
Community-Directed Vector Control to Accelerate Onchocerciasis Elimination


Abstract
:1. Introduction
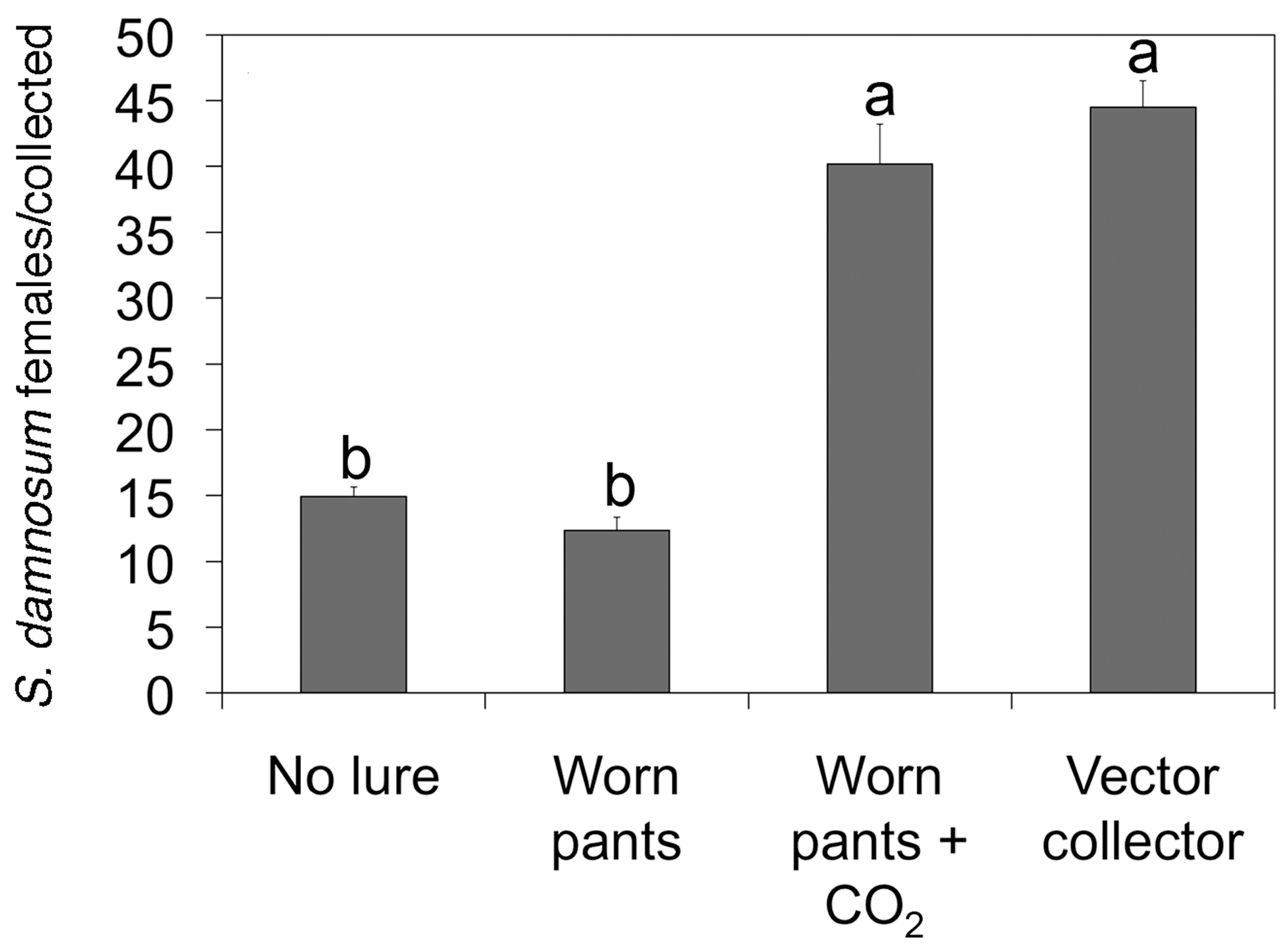
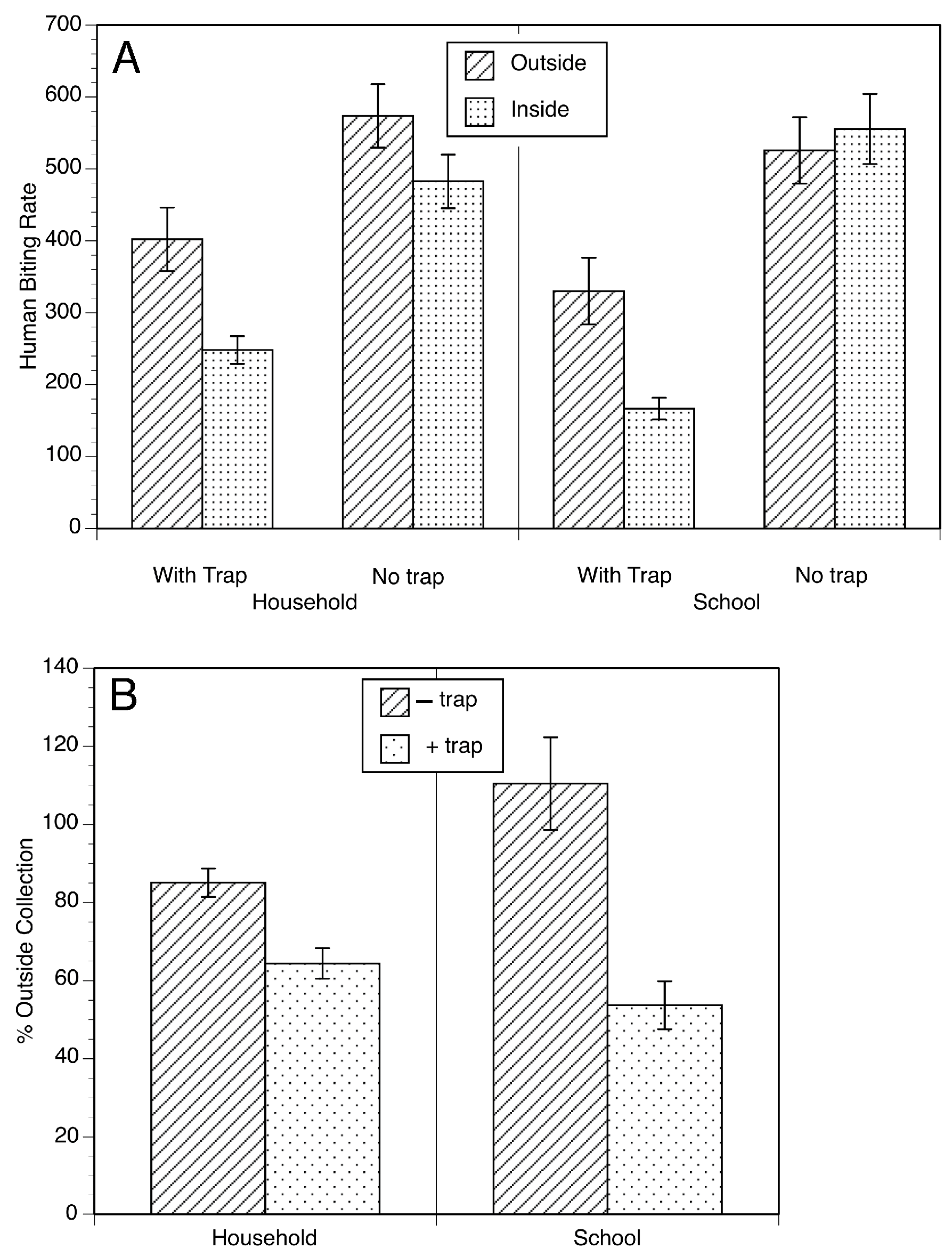
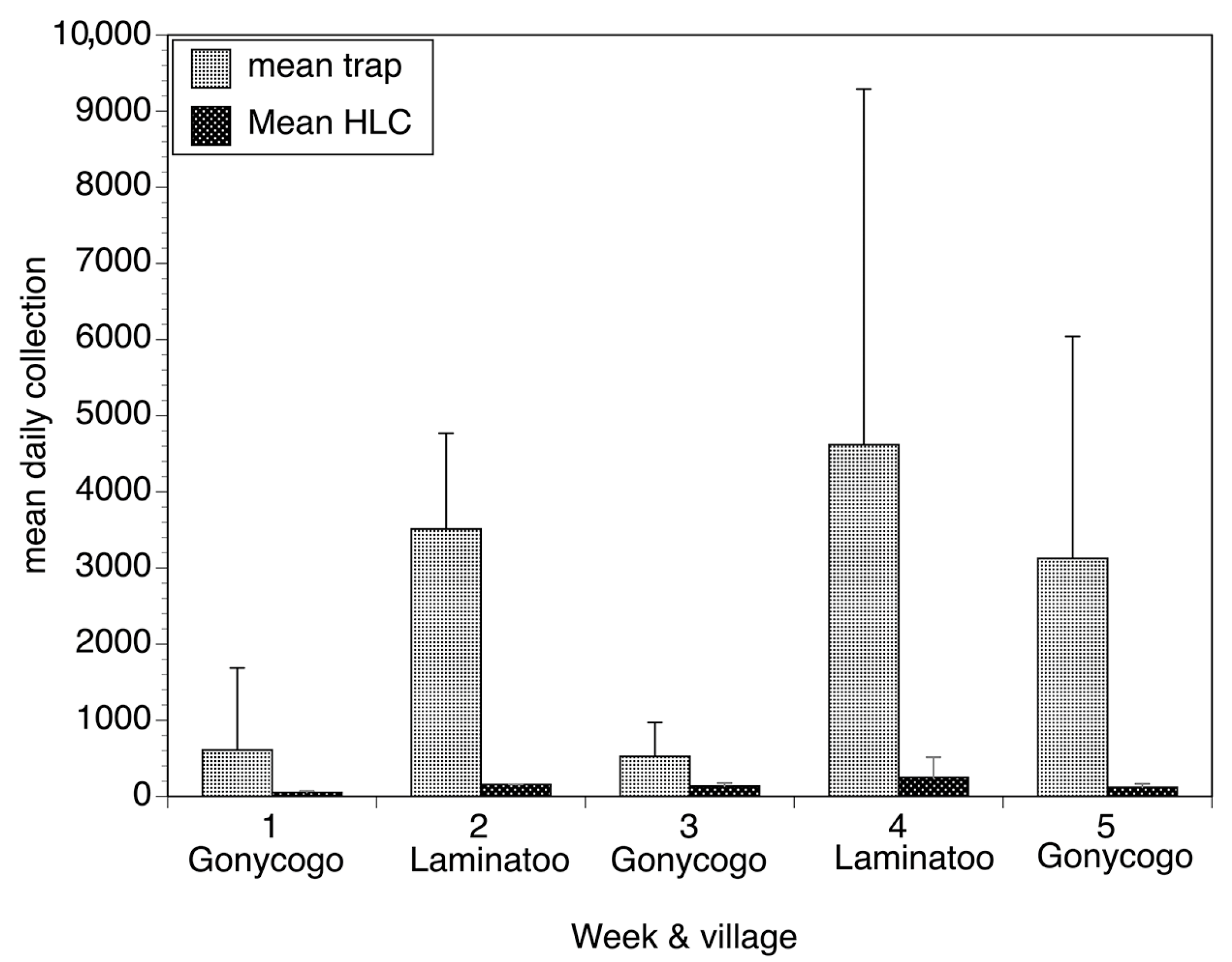
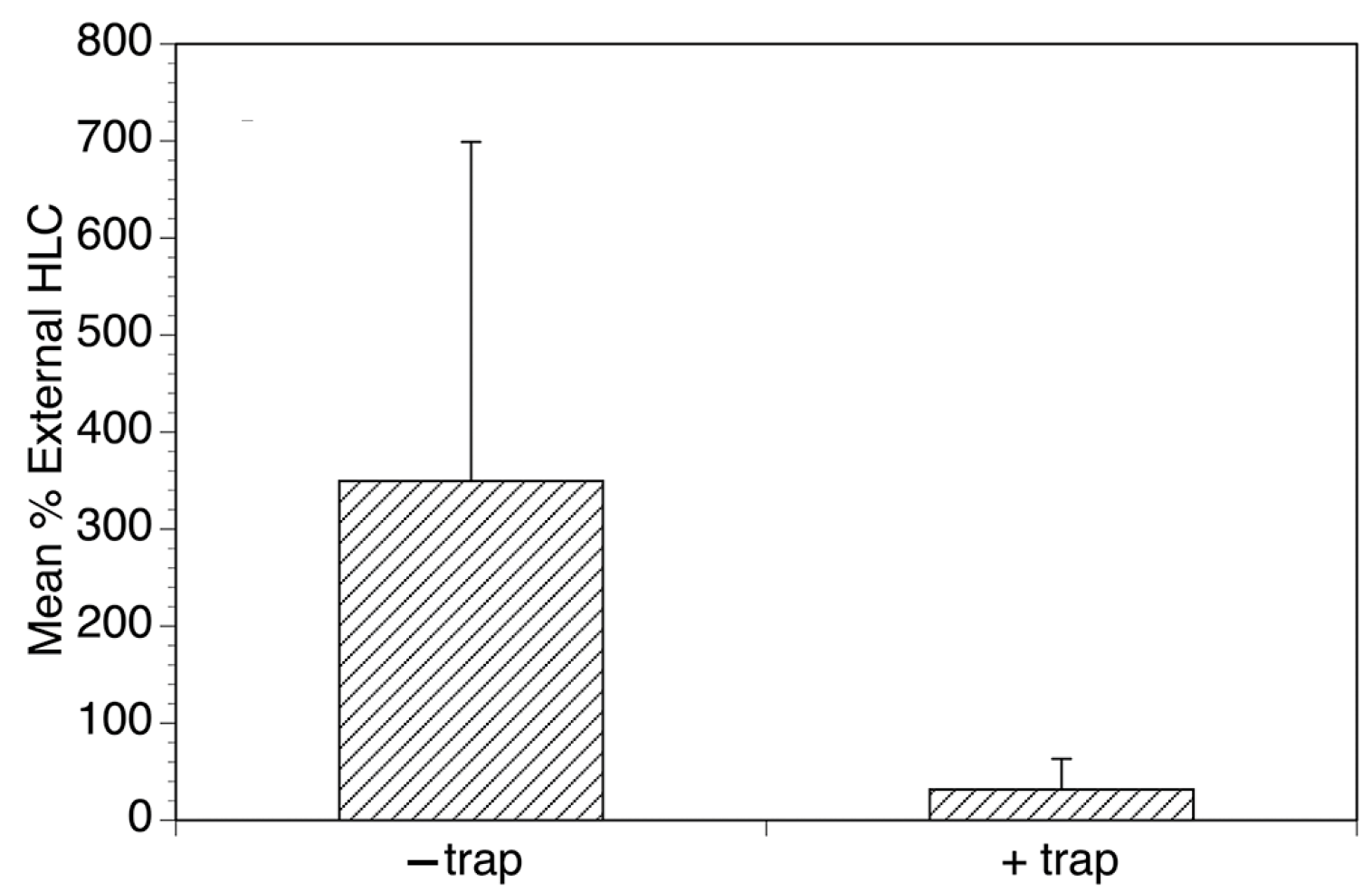
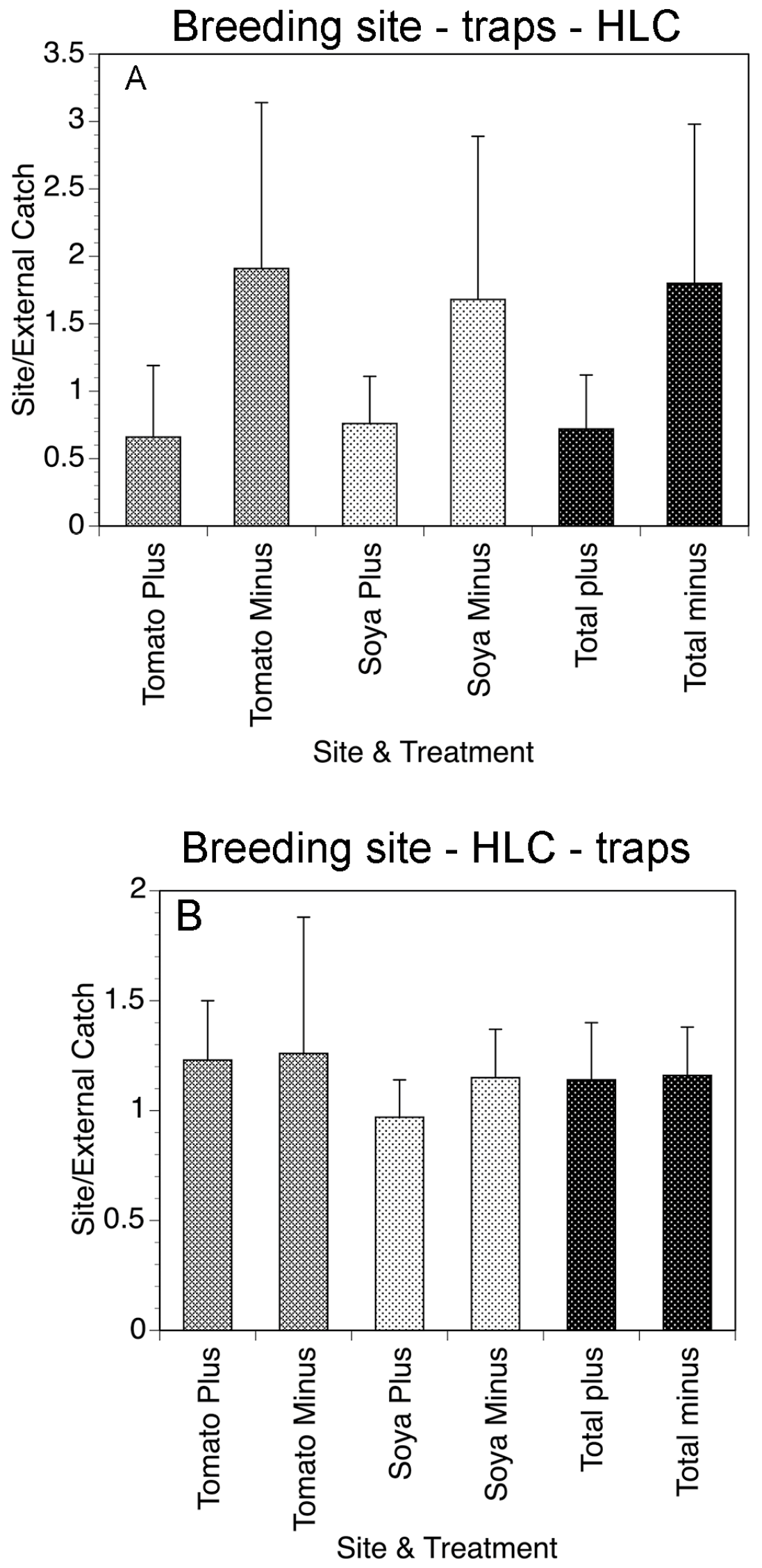
2. The Esperanza Window Trap (EWT)
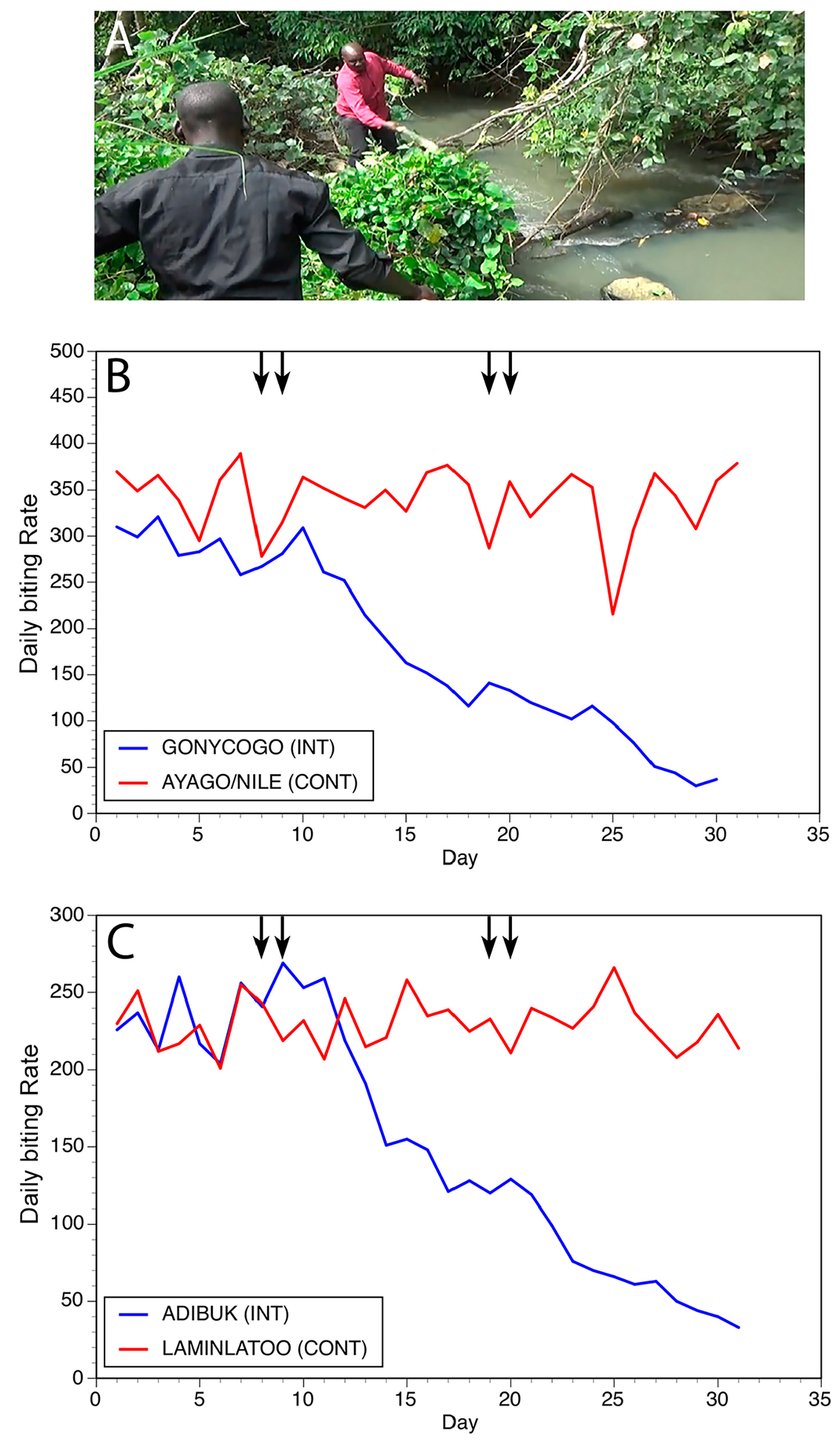
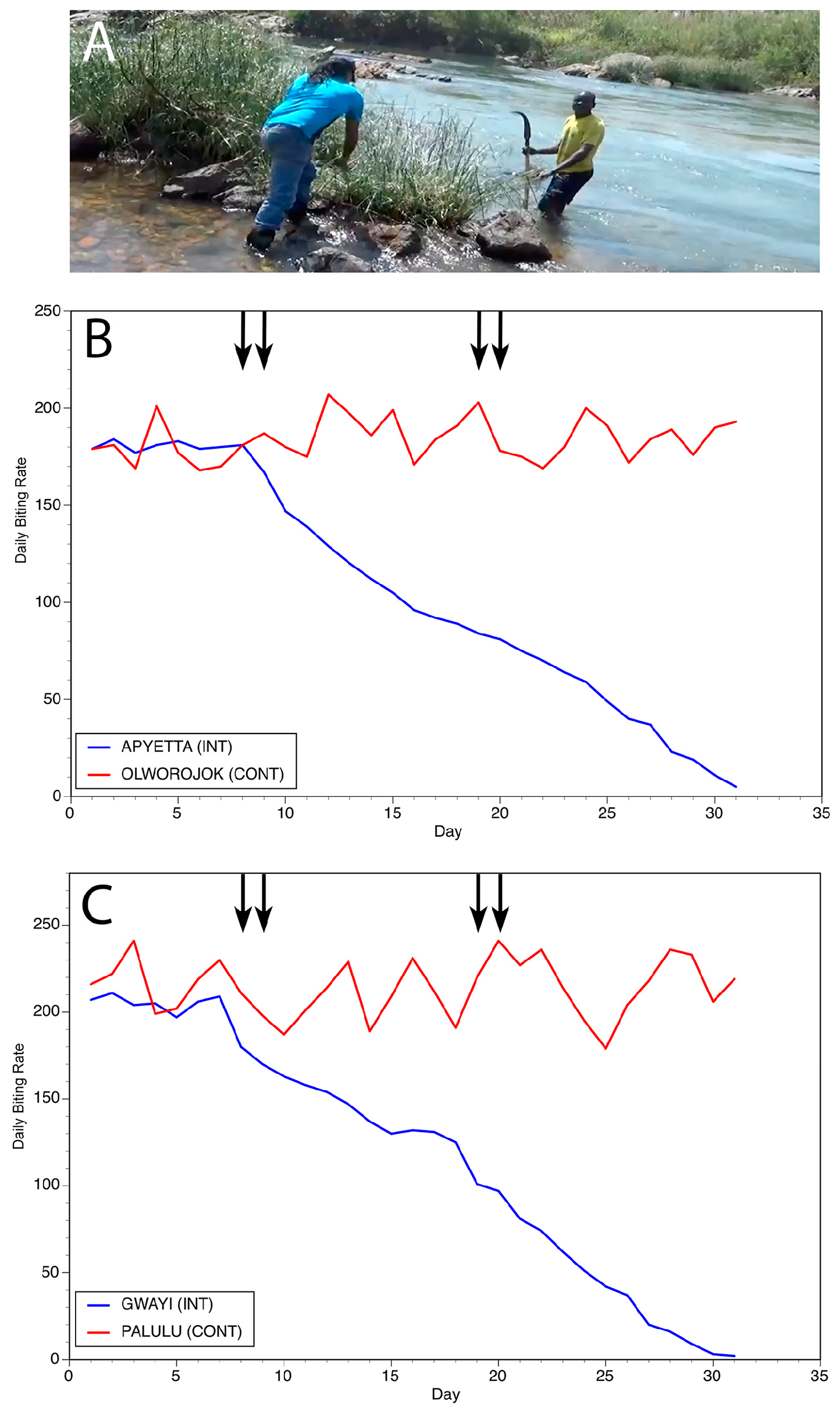
3. Slash and Clear
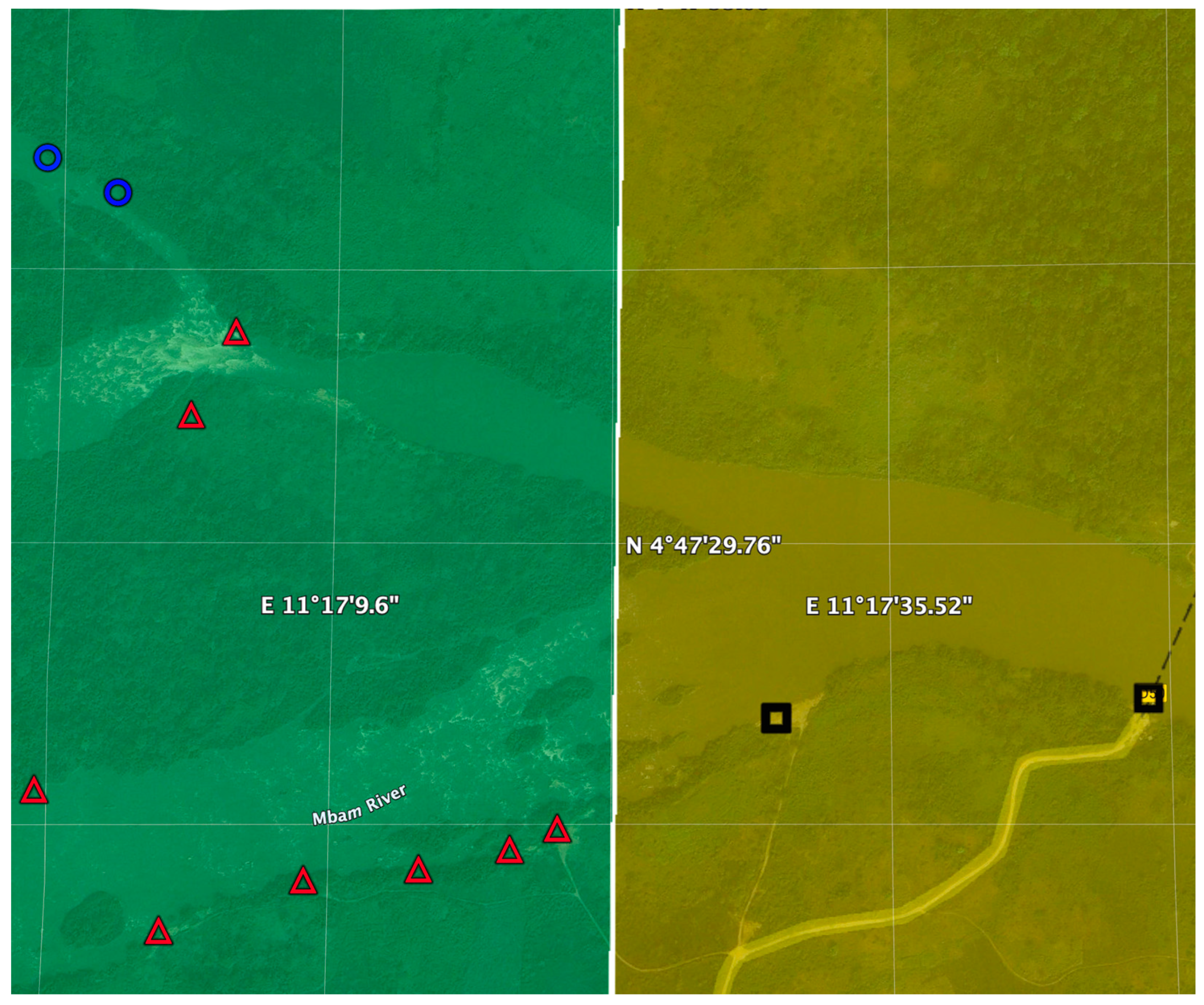
4. Breeding Site Identification
5. Modeling the Impact of Slash and Clear in Accelerating Elimination of Transmission
6. Research Challenges Remaining
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thylefors, B. Ocular Onchocerciasis. Bull. World Health Organ. 1978, 56, 63–72. [Google Scholar] [PubMed]
- World-Health-Organization-Expert-Committee. Onchocerciasis and Its Control; World Health Organization: Geneva, Switzerland, 1995; Volume 852. [Google Scholar]
- Opara, K.N.; Usip, L.P.; Akpabio, E.E. Transmission dynamics of Simulium damnosum in rural communities of Akwa Ibom State, Nigeria. J. Vector Borne Dis. 2008, 45, 225–230. [Google Scholar] [PubMed]
