
The Identification of Streptococcus pasteurianus Obtained from Six Regions in China by Multiplex PCR Assay and the Characteristics of Pathogenicity and Antimicrobial Resistance of This Zoonotic Pathogen

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. DNA Extraction
2.3. Multiplex PCR Assay
2.4. Sensitivity and Specificity Assays
2.5. Sample Processing and S. pasteurianus Isolation
2.6. Sequencing, Assembly, Annotation, and Bioinformatics Analysis of the Genome
2.7. Antimicrobial Susceptibility Testing
2.8. Mice Infection
3. Results
3.1. Multiplex PCR Assay for S. pasteurianus
3.2. Application of the Multiplex PCR Assay
3.3. The Complete Genome Sequence of Two S. pasteurianus Strains
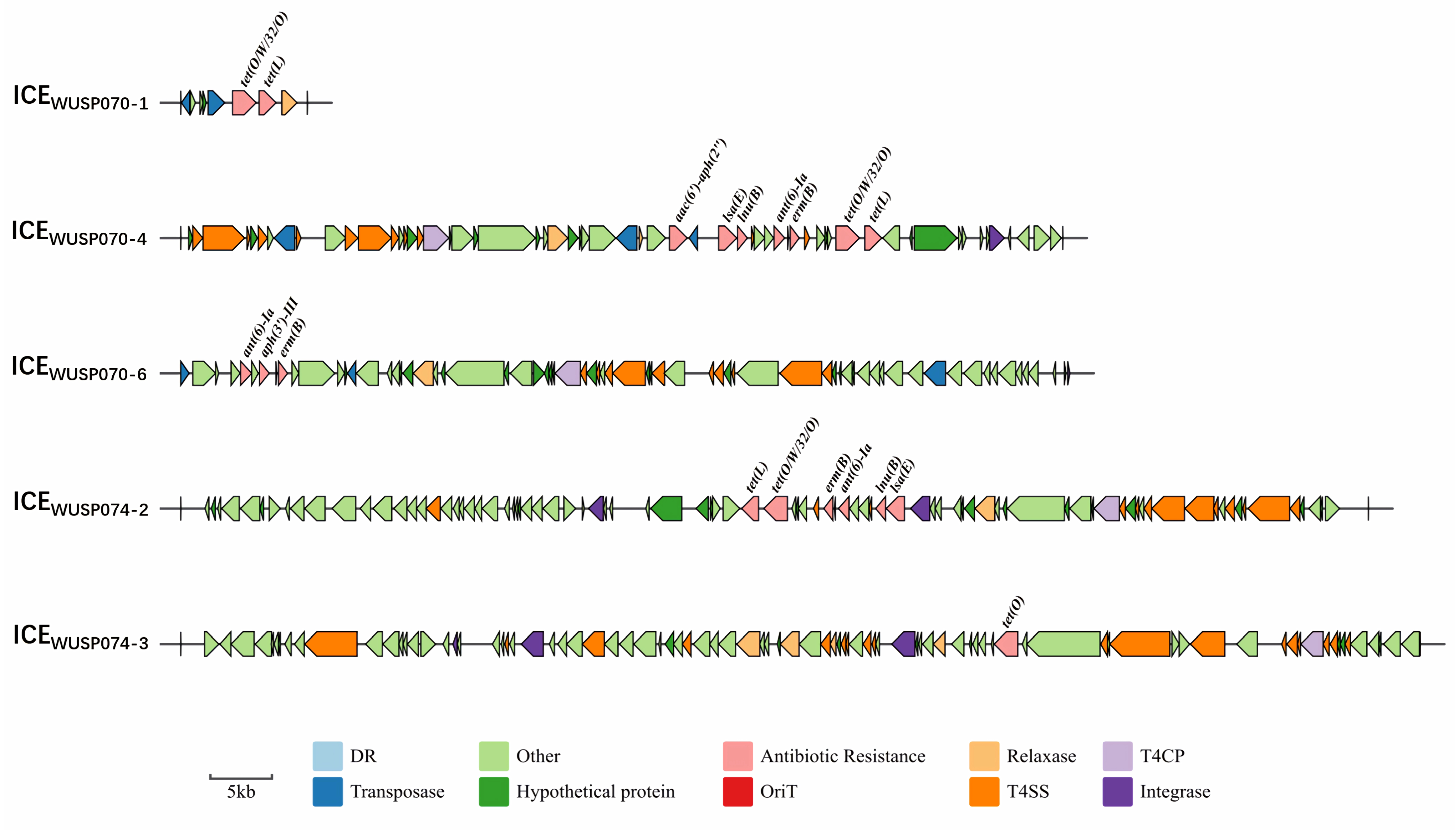
3.4. The Characteristics of Antimicrobial Resistance of Two S. pasteurianus Strains
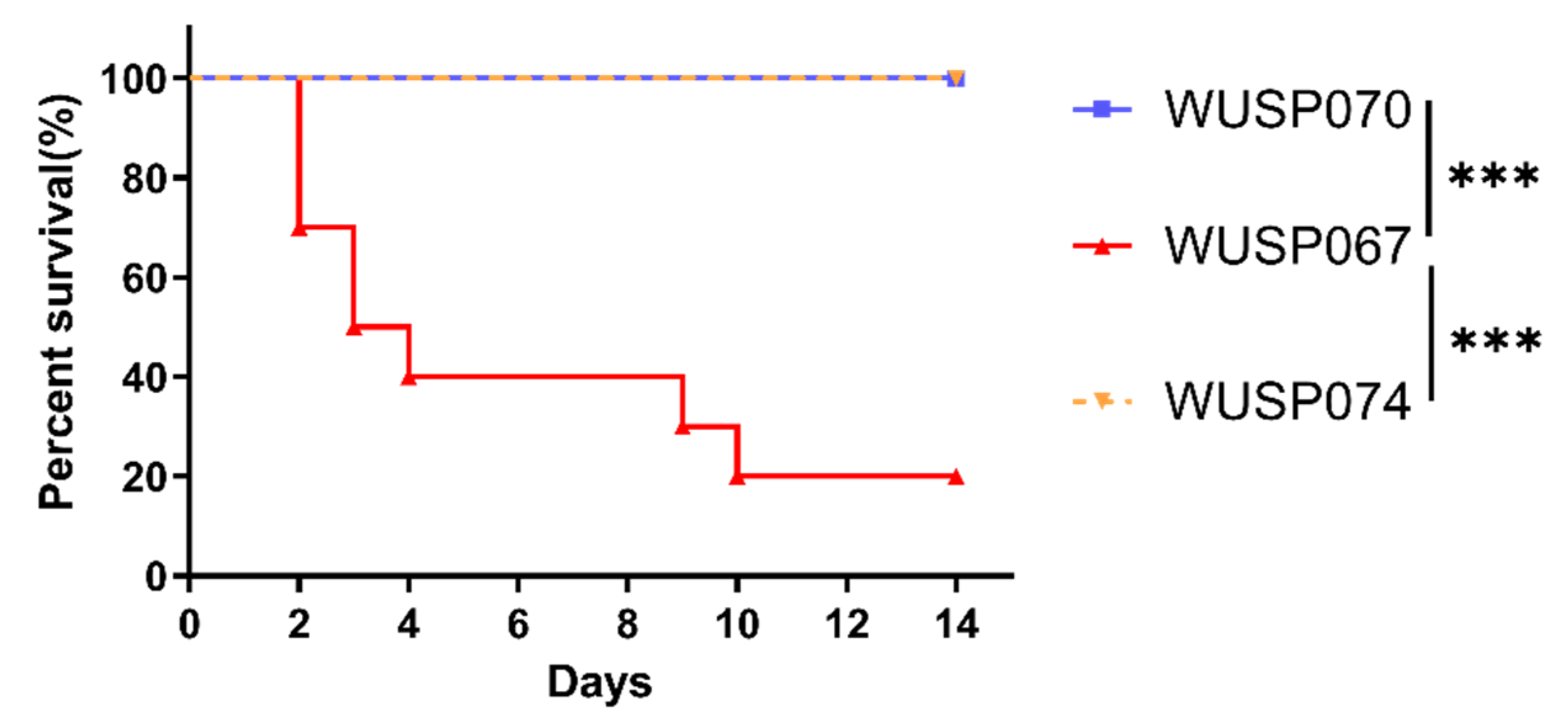
3.5. The Pathogenicity Characteristics of Two S. pasteurianus Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coykendall, A.L.; Gustafson, K.B. Deoxyribonucleic Acid Hybridizations Among Strains of Streptococcus salivarius and Streptococcus bovis. Int. J. Syst. Bacteriol. 1985, 35, 274–280. [Google Scholar] [CrossRef]
- Knight, R.G.; Shlaes, D.M. Physiological Characteristics and Deoxyribonucleic Acid Relatedness of Human Isolates of Streptococcus bovis and Streptococcus bovis (var.). Int. J. Syst. Bacteriol. 1985, 35, 357–361. [Google Scholar] [CrossRef]
- Poyart, C. Taxonomic Dissection of the Streptococcus bovis Group by Analysis of Manganese-Dependent Superoxide Dismutase Gene (SodA) Sequences: Reclassification of “Streptococcus infantarius subsp. coli” as Streptococcus lutetiensis sp. nov. and of Streptococcus bovis Biotype II.2 as Streptococcus pasteurianus sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Matesanz, M.; Rubal, D.; Iñiguez, I.; Rabuñal, R.; García-Garrote, F.; Coira, A.; García-País, M.J.; Pita, J.; Rodriguez-Macias, A.; López-Álvarez, M.J.; et al. Is Streptococcus Bovis a Urinary Pathogen? Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.C.; Dantuluri, K.; Maxey, T.; Paolillo, J. Ductal Stent Endocarditis Resulting in a Large Aortic Pseudoaneurysm. Catheter. Cardiovasc. Interv. 2021, 97, E826–E829. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Sra, H.K.; Bawa, S.; Stevens, R. Streptococcus bovis Meningitis and Hemorrhoids. J. Clin. Microbiol. 2010, 48, 2654–2655. [Google Scholar] [CrossRef] [PubMed]
