
Retrospective Screening of Anthrax-like Disease Induced by Bacillus tropicus str. JMT from Chinese Soft-Shell Turtles in Taiwan



Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Data Collection in Present Study
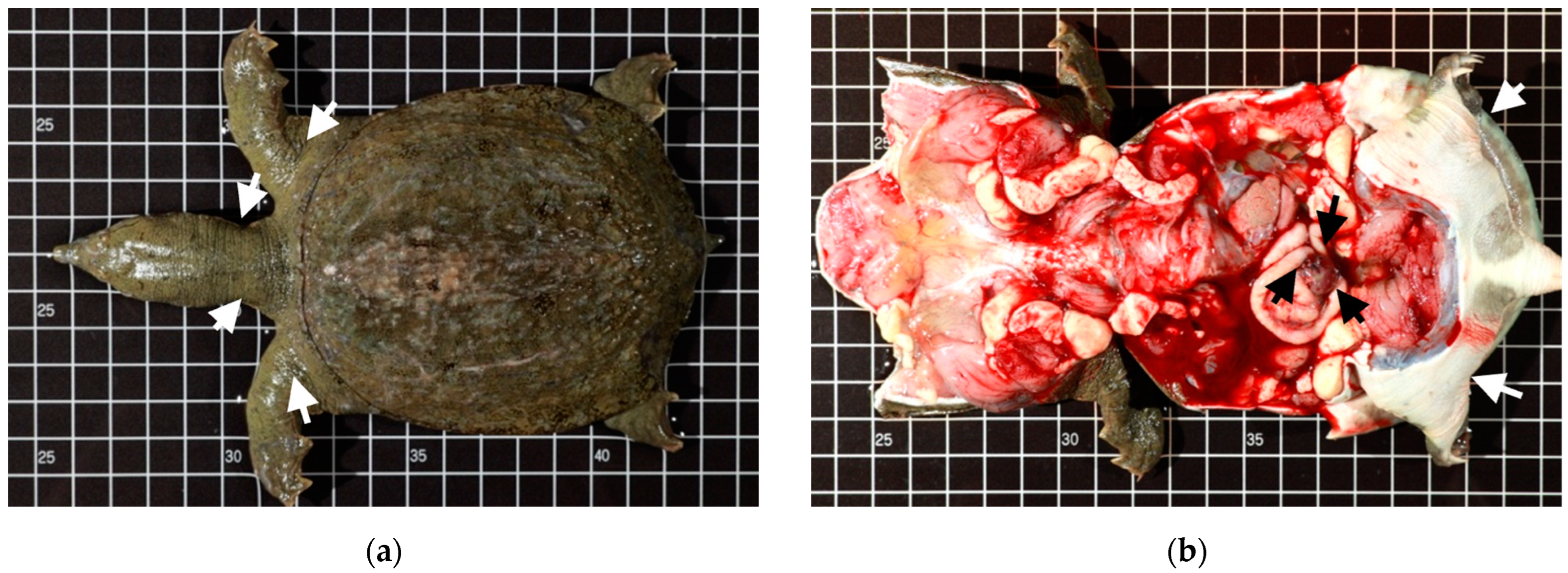
2.1.1. Case Profile of Chinese Soft-Shell Turtles
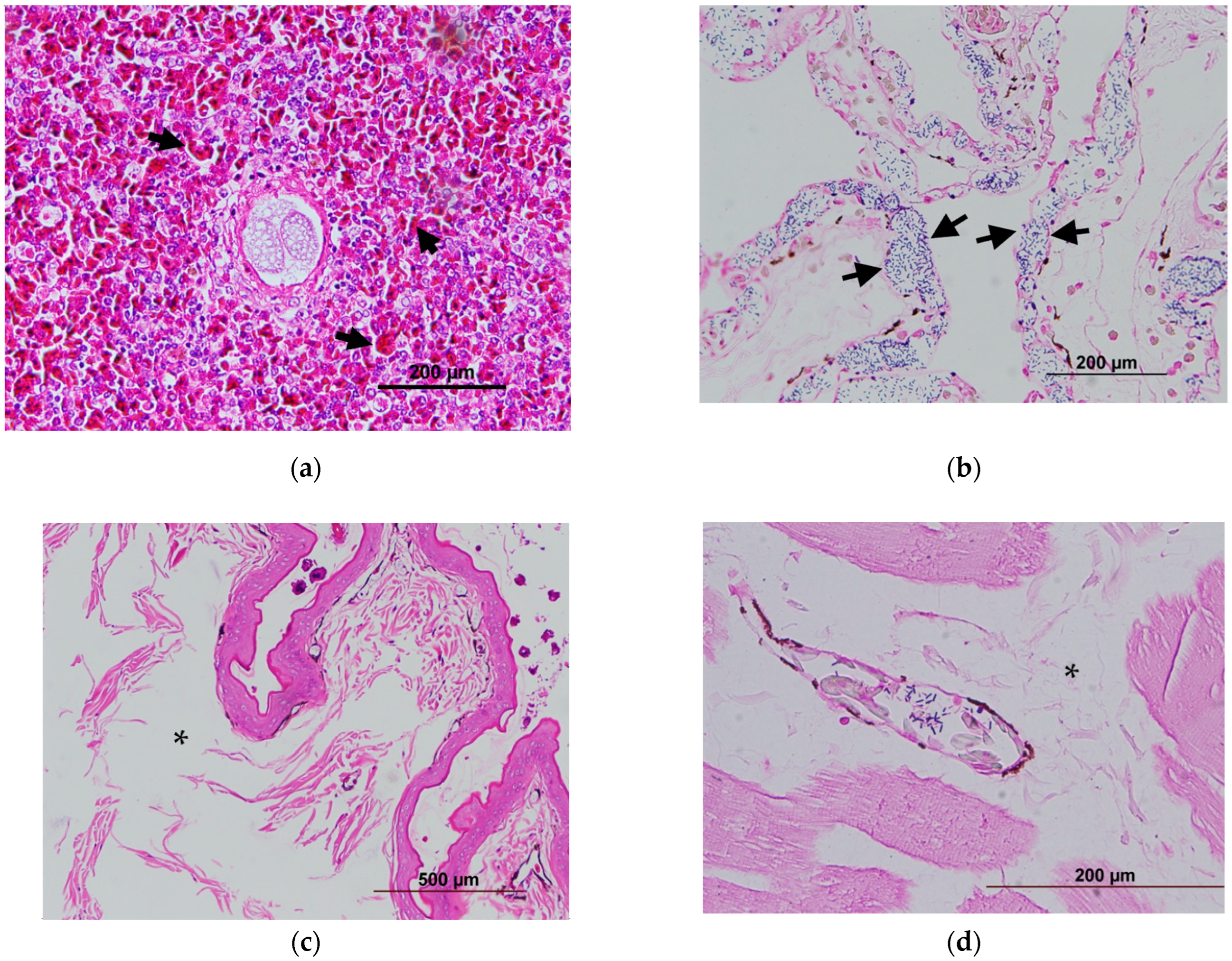
2.1.2. Identification of Bacteremia by Histopathology
2.2. Bacteriology Test
2.2.1. Bacterial Isolation, Culture and Collection
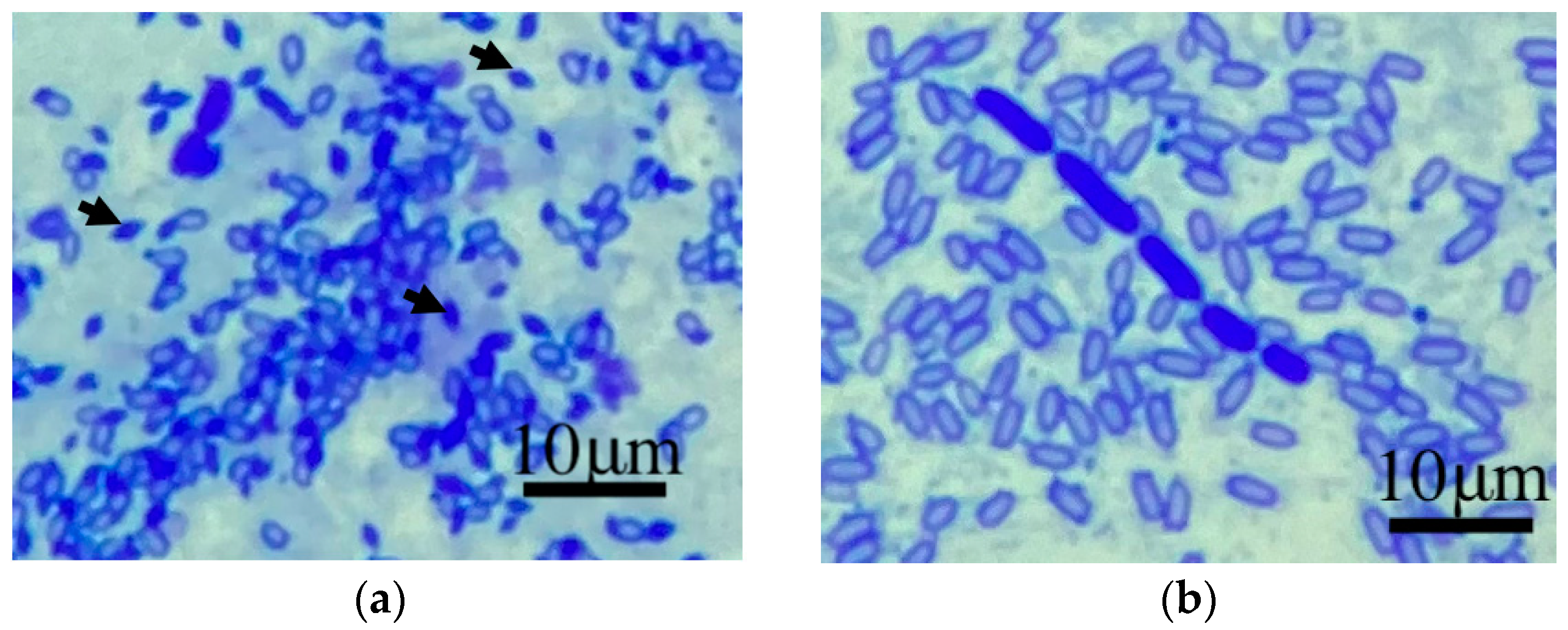
2.2.2. Parasporal Body Visualization with Coomassie Blue Stain
2.2.3. Capsule Visualization with Liu’s Stain and Negative Stain
2.2.4. Plasmid DNA and Genomic DNA Extraction
2.2.5. Oligonucleotide Primer Design
2.2.6. PCR Condition, Gene Cloning and Sequencing
2.3. Whole Genome Sequencing
2.3.1. Oxford Nanopore Technologies
2.3.2. Orthologous Average Nucleotide Identity and Digital DNA-DNA Hybridization
3. Results
3.1. Identification of Bacillus spp. JMT by Morphology and PCR
3.2. Capability of Capsule Production
3.3. Anthrax-like Virulence Genes
3.3.1. PCR Assay
3.3.2. The Absent Evidence of lef Gene via Nanopore Sequencing
3.4. Bacteremia
3.5. OrthoANI and GGD Analyses
4. Discussion
4.1. JMT Isolates Belong to Bacillus tropicus
4.2. Genome Analysis-Based Reclassification of Atypical Bacillus cereus as Bacillus tropicus
4.3. The Anthrax-like Property of B. tropicus JMT Isolated from Chinese Soft-Shell Turtle
4.4. Anthrax-like Disease
4.5. Spore Contamination
4.6. Absence of lef Gene in PCR Assay and Nanopore Sequencing
4.7. Remarkable Capsule-Production Capacity of B. tropicus JMT
4.8. A Biomarker Tool for Rapid Detecting B. tropicus JMT
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Zhu, N.; Kong, L.; Bei, Y.; Zheng, T.; Ding, X.; He, Z. First Case of Soft Shell Disease in Chinese Soft-shelled Turtle (Trionyx sinens) Associated with Aeromonas sobria–A. veronii complex. Aquaculture 2013, 406, 62–67. [Google Scholar] [CrossRef]
- Hsieh, C.; Chang, T.; Shen, Y.; Chang, C.; Tu, C.; Tung, M.; Chen, L.; Tsai, S. Pathological and PCR Detection of Mycobacteriosis in Pond-cultured Chinese Soft Shell Turtles, Trionyx sinensis. Aquaculture 2006, 261, 10–16. [Google Scholar] [CrossRef]
- Jensen, H. Mycobacterium kansasii Infection in Chinese Soft Shell Turtle (Pelodiscus sinensis). Vet. Rec. 2003, 152, 474–476. [Google Scholar]
- Tan, A.-P.; Zhao, F.; Jiang, L.; Luo, L.; Wang, W.-L.; Peng, H.-L.; Chen, Y.-L.; Zou, W.-M. Isolation and Identification of Bacillus cereus from Trionyx sinensis. Guangdong Agric. Sci. 2011, 20, 115–119. [Google Scholar]