- Fischer, P.; Kipp, W.; Bamuhiga, J.; Bintakahwa, J.; Kiefer, A.; Buttner, D.W. Parasitological and Clinical Characterization of Simulium-neavei Transmitted Onchocerciasis in Western Uganda. Trop. Med. Parasitol. 1993, 44, 311–321. [Google Scholar] [PubMed]
- Greene, B.M.; Taylor, H.R.; Cupp, E.W.; Murphy, R.P.; White, A.T.; Aziz, M.A.; Schulz-Key, H.; D’Anna, S.A.; Newland, H.S.; Goldschmidt, L.P.; et al. Comparison of ivermectin and diethylcarbamazine in the treatment of onchocerciasis. N. Eng. J. Med. 1985, 313, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Cupp, E.W.; Bernardo, M.J.; Kiszewski, A.E.; Collins, R.C.; Taylor, H.R.; Aziz, M.A.; Greene, B.M. The effects of ivermectin on transmission of Onchocerca volvulus. Science 1986, 231, 740–742. [Google Scholar] [CrossRef]
- Cupp, E.W.; Ochoa, A.O.; Collins, R.C.; Ramberg, F.R.; Zea, G. The effect of multiple ivermectin treatments on infection of Simulium ochraceum with Onchocerca volvulus. Am. J. Trop. Med. Hyg. 1989, 40, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Colatrella, B. The Mectizan Donation Program: 20 years of successful collaboration—A retrospective. Ann. Trop. Med. Parasitol. 2008, 102 (Suppl. 1), 7–11. [Google Scholar] [CrossRef]