- Gavin, P.J.; Thomson, R.B.; Horng, S.-J.; Yogev, R. Neonatal Sepsis Caused by Streptococcus bovis Variant (Biotype II/2): Report of a Case and Review. J. Clin. Microbiol. 2003, 41, 3433–3435. [Google Scholar] [CrossRef]
- Lu, B.; Sui, W.; Lu, X. Intrauterine Infection and Post-Partum Bacteraemia Due to Streptococcus gallolyticus subsp. pasteurianus. J. Med. Microbiol. 2013, 62, 1617–1619. [Google Scholar] [CrossRef]
- Hede, S.V.; Olarte, L.; Chandramohan, L.; Kaplan, S.L.; Hulten, K.G. Streptococcus gallolyticus subsp. Pasteurianus Infection in Twin Infants. J. Clin. Microbiol. 2015, 53, 1419–1422. [Google Scholar] [CrossRef]
- Tarakçı, N.; Dağı, H.T.; Uğur, A.R.; Tuncer, İ.; Taştekin, A. Late-Onset Streptococcus pasteurianus Sepsis in a Preterm Baby in a Neonatal Intensive Care Unit. Turk. Pediatr. Ars. 2014, 49, 157–159. [Google Scholar] [CrossRef]
- Su, Y.; Miao, B.; Wang, H.; Wang, C.; Zhang, S. Splenic Abscess Caused by Streptococcus gallolyticus subsp. pasteurianus as Presentation of a Pancreatic Cancer. J. Clin. Microbiol. 2013, 51, 4249–4251. [Google Scholar] [CrossRef] [PubMed]
- Akahane, T.; Takahashi, K.; Matsumoto, T.; Kawakami, Y. A case of peritonitis due to Streptococcus gallolyticus subsp. pasteurianus. Kansenshogaku Zasshi 2009, 83, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, J.; Alonso, M.P.; García-Garrote, F.; García-Pais, M.J.; Coira, A.; Rabuñal, R.; Gonzalez-Ramirez, A.; Pita, J.; Matesanz, M.; Velasco, D.; et al. Streptococcus bovis Group and Biliary Tract Infections: An Analysis of 51 Cases. Clin. Microbiol. Infect. 2014, 20, 405–409. [Google Scholar] [CrossRef]
- Saegeman, V.; Cossey, V.; Loens, K.; Schuermans, A.; Glaser, P. Streptococcus gallolyticus subsp. pasteurianus Infection in a Neonatal Intensive Care Unit. Pediatr. Infect. Dis. J. 2016, 35, 1272–1275. [Google Scholar] [CrossRef]
- Chand, G.; Shamban, L.; Forman, A.; Sinha, P. The Association of Streptococcus gallolyticus Subspecies pasteurianus Bacteremia with the Detection of Premalignant and Malignant Colonic Lesions. Case Rep. Gastrointest. Med. 2016, 2016, 7815843. [Google Scholar] [CrossRef]