- Yeh, S.-H. Pathological and Molecular Studies on Bacillus spp. of Soft-Shell Turtle (Trionyx sinensis). Master Thesis, National Pingtung University of Science and Technology, Pingtung, Taiwan, 2013. [Google Scholar]
- Chen, J.; Zhu, N.; Kong, L.; Bei, Y.; Zheng, T.; Ding, X.; He, Z. First Reported Fatal Bacillus thuringiensis Infections in Chinese Soft-shelled Turtles (Trionyx sinensis). Aquaculture 2014, 428, 16–20. [Google Scholar] [CrossRef]
- Cheng, L.W.; Rao, S.; Poudyal, S.; Wang, P.C.; Chen, S.C. Genotype and Virulence Gene Analyses of Bacillus cereus Group Clinical Isolates from the Chinese Softshell Turtle (Pelodiscus sinensis) in Taiwan. J. Fish Dis. 2021, 44, 1515–1529. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, Q.; Göker, M.; Meier-Kolthoff, J.P.; Wang, M.; Sun, Y.; Wang, L.; Shao, Z. Genomic Insights into the Taxonomic Status of the Bacillus cereus group. Sci. Rep. 2015, 5, 14082. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Du, J.; Lai, Q.; Zeng, R.; Ye, D.; Xu, J.; Shao, Z. Proposal of Nine Novel Species of the Bacillus cereus Group. Int. J. Syst. Evol. Microbiol. 2017, 67, 2499–2508. [Google Scholar] [CrossRef]
- Inglesby, T.V.; O’Toole, T.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Friedlander, A.M.; Gerberding, J.; Hauer, J.; Hughes, J. Anthrax as a Biological Weapon, 2002: Updated Recommendations for Management. Jama 2002, 287, 2236–2252. [Google Scholar] [CrossRef]
- Goepfert, J.; Spira, W.; Kim, H. Bacillus cereus: Food Poisoning Organism. A Review. J. Milk Food Technol. 1972, 35, 213–227. [Google Scholar] [CrossRef]
- Höfte, H.; Whiteley, H. Insecticidal Crystal Proteins of Bacillus thuringiensis. Microbiol. Mol. Biol. Rev. 1989, 53, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.; Abdullah, S. Bacillus cereus Food Poisoning: International and Indian Perspective. J. Food Sci. Technol. 2015, 52, 2500–2511. [Google Scholar] [CrossRef]
- Hoffmaster, A.R.; Ravel, J.; Rasko, D.A.; Chapman, G.D.; Chute, M.D.; Marston, C.K.; De, B.K.; Sacchi, C.T.; Fitzgerald, C.; Mayer, L.W.; et al. Identification of Anthrax Toxin Genes in a Bacillus cereus Associated with an Illness Resembling Inhalation Anthrax. Proc. Natl. Acad. Sci. USA 2004, 101, 8449–8454. [Google Scholar] [CrossRef] [PubMed]
- Firoved, A.M.; Miller, G.F.; Moayeri, M.; Kakkar, R.; Shen, Y.; Wiggins, J.F.; McNally, E.M.; Tang, W.-J.; Leppla, S.H. Bacillus anthracis Edema Toxin Causes Extensive Tissue Lesions and Rapid Lethality in Mice. Am. J. Pathol. 2005, 167, 1309–1320. [Google Scholar] [CrossRef]
