
Multigene Phylogeny and Pathogenicity Trials Revealed Alternaria alternata as the Causal Agent of Black Spot Disease and Seedling Wilt of Pecan (Carya illinoinensis) in South Africa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Alternaria
2.2. Morphological Identification
2.3. DNA Sequencing
2.4. Phylogenetic Analyses
2.5. Pathogenicity Tests
2.6. Seedling Wilt Assay
3. Results
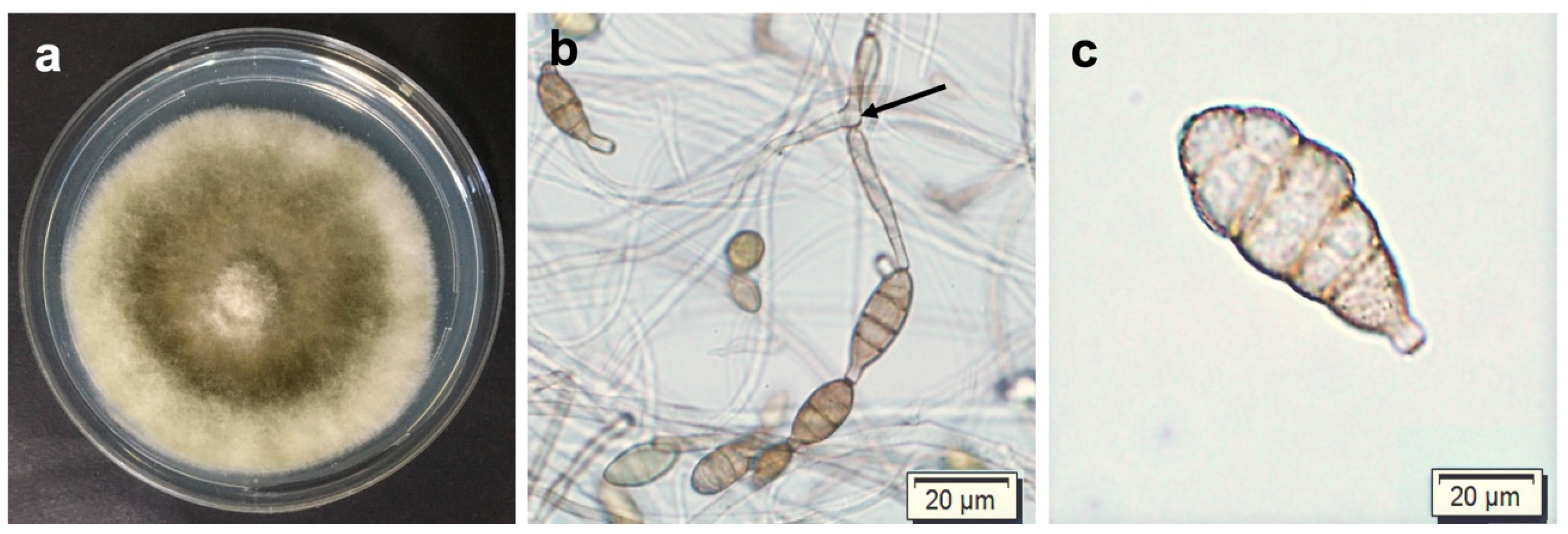
3.1. Morphological Characterization of Alternaria alternata
3.2. Phylogenetic Analyses
3.3. Pathogenicity Tests Analyses
3.4. Pathogenicity Analyses of Pecan Seedling Wilt
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, T.; Grauke, L. Pecans and Other Hickories (Carya). Acta Hortic. 1991, 290, 839–906. [Google Scholar] [CrossRef]
- Sparks, D. Adaptability of Pecan as a Species. Hortscience 2005, 40, 1175–1189. [Google Scholar] [CrossRef]
- Grauke, L.J.; Thompson, T. Pecans and Hickories. In Fruits Breeding III Nuts; Janick, J., Moore, J., Eds.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 1996; pp. 185–239. [Google Scholar]
- Wood, B. Edible Tree Nuts: Pecan and Other Hickories. In Encyclopedia of Agricultural Science; Arntzen, C., Ritter, E., Eds.; Academic Press: New York, NY, USA, 1994. [Google Scholar]
- Lemmer, B.Y.W. SA Pecan Production Still in a Boom Phase; Ryan, J., Mnyandu, E., Chetty, S., Moodley, P., Eds.; Farmer’s Weekly, Press Council: Johanessburg, South Africa, 2020; pp. 28–29. [Google Scholar]
- McEachern, G. Future Pecan Production. In Proceedings of the Texas Pecan Growers Association’s 93th Annual Conference & Trade Show, San Marcos, TX, USA, 13–16 July 2014; Texas Pecan Growers Association; Pecan South Magazine: Frisco, TX, USA, 2014. [Google Scholar]
- Zedan, D. South Africa–The Pecan Industry’s next Paradigm Shift. SA Pecan 2018, 80, 34–35. [Google Scholar]
- Sikuka, W.; Bonsu, K. Positive Outlook for South African Tree Nut Production and Trade; USDA’s Foreign Agricultural Service (FAS) and Global Agriculture Information Network (GAIN): Pretoria, South Africa, 2020. [Google Scholar]
- García-Palacios, P.; Gross, N.; Gaitán, J.; Maestre, F.T. Climate Mediates the Biodiversity–Ecosystem Stability Relationship Globally. Proc. Natl. Acad. Sci. USA 2018, 115, 8400–8405. [Google Scholar] [CrossRef]