- Barnett, J.; Ainsworth, H.; Boon, J.D.; Twomey, D.F. Streptococcus gallolyticus subsp. pasteurianus Septicaemia in Goslings. Vet. J. 2008, 176, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Hess, C.; Jandreski-Cvetkovic, D.; Liebhart, D.; Bilic, I.; Hess, M. Outbreaks of Streptococcus gallolyticus subsp. pasteurianus in Goslings Characterized by Central Nervous Symptoms. Avian Dis. 2021, 65, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gu, C.; Zhang, W.; Li, S.; Liu, J.; Qin, C.; Su, J.; Cheng, G.; Hu, X. Isolation and Characterization of Streptococcus gallolyticus subsp. pasteurianus Causing Meningitis in Ducklings. Vet. Microbiol. 2013, 162, 930–936. [Google Scholar] [CrossRef]
- Saumya, D.; Wijetunge, S.; Dunn, P.; Wallner-Pendleton, E.; Lintner, V.; Matthews, T.; Pierre, T.; Kariyawasam, S. Acute Septicemia Caused by Streptococcus gallolyticus subsp. pasteurianus in Turkey Poults. Avian Dis. 2013, 58, 318–322. [Google Scholar] [CrossRef]
- Trotta, A.; Sposato, A.; Marinaro, M.; Zizzo, N.; Passantino, G.; Parisi, A.; Buonavoglia, D.; Corrente, M. Neurological Symptoms and Mortality Associated with Streptococcus gallolyticus subsp. pasteurianus in Calves. Vet. Microbiol. 2019, 236, 108369. [Google Scholar] [CrossRef]
- Oliveira, A.R.; de Castro, M.F.; Pimentel, S.P.; de Carvalho, T.P.; Santana, C.H.; de Oliveira Santos, D.; Tinoco, H.P.; Coelho, C.M.; Pessanha, A.T.; da Paixão, T.A.; et al. Streptococcus pasteurianus-Induced Valvular Endocarditis and Sepsis in a Puerperal Emperor Tamarin (Saguinus imperator). J. Med. Primatol. 2022, 51, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ma, M.; Liang, Z.; Zhu, X.; Yao, H.; Wang, L.; Wu, Z. Pathogenic Investigations of Streptococcus pasteurianus, an Underreported Zoonotic Pathogen, Isolated from a Diseased Piglet with Meningitis. Transbound. Emerg. Dis. 2022, 69, 2609–2620. [Google Scholar] [CrossRef]
- Klatte, J.M.; Clarridge, J.E.; Bratcher, D.; Selvarangan, R. A Longitudinal Case Series Description of Meningitis Due to Streptococcus gallolyticus subsp. pasteurianus in Infants. J. Clin. Microbiol. 2012, 50, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Floret, N.; Bailly, P.; Thouverez, M.; Blanchot, C.; Alez-Martin, D.; Menget, A.; Thiriez, G.; Hoen, B.; Talon, D.; Bertrand, X. A Cluster of Bloodstream Infections Caused by Streptococcus gallolyticus Subspecies pasteurianus That Involved 5 Preterm Neonates in a University Hospital during a 2-Month Period. Infect. Control Hosp. Epidemiol. 