- Abramova, F.A.; Grinberg, L.M.; Yampolskaya, O.V.; Walker, D.H. Pathology of Inhalational Anthrax in 42 Cases from the Sverdlovsk Outbreak of 1979. Proc. Natl. Acad. Sci. USA 1993, 90, 2291–2294. [Google Scholar] [CrossRef]
- Avashia, S.B.; Riggins, W.; Lindley, C.; Hoffmaster, A.; Drumgoole, R.; Nekomoto, T.; Jackson, P.J.; Hill, K.K.; Williams, K.; Lehman, L. Fatal Pneumonia among Metalworkers Due to Inhalation Exposure to Bacillus cereus Containing Bacillus anthracis Toxin Genes. Clin. Infect. Dis. 2007, 44, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Hoffmaster, A.R.; Hill, K.K.; Gee, J.E.; Marston, C.K.; De, B.K.; Popovic, T.; Sue, D.; Wilkins, P.P.; Avashia, S.B.; Drumgoole, R. Characterization of Bacillus cereus Isolates Associated with Fatal Pneumonias: Strains Are Closely Related to Bacillus anthracis and Harbor B. anthracis Virulence Genes. J. Clin. Microbiol. 2006, 44, 3352–3360. [Google Scholar] [CrossRef]
- Klee, S.R.; Brzuszkiewicz, E.B.; Nattermann, H.; Bruggemann, H.; Dupke, S.; Wollherr, A.; Franz, T.; Pauli, G.; Appel, B.; Liebl, W.; et al. The Genome of a Bacillus Isolate Causing Anthrax in Chimpanzees Combines Chromosomal Properties of B. cereus with B. anthracis Virulence Plasmids. PLoS ONE 2010, 5, e10986. [Google Scholar] [CrossRef]
- Klee, S.R.; Ozel, M.; Appel, B.; Boesch, C.; Ellerbrok, H.; Jacob, D.; Holland, G.; Leendertz, F.H.; Pauli, G.; Grunow, R.; et al. Characterization of Bacillus anthracis-like Bacteria Isolated from Wild Great Apes from Cote d’Ivoire and Cameroon. J. Bacteriol. 2006, 188, 5333–5344. [Google Scholar] [CrossRef]
- Leendertz, F.H.; Ellerbrok, H.; Boesch, C.; Couacy-Hymann, E.; Mätz-Rensing, K.; Hakenbeck, R.; Bergmann, C.; Abaza, P.; Junglen, S.; Moebius, Y. Anthrax Kills Wild Chimpanzees in a Tropical Rainforest. Nature 2004, 430, 451–452. [Google Scholar] [CrossRef]
- Pilo, P.; Rossano, A.; Bamamga, H.; Abdoulkadiri, S.; Perreten, V.; Frey, J. Bovine Bacillus anthracis in Cameroon. Appl. Environ. Microbiol. 2011, 77, 5818–5821. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, V.M. You Can’t B. cereus–A Review of Bacillus cereus Strains That Cause Anthrax-like Disease. Front. Microbiol. 2020, 11, 1731. [Google Scholar] [CrossRef] [PubMed]
