
Inflammatory CD11b+ Macrophages Produce BAFF in Spleen of Mice Infected with Leishmania donovani


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Parasites
2.2. Experimental Infection
2.3. Flow Cytometric Analysis
2.4. Magnetic Separation of Splenocytes
2.5. BAFF Measurement
2.6. Immunohistochemical Analysis
2.7. Immunohistochemical Analysis
2.8. Western Blotting
3. Results
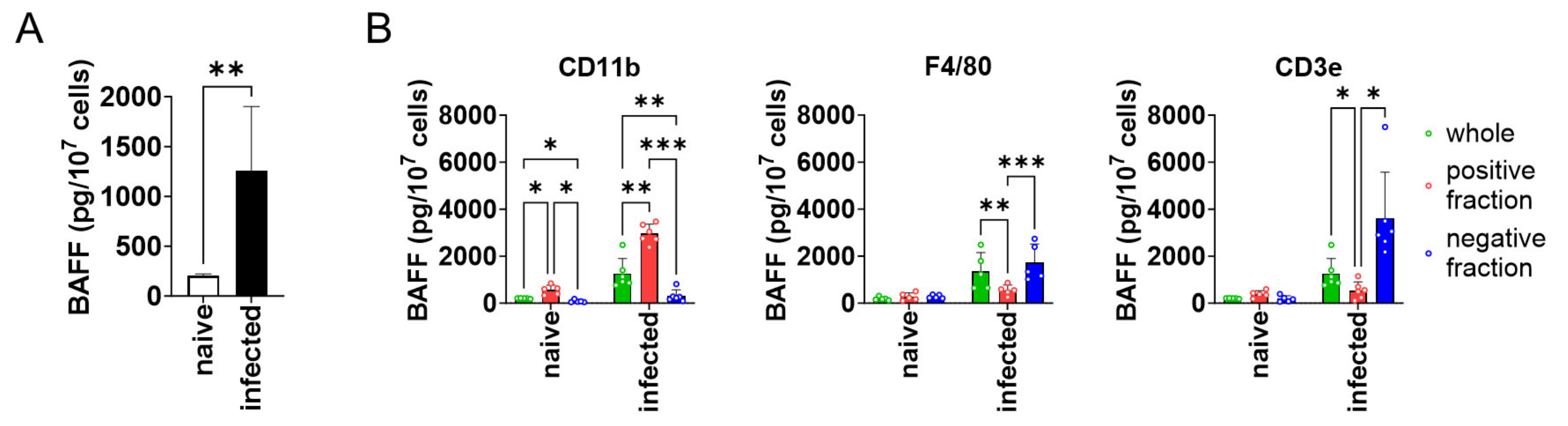
3.1. CD11b+ Cells in the Spleen-Produced BAFF during L. donovani Infection
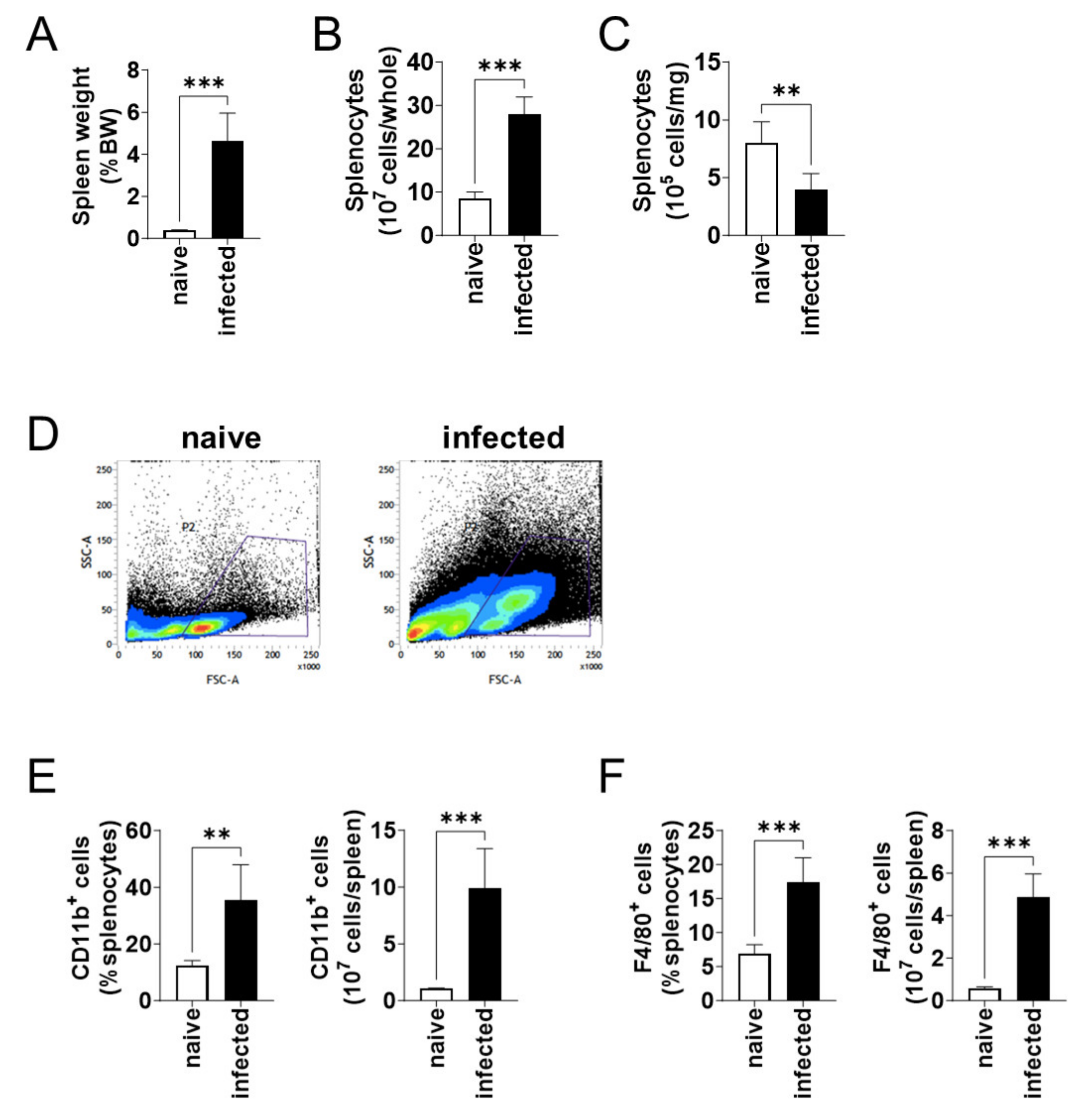
3.2. The Number of CD11b+ Cells in the Spleen Increased during L. donovani Infection
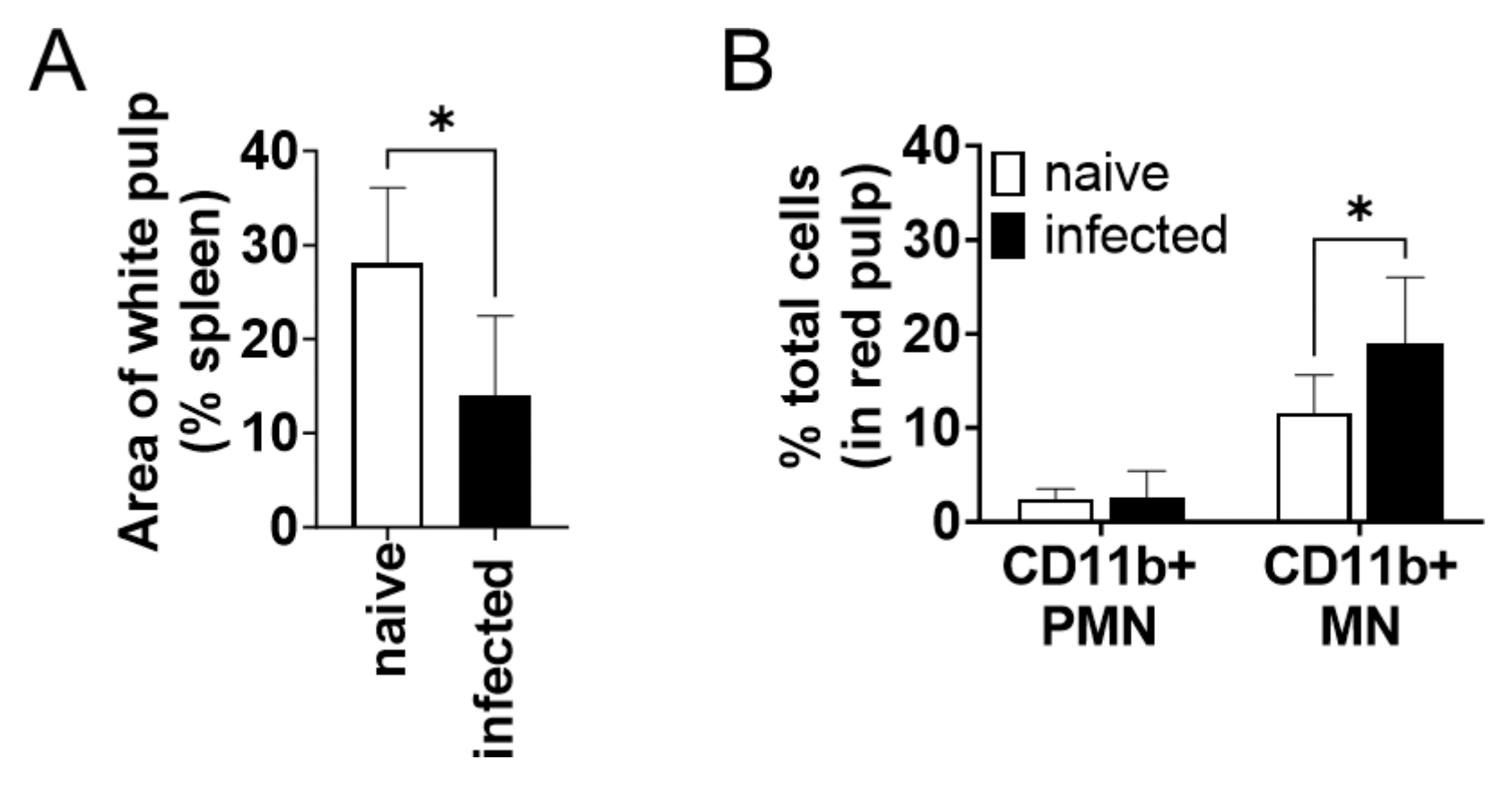
3.3. CD11b+ Mononuclear Cells Accumulated in the Spleen during L. donovani Infection
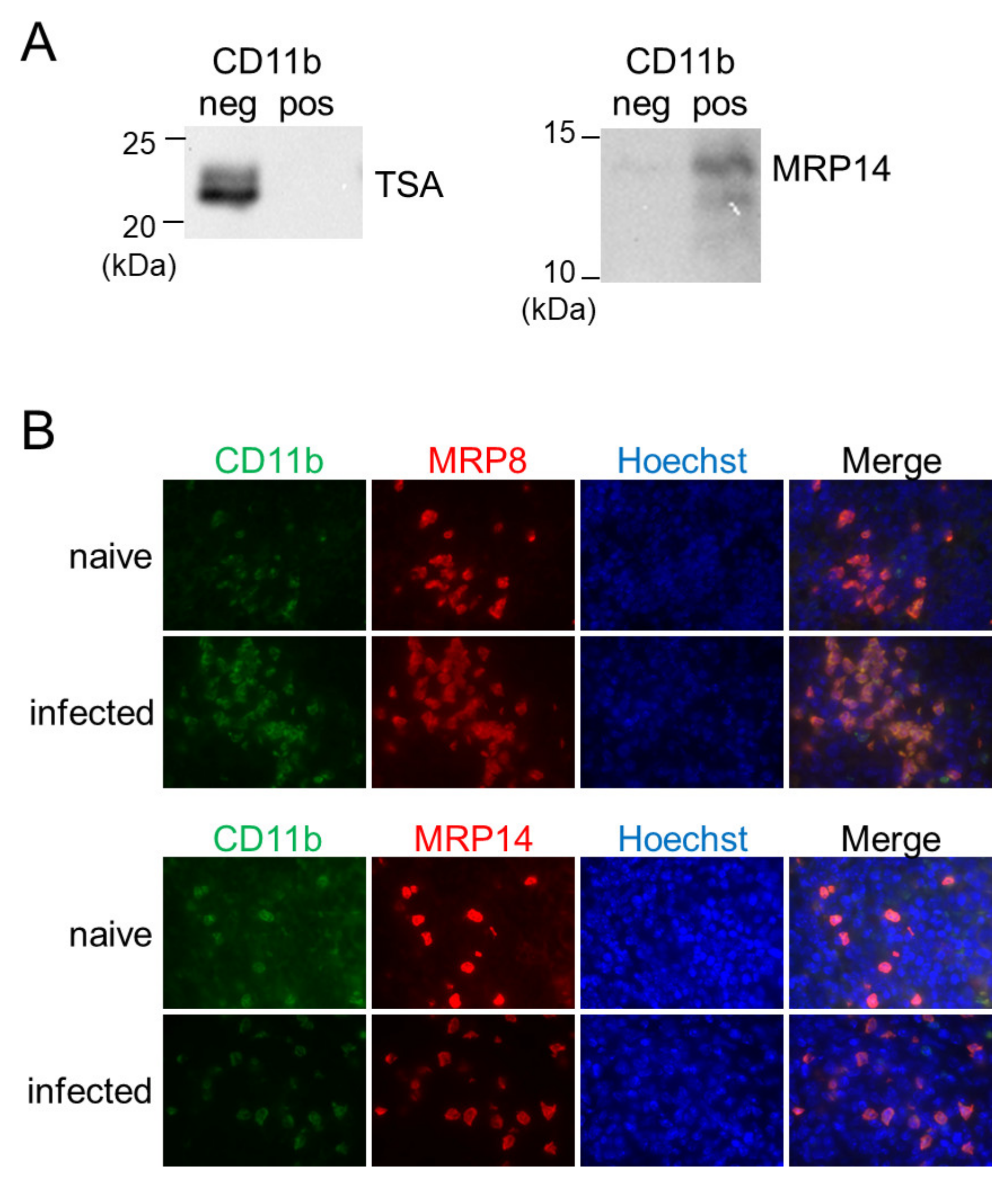
3.4. CD11b+ Cells in the Spleen during Infection Are MRP8/14-Positive and Are Distinct from Host Macrophages for L. donovani
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, P.A.; Belvedere, O.; Orr, A.; Pieri, K.; LaFleur, D.W.; Feng, P.; Soppet, D.; Charters, M.; Gentz, R.; Parmelee, D.; et al. BLyS: Member of the Tumor Necrosis Factor Family and B Lymphocyte Stimulator. Science 1999, 285, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Hase, H.; Kanno, Y.; Kojima, M.; Hasegawa, K.; Sakurai, D.; Kojima, H.; Tsuchiya, N.; Tokunaga, K.; Masawa, N.; Azuma, M.; et al. BAFF/BLyS Can Potentiate B-Cell Selection with the B-Cell Coreceptor Complex. Blood 2004, 103, 2257–2265. [Google Scholar] [CrossRef] [PubMed]
- Tardivel, A.; Tinel, A.; Lens, S.; Steiner, Q.; Sauberli, E.; Wilson, A.; Mackay, F.; Rolink, A.G.; Beermann, F.; Tschopp, J.; et al. The Anti-apoptotic Factor Bcl-2 Can Functionally Substitute for the B Cell Survival but Not for the Marginal Zone B Cell Differentiation Activity of BAFF. Eur. J. Immunol. 