- The Carter Center. Onchocerciasis Elimination Program for the Americas. Available online: http://www.cartercenter.org/health/river_blindness/oepa.html (accessed on 7 May 2015).
- Traore, M.O.; Sarr, M.D.; Badji, A.; Bissan, Y.; Diawara, L.; Doumbia, K.; Goita, S.F.; Konate, L.; Mounkoro, K.; Seck, A.F.; et al. Proof-of-principle of onchocerciasis elimination with ivermectin treatment in endemic foci in Africa: Final results of a study in Mali and Senegal. PLoS Negl. Trop. Dis. 2012, 6, e1825. [Google Scholar] [CrossRef]
- Tekle, A.H.; Elhassan, E.; Isiyaku, S.; Amazigo, U.V.; Bush, S.; Noma, M.; Cousens, S.; Abiose, A.; Remme, J.H. Impact of long-term treatment of onchocerciasis with ivermectin in Kaduna State, Nigeria: First evidence of the potential for elimination in the operational area of the African Programme for Onchocerciasis Control. Parasites Vectors 2012, 5, 28. [Google Scholar] [CrossRef]
- Turner, H.C.; Walker, M.; Churcher, T.S.; Osei-Atweneboana, M.Y.; Biritwum, N.K.; Hopkins, A.; Prichard, R.K.; Basanez, M.G. Reaching the london declaration on neglected tropical diseases goals for onchocerciasis: An economic evaluation of increasing the frequency of ivermectin treatment in Africa. Clin. Infect. Dis. 2014, 59, 923–932. [Google Scholar] [CrossRef]
