Mosquito Salivary Antigens and Their Relationship to Dengue and P. vivax Malaria

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Sample Collection and Diagnosis
2.2. Salivary Antigens
2.3. Human IgG Antibody Detection by ELISA
2.4. Data Analysis
3. Results
3.1. Plasmodium vivax Malaria and Exposure to the Non-Malaria Vector Aedes aegypti
3.2. DENV and Exposure to the Non-DENV Vector Anopheles albimanus
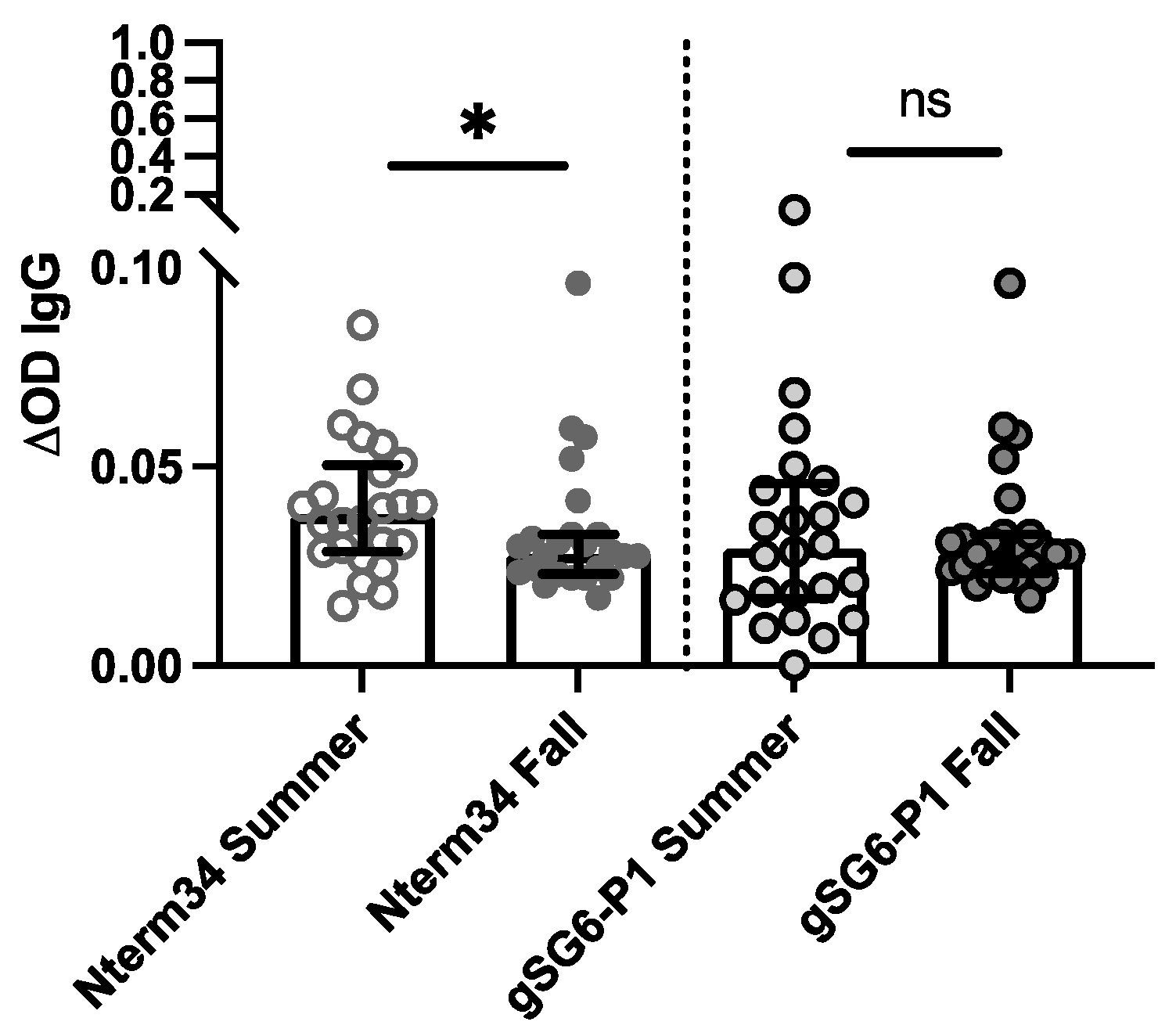
3.3. Healthy Individuals from Endemic and Non-Endemic Areas
3.4. Blood Parameters and Concordant Vector–Pathogen Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gebremariam, T.T.; Schallig, H.D.F.H.; Kurmane, Z.M.; Danquah, J.B. Increasing prevalence of malaria and acute dengue virus coinfection in Africa: A meta-analysis and meta-regression of cross-sectional studies. Malar. J. 2023, 22, 300. [Google Scholar] [CrossRef] [PubMed]
- Brathwaite Dick, O.; San Martín, J.L.; Montoya, R.H.; del Diego, J.; Zambrano, B.; Dayan, G.H. The history of dengue outbreaks in the Americas. Am. J. Trop. Med. Hyg. 2012, 87, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Romieu, A.C.; Adams, L.E.; Paz-Bailey, G. Opportunities for Improved Dengue Control in the US Territories. JAMA 2023, 330, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Bagcchi, S. Locally acquired malaria cases in the USA. Lancet Infect. Dis. 2023, 23, e401. [Google Scholar] [CrossRef] [PubMed]
- Agudelo Higuita, N.I.; Franco-Paredes, C.; Henao-Martínez, A.F.; Mendez Rojas, B.; Suarez, J.A.; Naranjo, L.; Alger, J. Migrants in transit across Central America and the potential spread of chloroquine resistant malaria-a call for action. Lancet Reg. Health Am. 2023, 22, 100505. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Guimaraes-Costa, A.B.; Valenzuela, J.G. Impact of insect salivary proteins in blood feeding, host immunity, disease, and in the development of biomarkers for vector exposure. Curr. Opin. Insect Sci. 2015, 10, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Arca, B.; Ribeiro, J.M. Saliva of hematophagous insects: A multifaceted toolkit. Curr. Opin. Insect Sci. 2018, 29, 102–109. [Google Scholar] [CrossRef]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Londoño-Rentería, B.; Cárdenas, J.C.; Giovanni, J.E.; Cárdenas, L.; Villamizar, P.; Rolón, J.; Chisenhall, D.M.; Christofferson, R.C.; Carvajal, D.J.; Pérez, O.G.; et al. Aedes aegypti anti-salivary gland antibody concentration and dengue virus exposure history in healthy individuals living in an endemic area in Colombia. Biomedica 2015, 35, 572–581. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing Aedes aegypti D7 Salivary Proteins in Colombia. Front. Public Health 2018, 6, 111. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Drame, P.M.; Weitzel, T.; Rosas, R.; Gripping, C.; Cardenas, J.C.; Alvares, M.; Wesson, D.M.; Poinsignon, A.; Remoue, F.; et al. An. gambiae gSG6-P1 evaluation as a proxy for human-vector contact in the Americas: A pilot study. Parasit. Vectors 2015, 8, 533. [Google Scholar] [CrossRef]
- Ndille, E.E.; Dubot-Pérès, A.; Doucoure, S.; Mouchet, F.; Cornelie, S.; Sidavong, B.; Fournet, F.; Remoue, F. Human IgG antibody response to Aedes aegypti Nterm-34 kDa salivary peptide as an indicator to identify areas at high risk for dengue transmission: A retrospective study in urban settings of Vientiane city, Lao PDR. Trop. Med. Int. Health 2014, 19, 576–580. [Google Scholar] [CrossRef]