2004, 34, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Derudder, E.; Hobeika, E.; Pelanda, R.; Reth, M.; Rajewsky, K.; Schmidt-Supprian, M. Canonical NF-ΚB Activity, Dispensable for B Cell Development, Replaces BAFF-Receptor Signals and Promotes B Cell Proliferation upon Activation. Immunity 2006, 24, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Rauch, M.; Tussiwand, R.; Bosco, N.; Rolink, A.G. Crucial Role for BAFF-BAFF-R Signaling in the Survival and Maintenance of Mature B Cells. PLoS ONE 2009, 4, e5456. [Google Scholar] [CrossRef] [PubMed]
- Tangye, S.G.; Bryant, V.L.; Cuss, A.K.; Good, K.L. BAFF, APRIL and Human B Cell Disorders. Semin. Immunol. 2006, 18, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Mackay, F.; Woodcock, S.A.; Lawton, P.; Ambrose, C.; Baetscher, M.; Schneider, P.; Tschopp, J.; Browning, J.L. Mice Transgenic for Baff Develop Lymphocytic Disorders along with Autoimmune Manifestations. J. Exp. Med. 1999, 190, 1697–1710. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.D.; Sarosi, I.; Xia, X.-Z.; McCabe, S.; Miner, K.; Solovyev, I.; Hawkins, N.; Kelley, M.; Chang, D.; Van, G.; et al. Severe B Cell Hyperplasia and Autoimmune Disease in TALL-1 Transgenic Mice. Proc. Natl. Acad. Sci. USA 2000, 97, 3370–3375. [Google Scholar] [CrossRef]
- Morimoto, A.; Omachi, S.; Osada, Y.; Chambers, J.K.; Uchida, K.; Sanjoba, C.; Matsumoto, Y.; Goto, Y. Hemophagocytosis in Experimental Visceral Leishmaniasis by Leishmania Donovani. PLoS Negl. Trop. Dis. 2016, 10, e0004505. [Google Scholar] [CrossRef]
- Omachi, S.; Fujii, W.; Azuma, N.; Morimoto, A.; Sanjoba, C.; Matsumoto, Y.; Goto, Y. B-Cell Activating Factor Deficiency Suppresses Splenomegaly during Leishmania Donovani Infection. Biochem. Biophys. Res. Commun. 2017, 489, 528–533. [Google Scholar] [CrossRef]
- Goto, Y.; Sanjoba, C.; Omachi, S.; Matsumoto, Y. Elevation of Serum B-Cell Activating Factor Levels During Visceral Leishmaniasis. Am. J. Trop. Med. Hyg. 2014, 91, 912–914. [Google Scholar] [CrossRef]