- Heaslip, W. Bacillus tropicus, a New Species Isolated from Man and Animals Described and Compared with Other Bacilli Resembling, Bacillus anthracis. Med. J. 1941, 2, 536–540. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An Improved Algorithm and Software for Calculating Average Nucleotide Identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome Sequence-based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Ammons, D.; Rampersad, J.; Khan, A. Usefulness of Staining Parasporal Bodies When Screening for Bacillus thuringiensis. J. Invertebr. Pathol. 2002, 79, 203–204. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Goodfellow, E.S.A.M., Ed.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Park, S.-H.; Kim, H.-J.; Kim, J.-H.; Kim, T.-W.; Kim, H.-Y. Simultaneous Detection and Identification of Bacillus cereus Group Bacteria Using Multiplex PCR. J. Microbiol. Biotechnol. 2007, 17, 1177–1182. [Google Scholar]
- Wei, S.; Chelliah, R.; Park, B.-J.; Kim, S.-H.; Forghani, F.; Cho, M.S.; Park, D.-S.; Jin, Y.-G.; Oh, D.-H. Differentiation of Bacillus thuringiensis From Bacillus cereus Group Using a Unique Marker Based on Real-Time PCR. Front. Microbiol. 2019, 10, 883. [Google Scholar] [CrossRef]
- Ramisse, V.; Patra, G.; Garrigue, H.; Guesdon, J.-L.; Mock, M. Identification and Characterization of Bacillus anthracis by Multiplex PCR Analysis of Sequences on Plasmids pXO1 and pXO2 and Chromosomal DNA. FEMS Microbiol. Lett. 1996, 145, 9–16. [Google Scholar] [CrossRef]
- Chang, H.-H.; Tsai, M.-F.; Chung, C.-P.; Chen, P.-K.; Hu, H.-I.; Kau, J.-H.; Huang, H.-H.; Lin, H.-C.; Sun, D.-S. Single-step Purification of Recombinant Anthrax Lethal Factor from Periplasm of Escherichia coli. J. Biotechnol. 2006, 126, 277–285. [Google Scholar] [CrossRef]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of Nanopore Sequencing to the Genomics Community. Genome Biol. 2016, 17, 239. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Baek, I.; Lee, K.; Goodfellow, M.; Chun, J. Comparative Genomic and Phylogenomic Analyses Clarify Relationships within and between Bacillus cereus and Bacillus thuringiensis: Proposal for the Recognition of Two Bacillus thuringiensis Genomovars. Front. Microbiol. 2019, 10, 1978. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Yin, F.; Fu, C.; Chen, F.; Liu, C.; Yuan, N.; Wang, L.; Zhang, H.; Qian, D. Isolation, Identification and Pathogenicity Analysis of Bacillus cereus from Chinese Soft-shelled Turtles, Pelodiscus sinensis. Acta Hydrobiol. Sin. 2019, 43, 570–578. [Google Scholar]
- Samanta, S.; Datta, D.; Halder, G. Biodegradation Efficacy of Soil Inherent Novel sp. Bacillus tropicus (MK318648) onto Low Density Polyethylene Matrix. J. Polym. Res. 2020, 27, 324. [Google Scholar] [CrossRef]