- Peng, Z.; Rasic, N.; Liu, Y.; Simons, F.E. Mosquito saliva-specific IgE and IgG antibodies in 1059 blood donors. J. Allergy Clin. Immunol. 2002, 110, 816–817. [Google Scholar] [CrossRef]
- Guerra-Silveira, F.; Abad-Franch, F. Sex bias in infectious disease epidemiology: Patterns and processes. PLoS ONE 2013, 8, e62390. [Google Scholar] [CrossRef]
- Pathak, S.; Rege, M.; Gogtay, N.J.; Aigal, U.; Sharma, S.K.; Valecha, N.; Bhanot, G.; Kshirsagar, N.A.; Sharma, S. Age-dependent sex bias in clinical malarial disease in hypoendemic regions. PLoS ONE 2012, 7, e35592. [Google Scholar] [CrossRef]
- Bardach, A.E.; Garcia-Perdomo, H.A.; Alcaraz, A.; Lopez, E.T.; Gandara, R.A.R.; Ruvinsky, S.; Ciapponi, A. Interventions for the control of Aedes aegypti in Latin America and the Caribbean: Systematic Review and Meta-Analysis. Trop. Med. Int. Health 2019, 24, 530–552. [Google Scholar] [CrossRef]
- Dhiman, S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect. Dis. Poverty 2019, 8, 14. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, Y.; Li, H.; Zhu, J.; Song, W.; Chen, K.; Zhang, Y.; Lou, Y. Vaccine development for mosquito-borne viral diseases. Front. Immunol. 2023, 14, 1161149. [Google Scholar] [CrossRef]
- Qian, X.; Qi, Z. Mosquito-Borne Flaviviruses and Current Therapeutic Advances. Viruses 2022, 14, 1226. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Poinsignon, A.; Mouchet, F.; Cornelie, S.; D’Ortenzio, E.; DeHecq, J.S.; Remoue, F. Human IgG Antibody Response to Aedes Nterm-34kDa Salivary Peptide, an Epidemiological Tool to Assess Vector Control in Chikungunya and Dengue Transmission Area. PLoS Negl. Trop. Dis. 2016, 10, e0005109. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Damien, G.; Mouchet, F.; Drame, P.M.; Cornelie, S.; Noukpo, H.; Yamadjako, S.; Djenontin, A.; Moiroux, N.; et al. First attempt to validate human IgG antibody response to Nterm-34kDa salivary peptide as biomarker for evaluating exposure to Aedes aegypti bites. PLoS Negl. Trop. Dis. 2012, 6, e1905. [Google Scholar] [CrossRef]
- Poinsignon, A.; Cornelie, S.; Mestres-Simon, M.; Lanfrancotti, A.; Rossignol, M.; Boulanger, D.; Cisse, B.; Sokhna, C.; Arcà, B.; Simondon, F.; et al. Novel peptide marker corresponding to salivary protein gSG6 potentially identifies exposure to Anopheles bites. PLoS ONE 2008, 3, e2472. [Google Scholar] [CrossRef]
- Lombardo, F.; Ronca, R.; Rizzo, C.; Mestres-Simòn, M.; Lanfrancotti, A.; Currà, C.; Fiorentino, G.; Bourgouin, C.; Ribeiro, J.M.; Petrarca, V.; et al. The Anopheles gambiae salivary protein gSG6: An anopheline-specific protein with a blood-feeding role. Insect Biochem. Mol. Biol. 2009, 39, 457–466. [Google Scholar] [CrossRef]
- Drame, P.M.; Poinsignon, A.; Dechavanne, C.; Cottrell, G.; Farce, M.; Ladekpo, R.; Massougbodji, A.; Cornelie, S.; Courtin, D.; Migot-Nabias, F.; et al. Specific antibodies to Anopheles gSG6-P1 salivary peptide to assess early childhood exposure to malaria vector bites. Malar. J. 2015, 14, 285. [Google Scholar] [CrossRef]