- Xiong, W.; Lahita, R.G. Pragmatic Approaches to Therapy for Systemic Lupus Erythematosus. Nat. Rev. Rheumatol. 2014, 10, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Stohl, W. Inhibition of B Cell Activating Factor (BAFF) in the Management of Systemic Lupus Erythematosus (SLE). Expert. Rev. Clin. Immunol. 2017, 13, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Allman, W.R.; Dey, R.; Liu, L.; Siddiqui, S.; Coleman, A.S.; Bhattacharya, P.; Yano, M.; Uslu, K.; Takeda, K.; Nakhasi, H.L.; et al. TACI Deficiency Leads to Alternatively Activated Macrophage Phenotype and Susceptibility to Leishmania Infection. Proc. Natl. Acad. Sci. USA 2015, 112, E4094–E4103. [Google Scholar] [CrossRef] [PubMed]
- Klinman, D.M.; Shirai, A.; Ishigatsubo, Y.; Conover, J.; Steinberg, A.D. Quantitation of IgM- and IgG-secreting B Cells in the Peripheral Blood of Patients with Systemic Lupus Erythematosus. Arthritis Rheumatol. 1991, 34, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Takahashi, Y.; Ogasawara, M.; Setoyama, Y.; Suzuki, K.; Tsuzaka, K.; Abe, T.; Takeuchi, T. Aberrant Expression of BAFF in T Cells of Systemic Lupus Erythematosus, Which Is Recapitulated by a Human T Cell Line, Loucy. Int. Immunol. 2006, 18, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Nakano, S.; Watanabe, T.; Tamayama, Y.; Mitsuo, A.; Nakiri, Y.; Suzuki, J.; Nozawa, K.; Amano, H.; Tokano, Y.; et al. Expression of B-Cell Activating Factor of the Tumour Necrosis Factor Family (BAFF) in T Cells in Active Systemic Lupus Erythematosus: The Role of BAFF in T Cell-Dependent B Cell Pathogenic Autoantibody Production. Rheumatology 2007, 46, 1083–1086. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Kuley, R.; Draves, K.E.; Elkon, K.B.; Giltiay, N.V.; Clark, E.A. B Cell-Activating Factor (BAFF) from Dendritic Cells, Monocytes and Neutrophils Is Required for B Cell Maturation and Autoantibody Production in SLE-like Autoimmune Disease. Front. Immunol. 2023, 14, 1050528. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Itoh, K.; Nagatani, K.; Okawa-Takatsuji, M.; Fujii, T.; Kuroki, H.; Katsuragawa, Y.; Aotsuka, S.; Mimori, A. Expression of BAFF and BAFF-R in the Synovial Tissue of Patients with Rheumatoid Arthritis. Scand. J. Rheumatol. 2007, 36, 365–372. [Google Scholar] [CrossRef]
