F.R.M.; Meijer, C.J.L.M.; de Melker, H.E. Review: Current knowledge on the role of HPV antibodies after natural infection and vaccination: Implications for monitoring an HPV vaccination programme. J. Med. Virol. 2013, 85, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Einstein, M.H.; Baron, M.; Levin, M.J.; Chatterjee, A.; Fox, B.; Scholar, S.; Rosen, J.; Chakhtoura, N.; Lebacq, M.; van der Most, R.; et al. Comparison of the immunogenicity of the human papillomavirus (HPV)-16/18 vaccine and the HPV-6/11/16/18 vaccine for oncogenic non-vaccine types HPV-31 and HPV-45 in healthy women aged 18-45 years. Hum. Vaccines 2011, 7, 1359–1373. [Google Scholar] [CrossRef] [PubMed]
- Frazer, I.H.; Cox, J.T.; Mayeaux, E.J.; Franco, E.L.; Moscicki, A.-B.; Palefsky, J.M.; Ferris, D.G.; Ferenczy, A.S.; Villa, L.L. Advances in prevention of cervical cancer and other human papillomavirus-related diseases. Pediatr. Infect. Dis. J. 2006, 25, S65–S81, quiz S82. [Google Scholar] [CrossRef]
- Doorbar, J. Host control of human papillomavirus infection and disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Mariani, L.; Vici, P.; Suligoi, B.; Checcucci-Lisi, G.; Drury, R. Early direct and indirect impact of quadrivalent HPV (4HPV) vaccine on genital warts: A systematic review. Adv. Ther. 2015, 32, 10–30. [Google Scholar] [CrossRef]
- Yousefi, Z.; Aria, H.; Ghaedrahmati, F.; Bakhtiari, T.; Azizi, M.; Bastan, R.; Hosseini, R.; Eskandari, N. An Update on Human Papilloma Virus Vaccines: History, Types, Protection, and Efficacy. Front. Immunol. 2022, 12, 805695. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Brotons, M.; Pavón, M.A. The natural history of human papillomavirus infection. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 2–13. [Google Scholar] [CrossRef]
- Smalley Rumfield, C.; Roller, N.; Pellom, S.T.; Schlom, J.; Jochems, C. Therapeutic Vaccines for HPV-Associated Malignancies. ImmunoTargets Ther. 2020, 9, 167–200. [Google Scholar] [CrossRef] [PubMed]
- Yeo-Teh, N.; Ito, Y.; Jha, S. High-Risk Human Papillomaviral Oncogenes E6 and E7 Target Key Cellular Pathways to Achieve Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araldi, R.P.; Assaf, S.M.R.; de Carvalho, R.F.; de Carvalho, M.A.C.R.; de Souza, J.M.; Magnelli, R.F.; Módolo, D.G.; Roperto, F.P.; Stocco, R.D.C.; Beçak, W. Papillomaviruses: A systematic review. Genet. Mol. Biol. 2017, 40, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO World Health Organisation, Weekly Epidemiological Record, 16 DECEMBER 2022, No 50. Wkly. Epidemiol. Rec. 2022, 97, 645–672.
- Shapiro, G. HPV Vaccination: An Underused Strategy for the Prevention of Cancer. Curr. Oncol. 2022, 29, 3780–3792. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Doolan, K.; Hung, C.-F.; Wu, T.C. Perspectives for preventive and therapeutic HPV vaccines. J. Formos. Med. Assoc. 2010, 109, 4–24. [Google Scholar] [CrossRef] [Green Version]
- van der Burg, S.H.; Melief, C.J.M. Therapeutic vaccination against human papilloma virus induced malignancies. Curr. Opin. Immunol. 2011, 23, 252–257. [Google Scholar] [CrossRef]
- Chabeda, A.; Yanez, R.J.R.; Lamprecht, R.; Meyers, A.E.; Rybicki, E.P.; Hitzeroth, I.I. Therapeutic vaccines for high-risk HPV-associated diseases. Papillomavirus Res. 2018, 5, 46–58. [Google Scholar] [CrossRef]
- Schiller, J.T.; Davies, P. Delivering on the promise: HPV vaccines and cervical cancer. Nat. Rev. Microbiol. 2004, 2, 343–347. [Google Scholar] [CrossRef]
- WHO. WHO Guideline for Screening and Treatment of Cervical Pre-Cancer Lesions for Cervical Cancer Prevention, 2nd ed.; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Parness, J.; Horwitz, S.B. Taxol binds to polymerized tubulin in vitro. J. Cell Biol. 1981, 91, 479–487. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, H.; Luo, H.; Zhang, W.; Shen, Z.; Hu, X. Molecular mechanisms of cisplatin resistance in cervical cancer. Drug Des. Devel. Ther. 2016, 10, 1885–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Song, Y.; Mi, Y.; Jin, H.; Cao, J.; Li, H.; Han, L.; Huang, T.; Zhang, X.; Ren, S.; et al. microRNA-499a promotes the progression and chemoresistance of cervical cancer cells by targeting SOX6. Apoptosis 2020, 25, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Wang, Y.; Huang, Y.; Yu, H.; Huang, Y.; Wu, L.; Huang, L. Up-regulated miR155 reverses the epithelial-mesenchymal transition induced by EGF and increases chemo-sensitivity to cisplatin in human Caski cervical cancer cells. PLoS ONE 2012, 7, e52310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, Z.; Xu, X.; Dong, M.; Xu, J. MicroRNA-148b Acts as a Tumor Suppressor in Cervical Cancer by Inducing G1/S-Phase Cell Cycle Arrest and Apoptosis in a Caspase-3-Dependent Manner. Med. Sci. Monit. 