- Rouached, H.; Tran, L.-S.P. Regulation of Plant Mineral Nutrition: Transport, Sensing and Signaling. Int. J. Mol. Sci. 2015, 16, 29717–29719. [Google Scholar] [CrossRef]
- Krüger, R. Growing a Valuable Pecan Nut Orchard. Available online: https://www.farmersweekly.co.za/farm-basics/how-to-crop/growing-a-valuable-pecan-nut-orchard/ (accessed on 13 November 2019).
- Johnson, J.D.; Black, M.C. Pecan Disease Identification and Control. In Texas Pecan Handbook; Stein, L.A., McEachern, G.R., Nesbitt, M.L., Eds.; Texas AgriLife Extension Service: College Station, TX, USA, 2012; pp. 128–179. [Google Scholar]
- Pecan Kernel Diseases. Texas A&M Forest Service. Available online: https://pecankernel.tamu.edu/diseases/ (accessed on 22 August 2019).
- Marais, G.J. Pecan Diseases in South Africa. SA Pecan 2016, 73, 26–27. [Google Scholar]
- Lourenço, V.; Moya, A.; González-Candelas, F.; Carbone, I.; Maffia, L.A.; Mizubuti, E.S.G. Molecular Diversity and Evolutionary Processes of Alternaria Solani in Brazil Inferred Using Genealogical and Coalescent Approaches. Phytopathology 2009, 99, 765–774. [Google Scholar] [CrossRef]
- Rotem, J. The Genus Alternaria: Biology, Epidemiology, and Pathogenicity; American Phytopathological Society Press: St Paul, MN, USA, 1994; ISBN 9780890541524. [Google Scholar]
- Pryor, B.M.; Gilbertson, R.L. Molecular Phylogenetic Relationships amongst Alternaria Species and Related Fungi Based upon Analysis of Nuclear ITS and Mt SSU RDNA Sequences. Mycol. Res. 2000, 104, 1312–1321. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria: An Identification Manual: Fully Illustrated and with Catalogue Raisonné 1796–2007; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007; ISBN 9070351684. [Google Scholar]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria Redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhao, M.; Huang, Y.; Liu, J.; Huang, Q.; Shu, Y.; Dong, Z. First Record of Alternaria Pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon Cablin. Pathogens 2022, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- Kgatle, M.G.; Truter, M.; Ramusi, T.M.; Flett, B.; Aveling, T.A.S. Alternaria alternata, the Causal Agent of Leaf Blight of Sunflower in South Africa. Eur. J. Plant Pathol. 2018, 151, 677–688. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The Sections of Alternaria: Formalizing Species-Group Concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Seidl, M.F.; Groenewald, J.Z.; Vries, M.D.; Stielow, J.B.; Crous, P.W. Studies in Mycology. Stud. Mycol. 2014, 77, ii. [Google Scholar] [CrossRef]