2010, 31, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-H.; Hsueh, P.-R.; Huang, Y.-T.; Chen, P.-Y.; Tang, H.-J.; Chen, J.-M. Prolonged Streptococcus gallolyticus subsp. pasteurianus Gut Colonization in Healthcare Workers and Potential Transmission Role in Neonatal Sepsis. J. Microbiol. Immunol. Infect. 2023; in press. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Zhang, Z.; Wang, L.; Wang, J.; Zeng, J.; Yang, J.; Lu, B. Microbiological and Clinical Characteristics of Streptococcus gallolyticus subsp. pasteurianus Infection in China. BMC Infect. Dis. 2019, 19, 791. [Google Scholar] [CrossRef]
- Hatrongjit, R.; Akeda, Y.; Hamada, S.; Gottschalk, M.; Kerdsin, A. Multiplex PCR for Identification of Six Clinically Relevant Streptococci. J. Med. Microbiol. 2017, 66, 1590–1595. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Wang, M.; Goh, Y.-X.; Tai, C.; Wang, H.; Deng, Z.; Ou, H.-Y. VRprofile2: Detection of Antibiotic Resistance-Associated Mobilome in Bacterial Pathogens. Nucleic Acids Res. 2022, 50, W768–W773. [Google Scholar] [CrossRef]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the Antimicrobial Susceptibility of Bacteria Obtained from Animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, H.; Bian, C.; Chen, H.; Shen, Y.; Gao, X.; Ma, J.; Yao, H.; Wang, L.; Wu, Z. The Antimicrobial Systems of Streptococcus suis Promote Niche Competition in Pig Tonsils. Virulence 2022, 13, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Kommadath, A.; Tingley, J.P.; Abbott, D.W. Novel Insights into the Pig Gut Microbiome Using Metagenome-Assembled Genomes. Microbiol. Spectr. 2022, 10, e02380-22. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis Virulence Factors: Are They All Really Critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef]
- Liang, P.; Wang, M.; Gottschalk, M.; Vela, A.I.; Estrada, A.A.; Wang, J.; Du, P.; Luo, M.; Zheng, H.; Wu, Z. Genomic and Pathogenic Investigations of Streptococcus suis Serotype 7 Population Derived from a Human Patient and Pigs. Emerg. Microbes Infect. 2021, 10, 1960–1974. [Google Scholar] [CrossRef]
- Li, M.; Cai, C.; Chen, J.; Cheng, C.; Cheng, G.; Hu, X.; Liu, C. Inducible Expression of Both ErmB and ErmT Conferred High Macrolide Resistance in Streptococcus gallolyticus subsp. pasteurianus Isolates in China. Int. J. Mol. Sci. 2016, 17, 1599. [Google Scholar] [CrossRef]