- Lavie, F.; Miceli-Richard, C.; Quillard, J.; Roux, S.; Leclerc, P.; Mariette, X. Expression of BAFF (BLyS) in T Cells Infiltrating Labial Salivary Glands from Patients with Sjögren’s Syndrome. J. Pathol. 2004, 202, 496–502. [Google Scholar] [CrossRef]
- Lokensgard, J.R.; Mutnal, M.B.; Prasad, S.; Sheng, W.; Hu, S. Glial Cell Activation, Recruitment, and Survival of B-Lineage Cells Following MCMV Brain Infection. J. Neuroinflamm. 2016, 13, 114. [Google Scholar] [CrossRef]
- Phares, T.W.; Marques, C.P.; Stohlman, S.A.; Hinton, D.R.; Bergmann, C.C. Factors Supporting Intrathecal Humoral Responses Following Viral Encephalomyelitis. J. Virol. 2011, 85, 2589–2598. [Google Scholar] [CrossRef]
- Triantafilou, K.; Kar, S.; Vakakis, E.; Kotecha, S.; Triantafilou, M. Human Respiratory Syncytial Virus Viroporin SH: A Viral Recognition Pathway Used by the Host to Signal Inflammasome Activation. Thorax 2013, 68, 66–75. [Google Scholar] [CrossRef]
- Kuley, R.; Draves, K.E.; Fuller, D.H.; Giltiay, N.V.; Clark, E.A.; Giordano, D. B Cell Activating Factor (BAFF) from Neutrophils and Dendritic Cells Is Required for Protective B Cell Responses against Salmonella Typhimurium Infection. PLoS ONE 2021, 16, e0259158. [Google Scholar] [CrossRef]
- Pandey, K.; Yanagi, T.; Pandey, B.D.; Mallik, A.K.; Sherchand, J.B.; Kanbara, H. Characterization of Leishmania Isolates from Nepalese Patients with Visceral Leishmaniasis. Parasitol. Res. 2007, 100, 1361–1369. [Google Scholar] [CrossRef]
- Webb, J.R.; Campos-Neto, A.; Ovendale, P.J.; Martin, T.I.; Stromberg, E.J.; Badaro, R.; Reed, S.G. Human and Murine Immune Responses to a Novel Leishmania Major. Recombinant Protein Encoded by Members of a Multicopy Gene Family. Infect. Immun. 1998, 66, 3279–3289. [Google Scholar] [CrossRef]