- Drame, P.M.; Poinsignon, A.; Besnard, P.; Cornelie, S.; Le Mire, J.; Toto, J.C.; Foumane, V.; Dos-Santos, M.A.; Sembène, M.; Fortes, F.; et al. Human antibody responses to the Anopheles salivary gSG6-P1 peptide: A novel tool for evaluating the efficacy of ITNs in malaria vector control. PLoS ONE 2010, 5, e15596. [Google Scholar] [CrossRef]
- Sagna, A.B.; Yobo, M.C.; Elanga Ndille, E.; Remoue, F. New Immuno-Epidemiological Biomarker of Human Exposure to Aedes Vector Bites: From Concept to Applications. Trop. Med. Infect. Dis. 2018, 3, 80. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Drame, P.M.; Montiel, J.; Vasquez, A.M.; Tobon-Castano, A.; Taylor, M.; Vizcaino, L.; Lenhart, A.A.E. Identification and Pilot Evaluation of Salivary Peptides from Anopheles albimanus as Biomarkers for Bite Exposure and Malaria Infection in Colombia. Int. J. Mol. Sci. 2020, 21, 691. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Montiel, J.; Calvo, E.; Tobon-Castano, A.; Valdivia, H.O.; Escobedo-Vargas, K.; Romero, L.; Bosantes, M.; Fisher, M.L.; Conway, M.J.; et al. Antibody Responses against Anopheles darlingi Immunogenic Peptides in Plasmodium Infected Humans. Front. Cell Infect. Microbiol. 2020, 10, 455. [Google Scholar] [CrossRef]
- Montiel, J.; Carbal, L.F.; Tobón-Castaño, A.; Vásquez, G.M.; Fisher, M.L.; Londono-Rentería, B. IgG antibody response against Anopheles salivary gland proteins in asymptomatic Plasmodium infections in Narino, Colombia. Malar. J. 2020, 19, 42. [Google Scholar] [CrossRef]
- Visser, I.; Koenraadt, C.J.M.; Koopmans, M.P.G.; Rockx, B. The significance of mosquito saliva in arbovirus transmission and pathogenesis in the vertebrate host. One Health 2023, 16, 100506. [Google Scholar] [CrossRef]
- Bellone, R.; Failloux, A.B. The Role of Temperature in Shaping Mosquito-Borne Viruses Transmission. Front. Microbiol. 2020, 11, 584846. [Google Scholar] [CrossRef]
- Rozo-Lopez, P.; Mengual, X. Updated list of the mosquitoes of Colombia (Diptera: Culicidae). Biodivers. Data J. 2015, e4567. [Google Scholar] [CrossRef]
- Cano-Pérez, E.; González-Beltrán, M.; Ampuero, J.S.; Gómez-Camargo, D.; Morrison, A.C.; Astete, H. Prevalence of Mosquito Populations in the Caribbean Region of Colombia with Important Public Health Implications. Trop. Med. Infect. Dis. 2022, 8, 11. [Google Scholar] [CrossRef]
- Baik, L.S.; Nave, C.; Au, D.D.; Guda, T.; Chevez, J.A.; Ray, A.; Holmes, T.C. Circadian Regulation of Light-Evoked Attraction and Avoidance Behaviors in Daytime- versus Nighttime-Biting Mosquitoes. Curr. Biol. 2020, 30, 3252–3259.e3253. [Google Scholar] [CrossRef]
- Tazeen, A.; Abdullah, M.; Hisamuddin, M.; Ali, S.; Naqvi, I.H.; Verma, H.N.; Ahmed, A.; Parveen, S. Concurrent Infection with Plasmodium vivax and the Dengue and Chikungunya Viruses in a Paediatric Patient from New Delhi, India in 2016. Intervirology 2017, 60, 48–52. [Google Scholar] [CrossRef]
- Eder, M.; Cortes, F.; Teixeira de Siqueira Filha, N.; Araujo de Franca, G.V.; Degroote, S.; Braga, C.; Ridde, V.; Turchi Martelli, C.M. Scoping review on vector-borne diseases in urban areas: Transmission dynamics, vectorial capacity and co-infection. Infect. Dis. Poverty 2018, 7, 90. [Google Scholar] [CrossRef]