- Thakur, P.; Singh, A.K.; Mukherjee, G. Isolation and Characterization of Alkaline Pectinase Productive Bacillus tropicus from Fruit and Vegetable Waste Dump Soil. Braz. Arch. Biol. Technol. 2021, 64, 319. [Google Scholar] [CrossRef]
- Uddin, M.; Swathi, K.; Anil, A.; Boopathy, R.; Ramani, K.; Sekaran, G. Biosequestration of Lignin in Municipal Landfill Leachate by Tailored Cationic Lipoprotein Biosurfactant through Bacillus tropicus Valorized Tannery Solid Waste. J. Environ. Manag. 2021, 300, 113755. [Google Scholar] [CrossRef]
- Brezillon, C.; Haustant, M.; Dupke, S.; Corre, J.P.; Lander, A.; Franz, T.; Monot, M.; Couture-Tosi, E.; Jouvion, G.; Leendertz, F.H.; et al. Capsules, Toxins and AtxA as Virulence Factors of Emerging Bacillus cereus Biovar anthracis. PLoS Negl. Trop. Dis. 2015, 9, e0003455. [Google Scholar] [CrossRef]
- Sue, D.; Hoffmaster, A.R.; Popovic, T.; Wilkins, P.P. Capsule Production in Bacillus cereus Strains Associated with Severe Pneumonia. J. Clin. Microbiol. 2006, 44, 3426–3428. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Budzik, J.M.; Garufi, G.; Schneewind, O. Two Capsular Polysaccharides Enable Bacillus cereus G9241 to Cause Anthrax-like Disease. Mol. Microbiol. 2011, 80, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Bhatnagar, R.; Gaur, D. Bacillus anthracis Poly-γ-d-Glutamate Capsule Inhibits Opsonic Phagocytosis by Impeding Complement Activation. Front. Immunol. 2020, 11, 462. [Google Scholar] [CrossRef]
- Owen, M.P.; Schauwers, W.; Hugh-Jones, M.E.; Kiernan, J.A.; Turnbull, P.C.; Beyer, W. A Simple, Reliable M’Fadyean Stain for Visualizing the Bacillus anthracis Capsule. J. Microbiol. Methods 2013, 92, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Luna, V.A.; Nguyen, K.; Gilling, D.H. Identification of pBC218/pBC210 Genes of Bacillus cereus G9241 in Five Florida Soils Using qPCR. Int. Sch. Res. Not. 2014, 2014, 197234. [Google Scholar] [CrossRef] [PubMed]
- Marston, C.K.; Ibrahim, H.; Lee, P.; Churchwell, G.; Gumke, M.; Stanek, D.; Gee, J.E.; Boyer, A.E.; Gallegos-Candela, M.; Barr, J.R.; et al. Anthrax Toxin-Expressing Bacillus cereus Isolated from an Anthrax-like Eschar. PLoS ONE 2016, 11, e0156987. [Google Scholar] [CrossRef]
- Pena-Gonzalez, A.; Rodriguez-R, L.M.; Marston, C.K.; Gee, J.E.; Gulvik, C.A.; Kolton, C.B.; Saile, E.; Frace, M.; Hoffmaster, A.R.; Konstantinidis, K.T. Genomic Characterization and Copy Number Variation of Bacillus anthracis Plasmids pXO1 and pXO2 in a Historical Collection of 412 Strains. MSystems 2018, 3, e00065-18. [Google Scholar] [CrossRef]