- Goto, Y.; Carter, D.; Guderian, J.; Inoue, N.; Kawazu, S.-I.; Reed, S.G. Upregulated Expression of B-Cell Antigen Family Tandem Repeat Proteins by Leishmania Amastigotes. Infect. Immun. 2010, 78, 2138–2145. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, K.; Fujii, W.; Azuma, N.; Mizobuchi, H.; Morimoto, A.; Sanjoba, C.; Matsumoto, Y.; Goto, Y. Pathological Roles of MRP14 in Anemia and Splenomegaly during Experimental Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2020, 14, e0008020. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.S.; Schneider, P.; Kalled, S.L.; Wang, L.; Lefevre, E.A.; Cachero, T.G.; MacKay, F.; Bixler, S.A.; Zafari, M.; Liu, Z.-Y.; et al. Baff Binds to the Tumor Necrosis Factor Receptor–Like Molecule B Cell Maturation Antigen and Is Important for Maintaining the Peripheral B Cell Population. J. Exp. Med. 2000, 192, 129–136. [Google Scholar] [CrossRef]
- Schiemann, B.; Gommerman, J.L.; Vora, K.; Cachero, T.G.; Shulga-Morskaya, S.; Dobles, M.; Frew, E.; Scott, M.L. An Essential Role for BAFF in the Normal Development of B Cells Through a BCMA-Independent Pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Boone, T.; Delaney, J.; Hawkins, N.; Kelley, M.; Ramakrishnan, M.; McCabe, S.; Qiu, W.; Kornuc, M.; Xia, X.-Z.; et al. APRIL and TALL-1 and Receptors BCMA and TACI: System for Regulating Humoral Immunity. Nat. Immunol. 2000, 1, 252–256. [Google Scholar] [CrossRef]
- Yan, M.; Marsters, S.A.; Grewal, I.S.; Wang, H.; Ashkenazi, A.; Dixit, V.M. Identification of a Receptor for BLyS Demonstrates a Crucial Role in Humoral Immunity. Nat. Immunol. 2000, 1, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Mantchev, G.T.; Cortesão, C.S.; Rebrovich, M.; Cascalho, M.; Bram, R.J. TACI Is Required for Efficient Plasma Cell Differentiation in Response to T-Independent Type 2 Antigens. J. Immunol. 2007, 179, 2282–2288. [Google Scholar] [CrossRef]