- Cardenas, J.C.; Giraldo-Parra, S.Y.; Gonzalez, M.U.; Gutierrez-Silva, L.Y.; Jaimes-Villamizar, L.; Roa-Parra, A.L.; Carvajal, D.J.; Valdivia, H.O.; Sanchez, J.F.; Colpitts, T.M.; et al. Laboratory Findings in Patients with Probable Dengue Diagnosis from an Endemic Area in Colombia in 2018. Viruses 2021, 13, 1401. [Google Scholar] [CrossRef]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E4209–E4218. [Google Scholar] [CrossRef]
- Busula, A.O.; Bousema, T.; Mweresa, C.K.; Masiga, D.; Logan, J.G.; Sauerwein, R.W.; Verhulst, N.O.; Takken, W.; de Boer, J.G. Gametocytemia and Attractiveness of Plasmodium falciparum-Infected Kenyan Children to Anopheles gambiae Mosquitoes. J. Infect. Dis. 2017, 216, 291–295. [Google Scholar] [CrossRef]
- Yobo, C.M.; Sadia-Kacou, C.A.M.; Adja, M.A.; Elanga-Ndille, E.; Sagna, A.B.; Guindo-Coulibaly, N.; Poinsignon, A.; Remoue, F.; Koudou, B.G. Evaluation of Human Exposure to Aedes Bites in Rubber and Palm Cultivations Using an Immunoepidemiological Biomarker. Biomed. Res. Int. 2018, 2018, 3572696. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Eisele, T.P.; Keating, J.; James, M.A.; Wesson, D.M. Antibody response against Anopheles albimanus (Diptera: Culicidae) salivary protein as a measure of mosquito bite exposure in Haiti. J. Med. Entomol. 2010, 47, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Cardenas, J.C.; Cardenas, L.D.; Christofferson, R.C.; Chisenhall, D.M.; Wesson, D.M.; McCracken, M.K.; Carvajal, D.; Mores, C.N. Use of anti-Aedes aegypti salivary extract antibody concentration to correlate risk of vector exposure and dengue transmission risk in Colombia. PLoS ONE 2013, 8, e81211. [Google Scholar] [CrossRef] [PubMed]
- Olajiga, O.M.; Marin-Lopez, A.; Cardenas, J.C.; Gutierrez-Silva, L.Y.; Gonzales-Pabon, M.U.; Maldonado-Ruiz, L.P.; Worges, M.; Fikrig, E.; Park, Y.; Londono-Renteria, B. Aedes aegypti anti-salivary proteins IgG levels in a cohort of DENV-like symptoms subjects from a dengue-endemic region in Colombia. Front. Epidemiol. 2022, 2, 1002857. [Google Scholar] [CrossRef]
- Rizzo, C.; Lombardo, F.; Ronca, R.; Mangano, V.; Sirima, S.B.; Nèbiè, I.; Fiorentino, G.; Modiano, D.; Arcà, B. Differential antibody response to the Anopheles gambiae gSG6 and cE5 salivary proteins in individuals naturally exposed to bites of malaria vectors. Parasit. Vectors 2014, 7, 549. [Google Scholar] [CrossRef] [PubMed]
- Doucoure, S.; Mouchet, F.; Cournil, A.; Le Goff, G.; Cornelie, S.; Roca, Y.; Giraldez, M.G.; Simon, Z.B.; Loayza, R.; Misse, D.; et al. Human antibody response to Aedes aegypti saliva in an urban population in Bolivia: A new biomarker of exposure to Dengue vector bites. Am. J. Trop. Med. Hyg. 2012, 87, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Debebe, Y.; Hill, S.R.; Birgersson, G.; Tekie, H.; Ignell, R. Plasmodium falciparum gametocyte-induced volatiles enhance attraction of Anopheles mosquitoes in the field. Malar. J. 2020, 19, 327. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol. 2005, 3, e298. [Google Scholar] [CrossRef] [PubMed]
- Wampfler, R.; Mwingira, F.; Javati, S.; Robinson, L.; Betuela, I.; Siba, P.; Beck, H.P.; Mueller, I.; Felger, I. Strategies for detection of Plasmodium species gametocytes. PLoS ONE 2013, 8, e76316. [Google Scholar] [CrossRef]
- Baton, L.A.; Ranford-Cartwright, L.C. Do malaria ookinete surface proteins P25 and P28 mediate parasite entry into mosquito midgut epithelial cells? Malar. J. 2005, 4, 15. [Google Scholar] [CrossRef]