- Cheung, M.; Pelot, M.; Nadarajah, R.; Kohl, S. Neonate with Late Onset Streptococcus bovis Meningitis: Case Report and Review of the Literature. Pediatr. Infect. Dis. J. 2000, 19, 891–893. [Google Scholar] [CrossRef]
- Sturt, A.S.; Yang, L.; Sandhu, K.; Pei, Z.; Cassai, N.; Blaser, M.J. Streptococcus gallolyticus Subspecies pasteurianus (Biotype II/2), a Newly Reported Cause of Adult Meningitis. J. Clin. Microbiol. 2010, 48, 2247–2249. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Idriss, S.; Guzman, E.; De Oliveira, E.R. Neonatal Meningitis, Endocarditis, and Pneumonitis Due to Streptococcus gallolyticus subsp. pasteurianus: A Case Report. BMC Pediatr. 2019, 19, 265. [Google Scholar] [CrossRef]
- Orbea, M.; Desai, N.; Foster, C. Invasive Streptococcus gallolyticus Infections in Infants at Texas Children’s Hospital: A 9-Year Retrospective Review. Pediatr. Infect. Dis. J. 2022, 41, e494. [Google Scholar] [CrossRef]
- Vélez Balestro, L.M.; Baroni, M.R.; Ochoteco, M.C.; Zurbriggen, M.L.; Virgolini, S.M. Streptococcus gallolyticus subsp. pasteurianus isolated from cerebrospinal fluid in a pediatric patient. Rev. Argent. Microbiol. 2013, 45, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Wardle, M.; Mu, A.; Tong, S.Y.C. Streptococcus gallolyticus subsp. pasteurianus Meningitis Complicated by Venous Sinus Thrombosis: A Case Report. Int. J. Infect. Dis. 2018, 71, 30–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-C.; Lee, P.-I.; Lin, H.-C.; Chang, L.-Y.; Lee, T.-F.; Chen, J.-M.; Hsueh, P.-R. Clustering of Streptococcus gallolyticus Subspecies pasteurianus Bacteremia and Meningitis in Neonates. J. Microbiol. Immunol. Infect. 2021, 54, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Víquez-Víquez, M.; Brenes-Meléndez, M.G.; Chacón-González, C.; Ávila-Agüero, M.L. Streptococcus gallolyticus subsp. pasteurianus: A common infection caused by an unusual agent. Bol. Med. Hosp. Infant Mex. 2021, 78, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Onoyama, S.; Ogata, R.; Wada, A.; Saito, M.; Okada, K.; Harada, T. Neonatal Bacterial Meningitis Caused by Streptococcus gallolyticus subsp. pasteurianus. J. Med. Microbiol. 2009, 58, 1252–1254. [Google Scholar] [CrossRef]