- Duerr, H.P.; Raddatz, G.; Eichner, M. Control of onchocerciasis in Africa: Threshold shifts, breakpoints and rules for elimination. Int. J. Parasitol. 2011, 41, 581–589. [Google Scholar] [CrossRef]
- Katabarwa, M.N.; Eyamba, A.; Nwane, P.; Enyong, P.; Kamgno, J.; Kuete, T.; Yaya, S.; Aboutou, R.; Mukenge, L.; Kafando, C.; et al. Fifteen years of annual mass treatment of onchocerciasis with ivermectin have not interrupted transmission in the west region of Cameroon. J. Parastiology Res. 2013, 2013, 420928. [Google Scholar] [CrossRef]
- Katabarwa, M.N.; Lakwo, T.; Habomugisha, P.; Agunyo, S.; Byamukama, E.; Oguttu, D.; Tukesiga, E.; Unoba, D.; Dramuke, P.; Onapa, A.; et al. Transmission of Onchocerca volvulus Continues in Nyagak-Bondo Focus of Northwestern Uganda after 18 Years of a Single Dose of Annual Treatment with Ivermectin. Am. J. Trop. Med. Hyg. 2013, 89, 293–300. [Google Scholar] [CrossRef]
- McMahon, J.P.; Highton, R.B.; Goiny, H. The eradication of Simulium neavei from Kenya. Bull. World Health Organ. 1958, 19, 75–107. [Google Scholar]