- Essuman, E.; Grabias, B.; Verma, N.; Chorazeczewski, J.K.; Tripathi, A.K.; Mlambo, G.; Addison, E.A.; Amoah, A.G.B.; Quakyi, I.; Oakley, M.S.; et al. A Novel Gametocyte Biomarker for Superior Molecular Detection of the Plasmodium falciparum Infectious Reservoirs. J. Infect. Dis. 2017, 216, 1264–1272. [Google Scholar] [CrossRef]
- Schneider, P.; Reece, S.E.; van Schaijk, B.C.; Bousema, T.; Lanke, K.H.; Meaden, C.S.; Gadalla, A.; Ranford-Cartwright, L.C.; Babiker, H.A. Quantification of female and male Plasmodium falciparum gametocytes by reverse transcriptase quantitative PCR. Mol. Biochem. Parasitol. 2015, 199, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Dash, M.; Sachdeva, S.; Bansal, A.; Sinha, A. Gametogenesis in Plasmodium: Delving Deeper to Connect the Dots. Front. Cell Infect. Microbiol. 2022, 12, 877907. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.A.; Cardenas, R.; Yañez, J.; Petzold, M.; Kroeger, A. Risk of dengue, Zika, and chikungunya transmission in the metropolitan area of Cucuta, Colombia: Cross-sectional analysis, baseline for a cluster-randomised controlled trial of a novel vector tool for water containers. BMC Public Health 2023, 23, 1000. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Lerma, J.; Solarte, Y.A.; Giraldo-Calderón, G.I.; Quiñones, M.L.; Ruiz-López, F.; Wilkerson, R.C.; González, R. Malaria vector species in Colombia: A review. Mem. Inst. Oswaldo Cruz 2011, 106 (Suppl. S1), 223–238. [Google Scholar] [CrossRef] [PubMed]
- Poinsignon, A.; Cornelie, S.; Ba, F.; Boulanger, D.; Sow, C.; Rossignol, M.; Sokhna, C.; Cisse, B.; Simondon, F.; Remoue, F. Human IgG response to a salivary peptide, gSG6-P1, as a new immuno-epidemiological tool for evaluating low-level exposure to Anopheles bites. Malar. J. 2009, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Sagna, A.B.; Sarr, J.B.; Gaayeb, L.; Drame, P.M.; Ndiath, M.O.; Senghor, S.; Sow, C.S.; Poinsignon, A.; Seck, M.; Hermann, E.; et al. gSG6-P1 salivary biomarker discriminates micro-geographical heterogeneity of human exposure to Anopheles bites in low and seasonal malaria areas. Parasit. Vectors 2013, 6, 68. [Google Scholar] [CrossRef]
- Ndo, C.; Elanga-Ndille, E.; Cheteug, G.; Metitsi, R.D.; Wanji, S.; Moukoko, C.E.E. IgG antibody responses to Anopheles gambiae gSG6-P1 salivary peptide are induced in human populations exposed to secondary malaria vectors in forest areas in Cameroon. PLoS ONE 2022, 17, e0276991. [Google Scholar] [CrossRef]
- Shieh, J.N.; Rossingnol, P.A. Opposite influences of host anaemia on blood feeding rate and fecundity of mosquitoes. Parasitology 1992, 105 Pt 2, 159–163. [Google Scholar] [CrossRef]
- Zhu, Y.; Tong, L.; Nie, K.; Wiwatanaratanabutr, I.; Sun, P.; Li, Q.; Yu, X.; Wu, P.; Wu, T.; Yu, C.; et al. Host serum iron modulates dengue virus acquisition by mosquitoes. Nat. Microbiol. 2019, 4, 2405–2415. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.; Liu, Z.; Peng, Y.; Peng, W.; Tong, L.; Wang, J.; Liu, Q.; Wang, P.; Cheng, G. A volatile from the skin microbiota of flavivirus-infected hosts promotes mosquito attractiveness. Cell 2022, 185, 2510–2522.e2516. [Google Scholar] [CrossRef]