- Nagamatsu, M.; Takagi, T.; Ohyanagi, T.; Yamazaki, S.; Nobuoka, S.; Takemura, H.; Akita, H.; Miyai, M.; Ohkusu, K. Neonatal Meningitis Caused by Streptococcus gallolyticus subsp. pasteurianus. J. Infect. Chemother. 2012, 18, 265–268. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ishiwada, N.; Tanaka, J.; Okusu, K.; Ichimura, S.; Hishiki, H.; Ota, S.; Kohno, Y. Streptococcus gallolyticus subsp. pasteurianus Meningitis in an Infant. Pediatr. Int. 2014, 56, 282–285. [Google Scholar] [CrossRef]
- Takamura, N.; Kenzaka, T.; Minami, K.; Matsumura, M. Infective Endocarditis Caused by Streptococcus gallolyticus Subspecies pasteurianus and Colon Cancer. BMJ Case Rep. 2014, 2014, bcr2013203476. [Google Scholar] [CrossRef]
- Yamamura, Y.; Mihara, Y.; Nakatani, K.; Nishiguchi, T.; Ikebe, T. Unexpected Ventriculitis Complication of Neonatal Meningitis Caused by Streptococcus gallolyticus subsp. pasteurianus: A Case Report. Jpn. J. Infect. Dis. 2018, 71, 68–71. [Google Scholar] [CrossRef]
- Kasamatsu, A.; Fukushima, K.; Horiuchi, M.; Sekiya, N. Streptococcus gallolyticus Subspecies pasteurianus Bacteremia Accompanied by Acute Pancreatitis. J. Infect. Chemother. 2022, 28, 1663–1666. [Google Scholar] [CrossRef]
- Shigemori, T.; Hiasa, A.; Inoue, Y.; Oka, S.; Yasuma, T.; Nishiwaki, R.; Sugimasa, N.; Hamaguchi, T.; Noji, M.; Takeuchi, K.; et al. Acute Calculous Cholecystitis Caused by Streptococcus gallolyticus Subspecies pasteurianus: A Case Report. Microorganisms 2022, 10, 1929. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Eun, S.-H.; Kim, E.-C.; Seong, M.-W.; Kim, Y.-K. Neonatal Invasive Streptococcus gallolyticus subsp. pasteurianus Infection with Delayed Central Nervous System Complications. Korean J. Pediatr. 2015, 58, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Van Samkar, A.; Brouwer, M.C.; Pannekoek, Y.; van der Ende, A.; van de Beek, D. Streptococcus gallolyticus Meningitis in Adults: Report of Five Cases and Review of the Literature. Clin. Microbiol. Infect. 2015, 21, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Nogueira, F.; Cunha Marques, J.; Ferreira, J.P.; Almeida, J.S. Endocarditis by Streptococcus pasteurianus. Cureus 2023, 15, e34529. [Google Scholar] [CrossRef] [PubMed]