- Roberts, J.M.; Neumann, E.; Gockel, C.W.; Highton, R.B. Onchocerciasis in Kenya 9, 11 and 18 years after elimination of the vector. Bull. World Health Organ. 1967, 37, 195–212. [Google Scholar]
- Hougard, J.M.; Alley, E.S.; Yameogo, L.; Dadzie, K.Y.; Boatin, B.A. Eliminating onchocerciasis after 14 years of vector control: A proved strategy. J. Infect. Dis. 2001, 184, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Vieta, F. River blindness. Protection for 54 cents a year. United Nations Chron. 1998, 1, 12–13. [Google Scholar]
- Ndyomugyenyi, R.; Tukesiga, E.; Buttner, D.W.; Garms, R. The impact of ivermectin treatment alone and when in parallel with Simulium neavei elimination on onchocerciasis in Uganda. Trop. Med. Int. Health 2004, 9, 882–886. [Google Scholar] [CrossRef] [PubMed]
- Katabarwa, M.; Lakwo, T.; Habomugisha, P.; Agunyo, S.; Byamukama, E.; Oguttu, D.; Ndyomugyenyi, R.; Tukesiga, E.; Ochieng, G.O.; Abwaimo, F.; et al. Transmission of Onchocerca volvulus by Simulium neavei in Mount Elgon focus of Eastern Uganda has been interrupted. Am. J. Trop. Med. Hyg. 2014, 90, 1159–1166. [Google Scholar] [CrossRef]