- Minakawa, N.; Sonye, G.; Mogi, M.; Githeko, A.; Yan, G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J. Med. Entomol. 2002, 39, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Asgarian, T.S.; Moosa-Kazemi, S.H.; Sedaghat, M.M. Impact of meteorological parameters on mosquito population abundance and distribution in a former malaria endemic area, central Iran. Heliyon 2021, 7, e08477. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.J.; Kim, H.C.; Klein, T.A.; Chong, S.T.; Sim, K.; Chung, Y.; Cheong, H.K. Comparison of climatic factors on mosquito abundance at US Army Garrison Humphreys, Republic of Korea. PLoS ONE 2020, 15, e0240363. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Cayan, D.; Tyree, M.; Barker, C.M.; Eldridge, B.; Dettinger, M. Impact of climate variation on mosquito abundance in California. J. Vector Ecol. 2008, 33, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.W.; Ionides, E.L.; Knepper, R.G.; Stanuszek, W.W.; Walker, E.D.; Wilson, M.L. Cross-correlation map analyses show weather variation influences on mosquito abundance patterns in Saginaw County, Michigan, 1989–2005. J. Med. Entomol. 2012, 49, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, L.P.; Montenegro-Cadena, L.; Blattner, B.; Menghwar, S.; Zurek, L.; Londono-Renteria, B. Differential Tick Salivary Protein Profiles and Human Immune Responses to Lone Star Ticks (Amblyomma americanum) from the Wild vs. a Laboratory Colony. Front. Immunol. 2019, 10, 1996. [Google Scholar] [CrossRef]
- Robertson, L.C.; Prior, S.; Apperson, C.S.; Irby, W.S. Bionomics of Anopheles quadrimaculatus and Culex erraticus (Diptera: Culicidae) in the Falls Lake basin, North Carolina: Seasonal changes in abundance and gonotrophic status, and host-feeding patterns. J. Med. Entomol. 1993, 30, 689–698. [Google Scholar] [CrossRef]
- Noblesse DeMoss, H. Biological and Taxonomic Observations on the Mosquitoes of Kansas. Trans. Kans. Acad. Sci. 1939, 42, 255–265. [Google Scholar]
- Dantur Juri, M.J.; Claps, G.L.; Santana, M.; Zaidenberg, M.; Almirón, W.R. Abundance patterns of Anopheles pseudopunctipennis and Anopheles argyritarsis in northwestern Argentina. Acta Trop. 2010, 115, 234–241. [Google Scholar] [CrossRef]
- Peng, Z.; Ho, M.K.; Li, C.; Simons, F.E. Evidence for natural desensitization to mosquito salivary allergens: Mosquito saliva specific IgE and IgG levels in children. Ann. Allergy Asthma Immunol. 2004, 93, 553–556. [Google Scholar] [CrossRef]
- Anker, M.; Arima, Y. Male-female differences in the number of reported incident dengue fever cases in six Asian countries. West. Pac. Surveill. Response J. 2011, 2, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.E.; Eliason, D.A.; Moore, M.; Sather, G.E.; Schonberger, L.B.; Cabrera-Coello, L.; Fernandez de Castro, J. Epidemiologic investigations of dengue infection in Mexico, 1980. Am. J. Epidemiol. 1983, 117, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Trravassos da Rosa, A.P.; Vasconcelos, P.F.; Travassos Da Rosa, E.S.; Rodrigues, S.G.; Mondet, B.; Cruz, A.C.; Sousa, M.R.; Travassos Da Rosa, J.F. Dengue epidemic in Belém, Pará, Brazil, 1996–1997. Emerg. Infect. Dis. 2000, 6, 298–301. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.M.; O’Donnell, J.S. Platelets in malaria pathogenesis. Blood 2018, 132, 1222–1224. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Abreu, C.; Harris, M.; Shrader, J.; Sarvepalli, S. Severe Thrombocytopenia Associated with Dengue Fever: An Evidence-Based Approach to Management of Thrombocytopenia. Case Rep. Hematol. 2022, 2022, 3358325. [Google Scholar] [CrossRef]