2016, 22, 2809–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Cong, L.; Ni, G.; Chen, M.; Sun, H.; Sun, Y.; Chen, M. MicroRNA-195 inhibits the behavior of cervical cancer tumors by directly targeting HDGF. Oncol. Lett. 2017, 14, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Hasanzadeh, M.; Movahedi, M.; Rejali, M.; Maleki, F.; Moetamani-Ahmadi, M.; Seifi, S.; Hosseini, Z.; Khazaei, M.; Amerizadeh, F.; Ferns, G.A.; et al. The potential prognostic and therapeutic application of tissue and circulating microRNAs in cervical cancer. J. Cell. Physiol. 2019, 234, 1289–1294. [Google Scholar] [CrossRef]
- Liang, H.; Luo, R.; Chen, X.; Zhao, Y.; Tan, A. miR-187 inhibits the growth of cervical cancer cells by targeting FGF9. Oncol. Rep. 2017, 38, 1977–1984. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Ding, J.; Zhang, Y.; Cai, M.; Yang, J.; Cho, W.C.; Zheng, Y. microRNA-21: A key modulator in oncogenic viral infections. RNA Biol. 2021, 18, 809–817. [Google Scholar] [CrossRef]
- Chen, M.; Mao, A.; Xu, M.; Weng, Q.; Mao, J.; Ji, J. CRISPR-Cas9 for cancer therapy: Opportunities and challenges. Cancer Lett. 2019, 447, 48–55. [Google Scholar] [CrossRef]
- Lei, J.; Zhang, G. Potential antitumor applications of a monoclonal antibody specifically targeting human papilloma virus 16 E749-57 peptide: Monoclonal antibody for HPV. Microbiol. Immunol. 2012, 56, 456–462. [Google Scholar] [CrossRef]
- Wang, L.; Du, W.; Zhu, S.; Jiang, P.; Zhang, L. A high-risk papillomavirus 18 E7 affibody-enabled in vivo imaging and targeted therapy of cervical cancer. Appl. Microbiol. Biotechnol. 2019, 103, 3049–3059. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Hu, R.; Du, Y.; Tan, L.; Li, L.; Du, J.; Bai, L.; Ma, Y.; Cui, H. Immunodiagnosis and Immunotherapeutics Based on Human Papillomavirus for HPV-Induced Cancers. Front. Immunol. 2021, 11, 586796. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Spanos, W.C.; Leidner, R.; Gonçalves, A.; Martens, U.M.; Kyi, C.; Sharfman, W.; Chung, C.H.; Devriese, L.A.; Gauthier, H.; et al. Neoadjuvant nivolumab for patients with resectable HPV-positive and HPV-negative squamous cell carcinomas of the head and neck in the CheckMate 358 trial. J. Immunother. Cancer 2021, 9, e002568. [Google Scholar] [CrossRef] [PubMed]
- Balachandra, S.; Kusin, S.B.; Lee, R.; Blackwell, J.-M.; Tiro, J.A.; Cowell, L.G.; Chiang, C.-M.; Wu, S.-Y.; Varma, S.; Rivera, E.L.; et al. Blood-based biomarkers of human papillomavirus-associated cancers: A systematic review and meta-analysis. Cancer 2021, 127, 850–864. [Google Scholar] [CrossRef] [PubMed]






| Significant Feature of HPV -Associated Malignancy | Mechanism Involved | Type of Cancer | |
|---|---|---|---|
| HPV Positive Cancers | Enhanced Cell cycle progression | E6 and E7 mediated degradation of p53 and pRb | All |
| Increased radiosensitivity | Regulation of wt-p53, accumulation of double-stranded DNA breaks | HNSCC, Cervical cancer, Anal cancer | |
| Impaired DNA repair | Dysregulation of homologous recombination by the overexpression of p16, Dysregulation of non-homologous end joining by pRB | HNSCC | |
| Upregulated immune checkpoints proteins like PD-L1 and CTLA-4 | E7 mediated PD-L1 expression, Hypermethylation of DNA repair genes | HNSCC, Cervical Cancer | |
| Altered tumor immune microenvironment | Abundance of CD8+ cells, Increased levels of IL-6, IL-8 and CXCL1 | HNSCC, Cervical Cancer | |
| Accumulation of mutations | Higher APOBEC mutations, Disrupted DNA repair mechanisms | HNSCC, Cervical Cancer | |
| CpG methylation in both repetitive and non-repetitive regions | Higher Line -1 Methylation and higher genomic instability | HNSCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, I.; Harshithkumar, R.; More, A.; Mukherjee, A. Human Papilloma Virus: An Unraveled Enigma of Universal Burden of Malignancies. Pathogens 2023, 12, 564. https://doi.org/10.3390/pathogens12040564
Khan I, Harshithkumar R, More A, Mukherjee A. Human Papilloma Virus: An Unraveled Enigma of Universal Burden of Malignancies. Pathogens. 2023; 12(4):564. https://doi.org/10.3390/pathogens12040564
Chicago/Turabian StyleKhan, Ishrat, R Harshithkumar, Ashwini More, and Anupam Mukherjee. 2023. "Human Papilloma Virus: An Unraveled Enigma of Universal Burden of Malignancies" Pathogens 12, no. 4: 564. https://doi.org/10.3390/pathogens12040564
APA StyleKhan, I., Harshithkumar, R., More, A., & Mukherjee, A. (2023). Human Papilloma Virus: An Unraveled Enigma of Universal Burden of Malignancies. Pathogens, 12(4), 564. https://doi.org/10.3390/pathogens12040564