- Cremasco, V.; Woodruff, M.C.; Onder, L.; Cupovic, J.; Nieves-Bonilla, J.M.; Schildberg, F.A.; Chang, J.; Cremasco, F.; Harvey, C.J.; Wucherpfennig, K.; et al. B Cell Homeostasis and Follicle Confines Are Governed by Fibroblastic Reticular Cells. Nat. Immunol. 2014, 15, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, L.; Gilbride, K.; Dobles, M.; Kalled, S.L.; Zandman, D.; Scott, M.L. Normal B Cell Homeostasis Requires B Cell Activation Factor Production by Radiation-Resistant Cells. J. Exp. Med. 2003, 198, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cho, B.; Suzuki, K.; Xu, Y.; Green, J.A.; An, J.; Cyster, J.G. Follicular Dendritic Cells Help Establish Follicle Identity and Promote B Cell Retention in Germinal Centers. J. Exp. Med. 2011, 208, 2497–2510. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Kuley, R.; Draves, K.E.; Roe, K.; Holder, U.; Giltiay, N.V.; Clark, E.A. BAFF Produced by Neutrophils and Dendritic Cells Is Regulated Differently and Has Distinct Roles in Antibody Responses and Protective Immunity against West Nile Virus. J. Immunol. 2020, 204, 1508–1520. [Google Scholar] [CrossRef] [PubMed]
- Ryckman, C.; Vandal, K.; Rouleau, P.; Talbot, M.; Tessier, P.A. Proinflammatory Activities of S100: Proteins S100A8, S100A9, and S100A8/A9 Induce Neutrophil Chemotaxis and Adhesion. J. Immunol. 2003, 170, 3233–3242. [Google Scholar] [CrossRef]
- Foell, D.; Frosch, M.; Sorg, C.; Roth, J. Phagocyte-Specific Calcium-Binding S100 Proteins as Clinical Laboratory Markers of Inflammation. Clin. Chim. Acta 2004, 344, 37–51. [Google Scholar] [CrossRef]
- Ehrchen, J.M.; Sunderkötter, C.; Foell, D.; Vogl, T.; Roth, J. The Endogenous Toll–like Receptor 4 Agonist S100A8/S100A9 (Calprotectin) as Innate Amplifier of Infection, Autoimmunity, and Cancer. J. Leukoc. Biol. 2009, 86, 557–566. [Google Scholar] [CrossRef]
- Abram, C.L.; Roberge, G.L.; Hu, Y.; Lowell, C.A. Comparative Analysis of the Efficiency and Specificity of Myeloid-Cre Deleting Strains Using ROSA-EYFP Reporter Mice. J. Immunol. Methods 2014, 408, 89–100. [Google Scholar] [CrossRef]