- de Azeredo, E.L.; Monteiro, R.Q.; de-Oliveira Pinto, L.M. Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediat. Inflamm. 2015, 2015, 313842. [Google Scholar] [CrossRef] [PubMed]
- Ghanchi, N.K.; Khan, M.H.; Arain, M.A.; Zubairi, M.B.A.; Raheem, A.; Khan, M.A.; Beg, M.A. Hematological Profile and Gametocyte Carriage in Malaria Patients from Southern Pakistan. Cureus 2019, 11, e4256. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; van der Heijden, W.; Reuling, I.J.; Wan, J.; Yan, Q.; de Laat-Kremers, R.M.W.; Van der Ven, A.J.; de Groot, P.G.; McCall, M.; Sauerwein, R.W.; et al. Functional changes in hemostasis during asexual and sexual parasitemia in a controlled human malaria infection. PLoS ONE 2022, 17, e0271527. [Google Scholar] [CrossRef]
- Cox, D.; McConkey, S. The role of platelets in the pathogenesis of cerebral malaria. Cell. Mol. Life Sci. 2010, 67, 557–568. [Google Scholar] [CrossRef]
- Islam, A.; Emran, T.B.; Yamamoto, D.S.; Iyori, M.; Amelia, F.; Yusuf, Y.; Yamaguchi, R.; Alam, M.S.; Silveira, H.; Yoshida, S. Anopheline antiplatelet protein from mosquito saliva regulates blood feeding behavior. Sci. Rep. 2019, 9, 3129. [Google Scholar] [CrossRef]
- Champagne, D.E.; Smartt, C.T.; Ribeiro, J.M.; James, A.A. The salivary gland-specific apyrase of the mosquito Aedes aegypti is a member of the 5′-nucleotidase family. Proc. Natl. Acad. Sci. USA 1995, 92, 694–698. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Status | All Age Years (Range) | Female (Range) | Male (Range) |
|---|---|---|---|
| Malaria | 33.4 (1–67), n = 49 | 36.0 (16–53), n = 14 | 32.4 (1–67), n = 35 |
| Dengue | 15.3 (1–76), n = 124 | 17.6 (1–76), n = 70 | 12.4 (1–69), n = 54 |
| Healthy | 28.9 (2–79), n = 103 | 27.0 (2–79), n = 65 | 32.3 (2–72), n = 38 |
| Nterm-34kDa | Pvs25 | Pvs230 | Parasite Count |
|---|---|---|---|
| All | −0.0109 (p = 0.9473) | −0.1553 (p = 0.3453) | −0.0314 (p = 0.8495) |
| Females | −0.5000 (p = 0.0819) | −0.7253 (p = 0.0050) | −0.3077 (p = 0.3064) |
| Males | 0.1829 (p = 0.3711) | 0.0715 (p = 0.7287) | 0.2151 (p = 0.2913) |
| Nterm-34kDa | Red Blood Cell Count | White Blood Cell Count | Platelet Count | Hemoglobin | Hematocrit |
|---|---|---|---|---|---|
| All | −0.3288 (p = 0.0313) | 0.2363 (p = 0.1271) | −0.2078 (p = 0.1812) | −0.1402 (p = 0.3699) | −0.1374 (p = 0.3797) |
| Females | −01560 (p = 0.5942) | 0.5923 (p = 0.0256) | −0.5560 (p = 0.0389) | −0.0683 (p = 0.8166) | −0.0989 (p = 0.7366) |
| Males | −0.4171 (p = 0.0244) | 0.0561 (p = 0.7727) | −0.0017 (p = 0.9929) | −0.1594 (p = 0.4090) | −0.1264 (p = 0.5135) |
| Peptide | Age |
|---|---|
| All | |
| Peroxi-P1 | −0.0207 (p = 0.8880) |
| Trans-1 | 0.3270 (p = 0.0218) |
| Trans-2 | 0.1596 (p = 2733) |
| An. albimanus SGE | 0.0471 (p = 0.07478) |
| gSG6-P1 | 0.2064 (p = 0.1547) |
| Nterm-34kDa | 0.0828 (p = 0.5719) |
| Females | |
| Peroxi-P1 | 0.3645 (p = 0.2001) |
| Trans-1 | 0.6196 (p = 0.0181) |
| Trans-2 | 0.3934 (p = 0.1641) |