- Scorpio, A.; Chabot, D.J.; Day, W.A.; O’brien, D.K.; Vietri, N.J.; Itoh, Y.; Mohamadzadeh, M.; Friedlander, A.M. Poly-γ-glutamate Capsule-Degrading Enzyme Treatment Enhances Phagocytosis and Killing of Encapsulated Bacillus anthracis. Antimicrob. Agents Chemother. 2007, 51, 215–222. [Google Scholar] [CrossRef]
- Scorpio, A.; Chabot, D.J.; Day, W.A.; Hoover, T.A.; Friedlander, A.M. Capsule Depolymerase Overexpression Reduces Bacillus anthracis Virulence. Microbiology 2010, 156, 1459–1467. [Google Scholar] [CrossRef]
- Wilson, M.K.; Vergis, J.M.; Alem, F.; Palmer, J.R.; Keane-Myers, A.M.; Brahmbhatt, T.N.; Ventura, C.L.; O’Brien, A.D. Bacillus cereus G9241 Makes Anthrax Toxin and Capsule like Highly Virulent B. anthracis Ames but Behaves Like Attenuated Toxigenic Nonencapsulated B. anthracis Sterne in Rabbits and Mice. Infect. Immun. 2011, 79, 3012–3019. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Case (Isolate) ID. | Location | Turtle Age | Cumulative Mortality | Number of Samples | Fixed Tissues a |
|---|---|---|---|---|---|---|
| April 2016 | JMT105-1 | Changzhi Township (PT) | adult | - | 5 | − |
| May 2016 | JMT105-2 | Ligang Township (PT) | subadult | - | 8 | − |
| March 2016 | JMT105-3 | Ligang Township (PT) | adult | - | 4 | − |
| April 2016 | JMT105-4 | Jiuru Township (PT) | adult | - | 3 | − |
| April 2016 | JMT105-5 | Meinong District (K) | adult | - | 7 | − |
| April 2016 | JMT105-6 | Jiuru Township (PT) | subadult | - | 3 | − |
| May 2016 | JMT105-7 | Yanpu Township (PT) | subadult | - | 5 | − |
| May 2017 | JMT106-8 | Jiuru Township (PT) | subadult | 800/1000 (80%) | 3 | − |
| March 2017 | JMT106-9 | Changzhi Township (PT) | adult | 300/1200 (25%) | 3 | − |
| March 2017 | JMT106-10 | Ligang Township (PT) | subadult | 450/1000 (45%) | 2 | − |
| June 2017 | JMT106-11 | Changzhi Township (PT) | adult | 900/1300 (69.2%) | 4 | − |
| March 2017 | JMT106-12 | Jiuru Township (PT) | adult | 450/1350 (33.3) | 3 | − |
| April 2017 | JMT106-13 | Ligang Township (PT) | subadult | 150/700 (21.4) | 4 | − |
| May 2017 | JMT106-14 | Ligang Township (PT) | subadult | 750/1400 (53%) | 4 | − |
| April 2018 | JMT107-15 | Jiuru Township (PT) | adult | 650/900 (72.2%) | 3 | − |
| June 2018 | JMT107-16 | Jiuru Township (PT) | subadult | 380/1100 (34.5%) | 3 | − |
| May 2018 | JMT107-17 | Yanpu Township (PT) | subadult | 230/900 (25.6%) | 5 | + |