- Goto, Y.; Sanjoba, C.; Arakaki, N.; Okamoto, M.; Saeki, K.; Onodera, T.; Ito, M.; Matsumoto, Y. Accumulation of Macrophages Expressing MRP8 and MRP14 in Skin Lesions during Leishmania Major Infection in BALB/c and RAG-2 Knockout Mice. Parasitol. Int. 2007, 56, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Yurdakul, P.; Dalton, J.; Beattie, L.; Brown, N.; Erguven, S.; Maroof, A.; Kaye, P.M. Compartment-Specific Remodeling of Splenic Micro-Architecture during Experimental Visceral Leishmaniasis. Am. J. Pathol. 2011, 179, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Terrazas, C.; Varikuti, S.; Oghumu, S.; Steinkamp, H.M.; Ardic, N.; Kimble, J.; Nakhasi, H.; Satoskar, A.R. Ly6Chi Inflammatory Monocytes Promote Susceptibility to Leishmania Donovani Infection. Sci. Rep. 2017, 7, 14693. [Google Scholar] [CrossRef] [PubMed]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood Monocytes Consist of Two Principal Subsets with Distinct Migratory Properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Tenbrock, K.; Ludwig, S.; Leukert, N.; Ehrhardt, C.; van Zoelen, M.A.D.; Nacken, W.; Foell, D.; van der Poll, T.; Sorg, C.; et al. Mrp8 and Mrp14 Are Endogenous Activators of Toll-like Receptor 4, Promoting Lethal, Endotoxin-Induced Shock. Nat. Med. 2007, 13, 1042–1049. [Google Scholar] [CrossRef]
- Loser, K.; Vogl, T.; Voskort, M.; Lueken, A.; Kupas, V.; Nacken, W.; Klenner, L.; Kuhn, A.; Foell, D.; Sorokin, L.; et al. The Toll-like Receptor 4 Ligands Mrp8 and Mrp14 Are Crucial in the Development of Autoreactive CD8+ T Cells. Nat. Med. 2010, 16, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Mizobuchi, H.; Fujii, W.; Isokawa, S.; Ishizuka, K.; Wang, Y.; Watanabe, S.; Sanjoba, C.; Matsumoto, Y.; Goto, Y. Exacerbation of Hepatic Injury during Rodent Malaria by Myeloid-Related Protein 14. PLoS ONE 2018, 13, e0199111. [Google Scholar] [CrossRef]