- World Health Organization. Onchocerciasis: Guidelines for Stopping Mass Drug Administration and Verifying Elimination of Human Onchocerciasis; Document Number WHO/HTM/NTD/PCT/2016.1; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Rodríguez-Pérez, M.A.; Adeleke, M.A.; Burkett-Cadena, N.D.; Garza-Hernández, J.A.; Reyes-Villanueva, F.; Cupp, E.W.; Toé, L.; Salinas-Carmona, M.C.; Rodríguez-Ramírez, A.D.; Katholi, C.R.; et al. Development of a novel trap for the collection of black flies of the Simulium ochraceum complex. PLoS ONE 2013, 8, e76814. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, M.A.; Adeleke, M.A.; Rodríguez-Luna, I.C.; Cupp, E.W.; Unnasch, T.R. Evaluation of a community-based trapping program to collect Simulium ochraceum sensu lato for verification of onchocerciasis elimination. PLoS Negl. Trop. Dis. 2014, 8, e3249. [Google Scholar] [CrossRef] [PubMed]
- Toé, L.D.; Koala, L.; Burkett-Cadena, N.D.; Traoré, B.M.; Sanfo, M.; Kambiré, S.R.; Cupp, E.W.; Traoré, S.; Yameogo, L.; Boakye, D.; et al. Optimization of the Esperanza Window Trap for the collection of the African onchocerciasis vector Simulium damnosum sensu lato. Acta