| March 2018 | JMT107-18 | Meinong District (K) | subadult | 300/1500 (20%) | 7 | + |
| March 2018 | JMT107-19 | Ligang Township (PT) | adult | 400/1300 (30.8%) | 3 | + |
| April 2018 | JMT107-20 | Ligang Township (PT) | subadult | 600/1300 (46.2%) | 3 | + |
| May 2018 | JMT107-21 | Ligang Township (PT) | adult | 1200/1200 (100%) | 2 | + |
| March 2018 | JMT107-22 | Changzhi Township (PT) | adult | 800/1450 (55.2%) | 5 | + |
| May 2019 | JMT108-23 | Changzhi Township (PT) | subadult | 130/1200 (59.1%) | 4 | + |
| May 2019 | JMT108-24 | Ligang Township (PT) | adult | 350/1200 (29.2%) | 3 | + |
| May 2019 | JMT108-25 | Ligang Township (PT) | subadult | 400/1250 (32%) | 3 | + |
| June 2019 | JMT108-26 | Yanpu Township (PT) | subadult | 350/1450 (24.1%) | 6 | + |
| March 2019 | JMT108-27 | Pingtung City (PT) | subadult | 500/130 (38.5%) | 5 | + |
| March 2019 | JMT108-28 | Ligang Township (PT) | adult | 400/900 (44.4%) | 5 | + |
| April 2020 | JMT109-29 | Yanpu Township (PT) | subadult | 700/1250 (56%) | 4 | + |
| May 2020 | JMT109-30 | Ligang Township (PT) | subadult | 130/2400 (54.2%) | 4 | + |
| May 2020 | JMT109-31 | Ligang Township (PT) | subadult | 2300/2300 (100%) | 3 | + |
| June 2020 | JMT109-32 | Jiuru Township (PT) | adult | 900/1300 (69.2%) | 3 | + |
| Species | Primer | Primer Sequence (5′-3′) | Amplicon Size (bp) | Target Gene | Reference |
|---|---|---|---|---|---|
| Bacteria | 27f | AGAGTTTGATCMTGGCTCAG | ~1500 | 16S rRNA | [28] |
| 1492r | TACGGYTACCTTGTTACGACTT | ||||
| B. cereus group | BCGSH-1F | GTGCGAACCCAATGGGTCTTC | 400 | groEL | [29] |
| BCGSH-1R | CCTTGTTGTACCACTTGCTC | ||||
| B. anthracis | BASH-2F | GGTAGATTAGCAGATTGCTCTTCAAAAGA | 253 | gyrB | [29] |
| BASH-2R | ACGAGCTTTCTCAATATCAAAATCTCCGC | ||||
| B. thuringiensis | BTJH-1F | GCTTACCAGGGAAATTGGCAG | 299 | gyrB | [29] |
| BTJH-R | ATCAACGTCGGCGTCGG | ||||
| B. cereus | BCJH-F | TCATGAAGAGCCTGTGTACG | 475 | gyrB | [29] |
| BCJH-1R | CGACGTGTCAATTCACGCGC | ||||
| B. mycoides | BMSH-F | TTTTAAGACTGCTCTAACACGTGTAAT | 603 | gyrB | [29] |
| BMSH-R | TTCAATAGCAAAATCCCCACCAAT | ||||
| B. thuringiensis | XRE1 | AAGATATTGCAAGCGGTAAGAT | 246 | XRE | [30] |
| XRE2 | GTTTTGTTTCAGCATTCCAGTAA | ||||
| B. anthracis | lef3 | CTTTTGCATATTATATCGAGC | 385 | lef | [31] |
| lef4 | GAATCACGAATATCAATTTGTAGC | ||||
| B. cereus G9241 | JMlef436f | GCGGTCATGGTGATGTAGGT | 436 | lef | This study |
| JMlef436r | ACGTTCAGTGCCTTTTCAGT | ||||
| JMpagA318f | CTGGGACGGCTCCAATCTAC | 318 | pagA | This study | |