- Cai, Z.Y.; Liu, Y.X.; Li, G.H.; Wang, Y.F.; Zhou, M. First Report of Alternaria alternata Causing Black Leaf Spot of Rubber Tree in China. Plant Dis. 2015, 99, 290. [Google Scholar] [CrossRef]
- Kumar, A.; Chahal, T.S.; Hunjan, M.S.; Kaur, H. Studies of Alternaria Black Spot Disease of Pomegranate Caused by Alternaria alternata in Punjab. J. Appl. Nat. Sci. 2017, 9, 156–161. [Google Scholar] [CrossRef]
- Troncoso-Rojas, R.; Tiznado-Hernández, M.E. Alternaria alternata (Black Rot, Black Spot); Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124115682. [Google Scholar]
- Wee, J.-I.; Park, J.-H.; Back, C.-G.; You, Y.-H.; Chang, T. Mycobiology First Report of Leaf Spot Caused by Alternaria tenuissima on Black Chokeberry (Aronia melanocarpa) in Korea. Mycobiology 2016, 44, 187–190. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Zhu, P.; Liu, Y.; Zhang, Z.; Mastuda, Y.; Toyoda, H.; Xu, L. Postharvest Biological Control of Melon Pathogens Using Bacillus subtilis EXWB1. J. Plant Pathol. 2010, 92, 645–652. [Google Scholar] [CrossRef]
- Berbegal, M.; López-Cortés, I.; Salazar, D.; Gramaje, D.; Pérez-Sierra, A.; García-Jiménez, J.; Armengol, J. First Report of Alternaria Black Spot of Pomegranate Caused by Alternaria alternata in Spain. Plant Dis. 2014, 98, 689. [Google Scholar] [CrossRef]
- CABI Alternaria Gaisen (Black Spot of Japanese Pear). Available online: https://www.cabi.org/isc/datasheet/4511 (accessed on 25 December 2019).
- Zhang, M.; Zhang, Y.; Li, Y.; Bi, Y.; Mao, R.; Yang, Y.; Jiang, Q.; Prusky, D. Cellular Responses Required for Oxidative Stress Tolerance of the Necrotrophic Fungus Alternaria alternata, Causal Agent of Pear Black Spot. Microorganisms 2022, 10, 621. [Google Scholar] [CrossRef]
- Van Der Waals, J.E.; Pitsi, B.E.; Marais, C.; Wairuri, C.K. First Report of Alternaria alternata Causing Leaf Blight of Potatoes in South Africa. Plant Dis. 2011, 95, 363. [Google Scholar] [CrossRef]
- Cram, M.M. Damping-Off in Tree Nurseries. Available online: http://pnwhandbooks.dev.extension.oregonstate.edu/node/388/print (accessed on 20 February 2021).