Trop. 2014, 137, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Perez, M.A.; Garza-Hernandez, J.A.; Salinas-Carmona, M.C.; Fernandez-Salas, I.; Reyes-Villanueva, F.; Real-Najarro, O.; Cupp, E.W.; Unnasch, T.R. The Esperanza Window Trap reduces the human biting rate of Simulium ochraceum s.l. in formerly onchocerciasis endemic foci in Southern Mexico. PLoS Negl. Trop. Dis. 2017, 11, e0005686. [Google Scholar] [CrossRef] [PubMed]
- Loum, D.; Cozart, D.; Lakwo, T.; Habomugisha, P.; Jacob, B.; Cupp, E.W.; Unnasch, T.R. Optimization and Evaluation of the Esperanza Window Trap to Reduce Biting Rates of Simulium damnosum sensu lato in Northern Uganda. PLoS Negl. Trop. Dis. 2019, 13, e0007558. [Google Scholar] [CrossRef] [PubMed]
- Jacob, B.G.; Loum, D.; Lakwo, T.L.; Katholi, C.R.; Habomugisha, P.; Byamukama, E.; Tukahebwa, E.; Cupp, E.W.; Unnasch, T.R. Community-Directed Vector Control to Supplement Mass Drug Distribution for Onchocerciasis Elimination in the Madi mid-North Focus of Northern Uganda. PLoS Negl. Trop. Dis. 2018, 12, e0006702. [Google Scholar] [CrossRef]