- Riva, M.; Källberg, E.; Björk, P.; Hancz, D.; Vogl, T.; Roth, J.; Ivars, F.; Leanderson, T. Induction of Nuclear Factor-κB Responses by the S100A9 Protein Is Toll-like Receptor-4-dependent. Immunology 2012, 137, 172–182. [Google Scholar] [CrossRef]
- Gardam, S.; Brink, R. Non-Canonical NF-ΚB Signaling Initiated by BAFF Influences B Cell Biology at Multiple Junctures. Front. Immunol. 2014, 4, 509. [Google Scholar] [CrossRef]
- Groom, J.R.; Fletcher, C.A.; Walters, S.N.; Grey, S.T.; Watt, S.V.; Sweet, M.J.; Smyth, M.J.; Mackay, C.R.; Mackay, F. BAFF and MyD88 Signals Promote a Lupuslike Disease Independent of T Cells. J. Exp. Med. 2007, 204, 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Luo, Z.; Yang, Q.; Chang, H.; Liu, P.; Li, Z.; Guo, S.; Zhou, C.; Song, J.; Cao, W. Neutrophil-Derived MRP14 Supports Plasma Cell Commitment and Protects Myeloma Cells from Apoptosis. J. Immunol. Res. 2019, 2019, 9561350. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagai, K.; Fujii, W.; Yamagishi, J.; Sanjoba, C.; Goto, Y. Inflammatory CD11b+ Macrophages Produce BAFF in Spleen of Mice Infected with Leishmania donovani. Pathogens 2024, 13, 232. https://doi.org/10.3390/pathogens13030232
Nagai K, Fujii W, Yamagishi J, Sanjoba C, Goto Y. Inflammatory CD11b+ Macrophages Produce BAFF in Spleen of Mice Infected with Leishmania donovani. Pathogens. 2024; 13(3):232. https://doi.org/10.3390/pathogens13030232
Chicago/Turabian StyleNagai, Kazuki, Wataru Fujii, Junya Yamagishi, Chizu Sanjoba, and Yasuyuki Goto. 2024. "Inflammatory CD11b+ Macrophages Produce BAFF in Spleen of Mice Infected with Leishmania donovani" Pathogens 13, no. 3: 232. https://doi.org/10.3390/pathogens13030232
APA StyleNagai, K., Fujii, W., Yamagishi, J., Sanjoba, C., & Goto, Y. (2024). Inflammatory CD11b+ Macrophages Produce BAFF in Spleen of Mice Infected with Leishmania donovani. Pathogens, 13(3), 232. https://doi.org/10.3390/pathogens13030232