- Marais, G.J. Plant Pathogenic Risks Associated with Pecans in South Africa. SA Pecan 2018, 78, 34–35. [Google Scholar]
- Marais, G.J. Pecan Diseases and Health Risks in South Africa. SA Pecan 2019, 81, 22–24. [Google Scholar]
- Marais, G.J. Fungi Associated with Pecan Diseases in South Africa. SA Pecan 2017, 75, 22–24. [Google Scholar]
- Woudenberg, J.; Seidl, M.; Groenewald, J.; De Vries, M.; Stielow, J.; Thomma, B.; Crous, P. Alternaria Section Alternaria: Species, Formae Speciales or Pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Aung, S.L.L.; Liu, H.F.; Pei, D.F.; Lu, B.B.; Oo, M.M.; Deng, J.X. Morphology and Molecular Characterization of a Fungus from the Alternaria alternata Species Complex Causing Black Spots on Pyrus sinkiangensis (Koerle Pear). Mycobiology 2020, 48, 233–239. [Google Scholar] [CrossRef]
- Chen, Y.J.; Meng, Q.; Zeng, L.; Tong, H.R. Phylogenetic and Morphological Characteristics of Alternaria alternata Causing Leaf Spot Disease on Camellia sinensis in China. Australas. Plant Pathol. 2018, 47, 335–342. [Google Scholar] [CrossRef]
- Nowicki, M.; Nowakowska, M.; Niezgoda, A.; Konstytucji, H.; Received, P. Aternaria Black Spot of Crucifers: Symptoms, Importance of Disease, and Perspective of Resistance Breeding. Veg. Crop. Res. Bull. 2012, 76, 5–19. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute and British Mycological Society: Kew, UK, 1970; Volume 64. [Google Scholar]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 Allergen Homologs from Alternaria and Related Taxa: Analysis of Phylogenetic Content and Secondary Structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus Phylogenetics and the Origin of Known, Highly Virulent Pathogens, Inferred from ITS and Glyceraldehyde-3-Phosphate Dehydrogenase Gene Sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular Systematics and Phylogeography of the Gibberella fujikuroi Species Complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; van der Merwe, N.A.; Crous, P.W. Diversity and Movement of Indoor Alternaria alternata across the Mainland USA. Fungal Genet. Biol. 2015, 81, 62–72. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Posada, D. JModelTest 2.0 Manual v0.1.1. Available online: http://www.phylo.org/pdf_docs/jmodeltest-2.1.6-manual.pdf (accessed on 26 October 2020).
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A Graphical Interface and Toolkit for Phylogenetic Analyses Using RAxML. Methods Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.4 [Computer Program]. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 February 2020).
- Saaiman, J. Insect-Fungal Interactions in Tree Nut Crop Orchards in South Africa, with Specific Reference to Pecans. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2014. [Google Scholar]
- Giri, P.; Taj, G.; Kumar, A. Comparison of Artificial Inoculation Methods for Studying Pathogenesis of Alternaria brassicae (Berk.) Sacc on Brassica juncea (L.) Czern. (Indian Mustard). Afr. J. Biotechnol. 2013, 12, 2422–2426. [Google Scholar] [CrossRef]
- Mohamed-Azni, I.N.A.; Sritharan, K.; Ho, S.H.; Roslan, N.D.; Arulandoo, X.; Sundram, S. Isolation, Identification and Pathogenicity of Fungi Associated with Leaf Blotches in Tenera x Tenera (TxT) Variety of Oil Palm in Malaysia. J. Plant Pathol. 2021, 104, 167–177. [Google Scholar] [CrossRef]
- Gryzenhout, M.; Khooa, B.; Landman, L. First Report of Fusarium boothii from Pecan (Carya illinoinensis) and Camel Thorn (Vachellia erioloba) Trees in South Africa. S. Afr. J. Bot. 2016, 105, 158–162. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2020, 2, 1–3559.
- RStudio Team. RStudio: Integrated Development Environment for R. Available online: https://www.rstudio.com/ (accessed on 17 June 2022).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. Available online: http://www.R-project.org (accessed on 27 June 2022).
- Popat, R.; Banakara, K. Doebioresearch: Analysis of Design of Experiments for Biological Research. Available online: http://www.r-project.org/ (accessed on 4 July 2021).