| JMpagA318r | AGCCTGTATCCACCCTCACT | ||||
| JMcya882f | TGCACCTGACCATAGAACGG | 882 | cya | This study | |
| JMcya882r | TCCGGTTTCCTCTCAATTCCA | ||||
| JMhasA700f | GTAATGGCCACAACTGGTCACGTGAAC | 700 | hasA | This study | |
| JMhasA700r | AACGTGTTCCCCAACCATTAGA | ||||
| JMbpsB208f | AAAACAGACCCCCAATCACC | 208 | bpsB | This study | |
| JMbpsB208r | GTTTCCGCAGGTCAGCATTT |
| Bacterial Isolates | Colony Morphology | Microscopical Examination | PCR Assay | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhizoid Growth | Parasporal Body | Capsule Expression | B. c a | B. t a | XRE | bpsB | hasA | pagA | cya | lef | |
| Bacillus spp. JMT (32) | − | − | 32+ | − | 32+ | − | 32+ | 32+ | 32+ | 32+ | − |
| B. cereus ATCC 14579 (1) | − | − | − | 1+ | − | − | − | − | − | − | − |
| B. thuringiensis ATCC 10792 (1) | − | 1+ | − | − | 1+ | 1+ | − | − | − | − | − |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
|---|---|---|---|---|---|---|---|---|
| 1 | 96.64 # | 96.56 # | 96.83 # | 92.06 | 91.80 | 94.81 | 89.68 | |
| 2 | 70.20 # | 99.98 # | 97.63 # | 91.97 | 91.77 | 94.82 | 89.65 | |
| 3 | 70.10 # | 99.80 # | 97.59 # | 91.91 | 91.61 | 94.83 | 89.66 | |
| 4 | 72.20 # | 78.40 # | 78.30 # | 92.14 | 91.92 | 95.15 # | 89.58 | |
| 5 | 46.30 | 46.10 | 46.10 | 46.60 | 96.80 # | 91.71 | 89.49 | |
| 6 | 45.40 | 45.20 | 45.00 | 45.40 | 71.20 # | 91.45 | 89.58 | |
| 7 | 59.00 | 58.70 | 58.60 | 60.90 | 45.00 | 44.10 | 89.54 | |
| 8 | 38.70 | 38.60 | 38.60 | 38.50 | 38.10 | 38.30 | 38.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, J.-M.; Kuo, H.-W.; Cheng, W. Retrospective Screening of Anthrax-like Disease Induced by Bacillus tropicus str. JMT from Chinese Soft-Shell Turtles in Taiwan. Pathogens 2023, 12, 693. https://doi.org/10.3390/pathogens12050693
Tsai J-M, Kuo H-W, Cheng W. Retrospective Screening of Anthrax-like Disease Induced by Bacillus tropicus str. JMT from Chinese Soft-Shell Turtles in Taiwan. Pathogens. 2023; 12(5):693. https://doi.org/10.3390/pathogens12050693
Chicago/Turabian StyleTsai, Jia-Ming, Hsin-Wei Kuo, and Winton Cheng. 2023. "Retrospective Screening of Anthrax-like Disease Induced by Bacillus tropicus str. JMT from Chinese Soft-Shell Turtles in Taiwan" Pathogens 12, no. 5: 693. https://doi.org/10.3390/pathogens12050693
APA StyleTsai, J. -M., Kuo, H. -W., & Cheng, W. (2023). Retrospective Screening of Anthrax-like Disease Induced by Bacillus tropicus str. JMT from Chinese Soft-Shell Turtles in Taiwan. Pathogens, 12(5), 693. https://doi.org/10.3390/pathogens12050693