| An. albimanus SGE | 0.5934 (p = 0.0253) |
| gSG6-P1 | 0.5334 (p = 0.0495) |
| Nterm-34kDa | 0.2356 (p = 0.4175) |
| Males | |
| Peroxi-P1 | −0.1172 (p = 0.5027) |
| Trans-1 | 0.2603 (p = 0.1310) |
| Trans-2 | 0.0793 (p = 0.6507) |
| An. albimanus SGE | −0.0771 (p = 0.6599) |
| gSG6-P1 | 0.1113 (p = 0.5243) |
| Nterm-34kDa | 0.0417 (p = 0.8121) |
| Peptide | Red Blood Cell Count | White Blood Cell Count | Platelet Count | Hemoglobin | Hematocrit |
|---|---|---|---|---|---|
| All | |||||
| AnDarApy-1 | 0.0955 (p = 0.2934) | 0.0277 (p = 0.7607) | −0.0831 (p = 0.3609) | 0.0999 (p = 0.2715) | 0.1092 (p = 0.2294) |
| gSG6-P1 | 0.1807 (p = 0.0455) | −0.0829 (p = 0.3617) | −0.0484 (p = 0.5949) | 0.0921 (p = 0.3112) | −0.1097 (p = 0.2269) |
| Females | |||||
| AnDarApy-1 | −0.0571 (p = 0.6390) | −0.0641 (p = 0.5980) | 0.0739 (p = 0.5434) | 0.0847 (p = 0.4858) | 0.0520 (p = 0. 6689) |
| gSG6-P1 | 0.0216 (p = 0.8591) | −0.1233 (p = 0.3092) | 0.0338 (p = 0.7815) | 0.0015 (p = 0.9899) | −0.0156 (p = 0.8981) |
| Males | |||||
| AnDarApy-1 | 0.2797 (p = 0.0425) | 0.2063 (p = 0.1382) | −0.3052 (p = 0.0263) | 0.1138 (p = 0.4173) | 0.1681 (p = 0.2290) |
| gSG6-P1 | 0.4099 (p = 0.0023) | 0.0559 (p = 0.6908) | −0.2038 (p = 0.1433) | 0.2074 (p = 0.1362) | 0.2408 (p = 0.0824) |
| Peptide | Age |
|---|---|
| All | |
| gSG6-P1 | −0.3533 (p = 0.0003) |
| Nterm-34kDa | −0.4182 (p = 0.0000) |
| Correlations by gender | |
| Females | |
| gSG6-P1 | −0.3575 (p = 0.0035) |
| Nterm-34kDa | −0.4087 (p = 0.0007) |
| Males | |
| gSG6-P1 | −0.3702 (p = 0.0370) |
| Nterm-34kDa | −0.4105 (p = 0.0196) |
| Correlations by location | |
| Colombia | |
| gSG6-P1 | 0.2702 (p = 0.0481) |
| Nterm-34kDa | 0.0796 (p = 0.5675) |
| US | |
| gSG6-P1 | −0.4772 (p = 0.0005) |
| Nterm-34kDa | −0.4049 (p = 0.0039) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howell, M.M.; Olajiga, O.M.; Cardenas, J.C.; Parada-Higuera, C.A.; Gonzales-Pabon, M.U.; Gutierrez-Silva, L.Y.; Jaimes-Villamizar, L.; Werner, B.M.; Shaffer, J.G.; Manuzak, J.A.; et al. Mosquito Salivary Antigens and Their Relationship to Dengue and P. vivax Malaria. Pathogens 2024, 13, 52. https://doi.org/10.3390/pathogens13010052
Howell MM, Olajiga OM, Cardenas JC, Parada-Higuera CA, Gonzales-Pabon MU, Gutierrez-Silva LY, Jaimes-Villamizar L, Werner BM, Shaffer JG, Manuzak JA, et al. Mosquito Salivary Antigens and Their Relationship to Dengue and P. vivax Malaria. Pathogens. 2024; 13(1):52. https://doi.org/10.3390/pathogens13010052
Chicago/Turabian StyleHowell, McKenna M., Olayinka M. Olajiga, Jenny C. Cardenas, Claudia A. Parada-Higuera, Maria U. Gonzales-Pabon, Lady Y. Gutierrez-Silva, Lucy Jaimes-Villamizar, Brett M. Werner, Jeffrey G. Shaffer, Jennifer A. Manuzak, and et al. 2024. "Mosquito Salivary Antigens and Their Relationship to Dengue and P. vivax Malaria" Pathogens 13, no. 1: 52. https://doi.org/10.3390/pathogens13010052
APA StyleHowell, M. M., Olajiga, O. M., Cardenas, J. C., Parada-Higuera, C. A., Gonzales-Pabon, M. U., Gutierrez-Silva, L. Y., Jaimes-Villamizar, L., Werner, B. M., Shaffer, J. G., Manuzak, J. A., & Londono-Renteria, B. (2024). Mosquito Salivary Antigens and Their Relationship to Dengue and P. vivax Malaria. Pathogens, 13(1), 52. https://doi.org/10.3390/pathogens13010052