- Basim, E.; Basim, H.; Abdulai, M.; Baki, D.; Öztürk, N. Identification and Characterization of Alternaria alternata Causing Leaf Spot of Olive Tree (Olea europaea) in Turkey. Crop Prot. 2017, 92, 79–88. [Google Scholar] [CrossRef]
- Nayyar, B.G.; Woodward, S.; Mur, L.A.J.; Akram, A.; Arshad, M.; Naqvi, S.M.S.; Akhund, S. Identification and Pathogenicity of Fusarium Species Associated with Sesame (Sesamum indicum L.) Seeds from the Punjab, Pakistan. Physiol. Mol. Plant Pathol. 2018, 102, 128–135. [Google Scholar] [CrossRef]
- Bouwmester, A. SA Pecan: Celebrating 25 Years Together. SA Pecan 2017, 78, 2–3. [Google Scholar]
- Rahimloo, T.; Ghosta, Y. The Occurrence of Alternaria Species on Cabbage in Iran. Zemdirb. Agric. 2015, 102, 343–350. [Google Scholar] [CrossRef]
- Armitage, A.D.; Barbara, D.J.; Harrison, R.J.; Lane, C.R.; Sreenivasaprasad, S.; Woodhall, J.W.; Clarkson, J.P. Discrete Lineages within Alternaria alternata Species Group: Identification Using New Highly Variable Loci and Support from Morphological Characters. Fungal Biol. 2015, 119, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the Plant Tissue: The Defense of a Dangerous Passage. Front. Plant Sci. 2014, 5, 481. [Google Scholar] [CrossRef]
- Theron, N. Cladosporium cladosporioides, the Causal Organism of Pecan Scab in South Africa. Master’s Thesis, University of the Free State, Bloemfontein, South Africa, 2020. [Google Scholar]
- Wenneker, M.; Pham, K.T.K.; Woudenberg, J.H.C.; Thomma, B.P.H.J. Identification of Alternaria spp. as Causal Agent of Dead Flower Buds Disease of Pear (Pyrus communis) in the Netherlands and Methods for Disease Control. Eur. J. Plant Pathol. 2019, 155, 967–981. [Google Scholar] [CrossRef]
- Chung, K.-R. Stress Response and Pathogenicity of the Necrotrophic Fungal Pathogen Alternaria alternata. Scientifica 2012, 2012, 635431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, S.; Jiang, T.; Liu, Z.; Zhang, W.; Jian, G.; Qi, F. Chilling Stress—The Key Predisposing Factor for Causing Alternaria alternata Infection and Leading to Cotton (Gossypium hirsutum L.) Leaf Senescence. PLoS ONE 2012, 7, e36126. [Google Scholar] [CrossRef]
- Yadav, R.; Ghasolia, R.P.; Coa, S. Management of Alternaria alternata of Tomato (Lycopersicon esculentum Mill.) through SAR Activators in Natural Conditions. Int. J. Chem. Stud. 2020, 8, 2979–2981. [Google Scholar] [CrossRef]
- Mirabile, G.; Bella, P.; Vella, A.; Ferrantelli, V.; Torta, L.; Mirabile, G.; Bella, P.; Vella, A.; Ferrantelli, V.; Torta, L. Fungal Contaminants and Mycotoxins in Nuts. In Nuts and Nut Products in Human Health and Nutrition; Rao, V., Rao, L., Ahiduzzaman, M., Aminul, A., Eds.; IntechOpen: London, UK, 2021; pp. 146–165. ISBN 978-1-78985-511-1. [Google Scholar]
- Nayyar, B.G.; Woodward, S.; Mur, L.A.J.; Akram, A.; Arshad, M.; Saqlan Naqvi, S.M.; Akhund, S. The Incidence of Alternaria Species Associated with Infected Sesamum indicum L. Seeds from Fields of the Punjab, Pakistan. Plant Pathol. J. 2017, 33, 543–553. [Google Scholar] [CrossRef]