- Thatrimontrichai, A.; Chanvitan, P.; Janjindamai, W.; Dissaneevate, S.; Maneenil, G. Early Onset Neonatal Bacterial Meningitis Caused by Streptococcus gallolyticus subsp. pasteurianus. Southeast Asian J. Trop. Med. Public Health 2012, 43, 145–151. [Google Scholar] [PubMed]
- Niyas, V.K.; Arjun, R.; Sasidharan, A.; Palakunnath, G.A. Streptococcus gallolyticus Bacteremia: An Experience from a Tertiary Center in South India. Indian J. Crit. Care Med. 2020, 24, 943–945. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Origin | NCBI Accession |
|---|---|---|---|
| WUSP067 | S. pasteurianus | isolated from a newly weaned piglet’s brain with meningitis | NZ_CP039457 |
| WUSP070 | S. pasteurianus | isolated from a diarrheal cattle fecal sample | NZ_CP116957.1 |
| WUSP074 | S. pasteurianus | isolated from a healthy porcine tonsil | NZ_CP116958.1 |
| SC070731 | S. suis | isolated from a pig with meningitis | NC_020526 |
| ATCC 35246 | S. equi subsp. zooepidemicus | isolated from a dead pig | CP002904 |
| GD201008-001 | S. agalactiae | isolated from a tilapia with meningoencephalitis | NC_018646 |
| ML20171221B6-2 | S. pluranimalium | isolated from a healthy pig | |
| 1.460 | B. subtilis | China General Microbiological Culture Collection Center | |
| WUQT018 | A. hydrophila | isolated from a healthy porcine tonsil | |
| WUQT019 | K. pneumoniae | isolated from a healthy porcine tonsil | |
| WUQT020 | S. dysgalactiae | isolated from a healthy porcine tonsil | |
| WUQT022 | E. coli | isolated from a healthy porcine tonsil | |
| WUQT024 | Enterococcus sp. | isolated from a piglet lung | |
| WUQT033 | S. hyovaginalis | isolated from a piglet lung | |
| ATCC 29213 | S. aureus | American Type Culture Collection |
| Gene | Primer Name | Primer Sequence (5′−3′) | Size (bp) |
|---|---|---|---|
| E8M05_RS04035 | 1−F | GTAGATACTGATGGAGATGGT | 594 |
| 1−R | ATAATCGCCTGGTTGAGTC | ||
| E8M05_RS05155 | 2−F | TTGTTCCGTTGTCAGCATA | 767 |
| 2−R | AGCACCGATTCTATCCATAA | ||
| E8M05_RS06300 | 3−F | GTTCTGGAATGGTTAGGAATC | 409 |
| 3−R | AAGCAGCCGCAATATCAA | ||
| sodA | sodA−F | ATGGCTATTATTTTACCAAAACTAC | 609 |
| sodA−R | TCACTTTGTTGCTTTTGAGTA |
| Area | Time | Sample Type | Sample Number | Number of Positive Samples | Positive Rate (%) |
|---|---|---|---|---|---|
| Inner Mongolia | 2020 | cattle feces | 6 | 1 | 16.67 |
| Chongqing | 2021 | pig tonsil | 50 | 2 | 4.00 |
| Yunnan | 2021 | pig hilar lymph node | 50 | 18 | 36.00 |