- Tang, J.; Huang, L.; Liu, Y.; Toshmatov, Z.; Zhang, C.; Shao, H. Two Phytotoxins Isolated from the Pathogenic Fungus of the Invasive Weed Xanthium italicum. Chem. Biodivers. 2020, 17, e2000043. [Google Scholar] [CrossRef] [PubMed]
- Wachowska, U.; Kwiatkowska, E.; Pluskota, W. Alternaria alternata as a Seed-Transmitted Pathogen of Sida Hermaphrodita (Malvaceae) and Its Suppression by Aureobasidium pullulans. Agriculture 2021, 11, 1264. [Google Scholar] [CrossRef]
- Ramires, F.A.; Masiello, M.; Somma, S.; Villani, A.; Susca, A.; Logrieco, A.F.; Luz, C.; Meca, G.; Moretti, A. Phylogeny and Mycotoxin Characterization of Alternaria Species Isolated from Wheat Grown in Tuscany, Italy. Toxins 2018, 10, 472. [Google Scholar] [CrossRef]
- Taylor, N.; Annandele, J.; Rossouw, W.; Kunene, S.; Zwane, M.; Human, A.; Shongwe, N.; Pandor, M.; Lotze, E.; Clulow, A.; et al. Qualifying Water Use of Mature Pecan Orchards in Selected Irrigation Areas of the Northern Cape. SA Pecan 2020, 87, 12–16. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | Tm (°C) | Amplicon Length (Approx. No. bp) | Reference |
|---|---|---|---|---|---|
| Alt a1 | Alt-F | ATG CAG TTC ACC ACC ATC GC | 63.3 °C | 980 | [42] |
| Alt-R | ACG AGG GTG AYG TAG GCG TC | ||||
| Gapdh | Gpd1 | CAA CGG CTT CGG TCG CAT TG | 58.2 °C | 860 | [43] |
| Gpd2 | GCC AAG CAG TTG GTT GTG | ||||
| Rpb2 | Rpb2-5F | GAY GAY MGW GAT CAY TTY GG | 58.2 °C | 1020 | [44] |
| Rpb2-7cR | CCC ATR GCT TGY TTR CCC AT | ||||
| Tef1 | EF1 | ATG GGT AAG GAR GAC AAG AC | 61.3 °C | 720 | [45] |
| EF2 | GGA RGT ACC AGT SAT CAT G |
| Alternaria alternata Isolate | Disease Severity (DSI) a | Rating Scale b | Aggressiveness Level c |
|---|---|---|---|
| CGJM3006 | 80.0 b | 3 | A |
| CGJM3103 | 80.0 b | 3 | A |
| CGJM3078 | 90.0 a | 3 | A |
| CGJM3136 | 80.0 b | 3 | A |
| CGJM3137 | 50.0 c | 2 | M |
| CGJM3142 | 90.0 a | 3 | A |
| Control | 0.00 d | 0 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Achilonu, C.C.; Marais, G.J.; Ghosh, S.; Gryzenhout, M. Multigene Phylogeny and Pathogenicity Trials Revealed Alternaria alternata as the Causal Agent of Black Spot Disease and Seedling Wilt of Pecan (Carya illinoinensis) in South Africa. Pathogens 2023, 12, 672. https://doi.org/10.3390/pathogens12050672
Achilonu CC, Marais GJ, Ghosh S, Gryzenhout M. Multigene Phylogeny and Pathogenicity Trials Revealed Alternaria alternata as the Causal Agent of Black Spot Disease and Seedling Wilt of Pecan (Carya illinoinensis) in South Africa. Pathogens. 2023; 12(5):672. https://doi.org/10.3390/pathogens12050672
Chicago/Turabian StyleAchilonu, Conrad Chibunna, Gert Johannes Marais, Soumya Ghosh, and Marieka Gryzenhout. 2023. "Multigene Phylogeny and Pathogenicity Trials Revealed Alternaria alternata as the Causal Agent of Black Spot Disease and Seedling Wilt of Pecan (Carya illinoinensis) in South Africa" Pathogens 12, no. 5: 672. https://doi.org/10.3390/pathogens12050672
APA StyleAchilonu, C. C., Marais, G. J., Ghosh, S., & Gryzenhout, M. (2023). Multigene Phylogeny and Pathogenicity Trials Revealed Alternaria alternata as the Causal Agent of Black Spot Disease and Seedling Wilt of Pecan (Carya illinoinensis) in South Africa. Pathogens, 12(5), 672. https://doi.org/10.3390/pathogens12050672