| Jiangsu | 2021 | pig mesenteric lymph node | 50 | 0 | 0.00 |
| Guangxi | 2021 | pig hilar lymph node | 45 | 0 | 0.00 |
| pig tonsil | 30 | 0 | 0.00 | ||
| Sichuan | 2021 | pig tonsil | 60 | 3 | 5.00 |
| Classes | Antibiotics | Breakpoints for Resistance (mg/L) | MICs (mg/L) | Resistance Mechanisms |
|---|---|---|---|---|
| WUSP070 | ||||
| Macrolides | Erythromycin | ≥1 | >256 | erm(B) |
| Lincosamides | Lincomycin | ≥1 | >256 | lnu(B), erm(B), lsa(E) |
| Clindamycin | ≥1 | >256 | lnu(B), lnu(B), lsa(E) | |
| Tetracyclines | Doxycycline | ≥1 | 32 | tet(O/W/32/O), tet(L) |
| Aminoglycosides | Gentamicin | ≥16 | >256 | aac(6′)-aph(2″) |
| WUSP074 | ||||
| Macrolides | Erythromycin | ≥1 | >256 | erm(B) |
| Lincosamides | Lincomycin | ≥1 | >256 | lsa(E), lnu(B), erm(B) |
| Clindamycin | ≥1 | >256 | lsa(E), lnu(B), erm(B) | |
| Tetracyclines | Doxycycline | ≥1 | 32 | tet(O/W/32/O), tet(L), tet(O) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, M.; Wang, S.; Zhu, X.; Li, X.; Bao, Y.; Chen, X.; Wu, Z. The Identification of Streptococcus pasteurianus Obtained from Six Regions in China by Multiplex PCR Assay and the Characteristics of Pathogenicity and Antimicrobial Resistance of This Zoonotic Pathogen. Pathogens 2023, 12, 615. https://doi.org/10.3390/pathogens12040615
Ma M, Wang S, Zhu X, Li X, Bao Y, Chen X, Wu Z. The Identification of Streptococcus pasteurianus Obtained from Six Regions in China by Multiplex PCR Assay and the Characteristics of Pathogenicity and Antimicrobial Resistance of This Zoonotic Pathogen. Pathogens. 2023; 12(4):615. https://doi.org/10.3390/pathogens12040615
Chicago/Turabian StyleMa, Miaohang, Shuoyue Wang, Xinchi Zhu, Xinchun Li, Yinli Bao, Xiang Chen, and Zongfu Wu. 2023. "The Identification of Streptococcus pasteurianus Obtained from Six Regions in China by Multiplex PCR Assay and the Characteristics of Pathogenicity and Antimicrobial Resistance of This Zoonotic Pathogen" Pathogens 12, no. 4: 615. https://doi.org/10.3390/pathogens12040615
APA StyleMa, M., Wang, S., Zhu, X., Li, X., Bao, Y., Chen, X., & Wu, Z. (2023). The Identification of Streptococcus pasteurianus Obtained from Six Regions in China by Multiplex PCR Assay and the Characteristics of Pathogenicity and Antimicrobial Resistance of This Zoonotic Pathogen. Pathogens, 12(4), 615. https://doi.org/10.3390/pathogens12040615