- Jacob, B.; Loum, D.; Munu, D.; Lakwo, T.; Byamukama, E.; Habomugisha, P.; Cupp, E.W.; Unnasch, T.R. Optimization of Slash and Clear Community Directed Control of Simulium damnosum sensu stricto in Northern Uganda. Am. J. Trop. Med. Hyg. 2021, 104, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Raimon, S.; Lakwo, T.L.; Sebit, W.J.; Siewe Fodjo, J.N.; Alinda, P.; Carter, J.Y.; Post, R.J.; Colebunders, R. “Slash and Clear”, a Community-Based Vector Control Method to Reduce Onchocerciasis Transmission by Simulium sirbanum in Maridi, South Sudan: A Prospective Study. Pathogens 2021, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Jacob, B.G.; Novak, R.J.; Toe, L.; Sanfo, M.S.; Afriyie, A.; Ibrahim, M.A.; Griffith, D.A.; Mutabakaru, E.; Unnasch, T.R. Quasi-likelihood techniques in a logistic regression equation for identifying Simulium damnosum s.l. larval habitats intra-cluster covariates in Togo. Geospat. Inf. Sci. 2012, 15, 117–133. [Google Scholar] [CrossRef]
- Du, Y.; Chang, C.-I.; Ren, H.; Chang, C.-C.; Jensen, J.; D’Amico, F. New hyperspectral discrimination measure for spectral characterization. Opt. Eng. 2004, 43, 1777–1786. [Google Scholar]
- Jacob, B.G.; Novak, R.J.; Toe, L.; Sanfo, M.; Griffith, D.A.; Lakwo, T.L.; Habomugisha, P.; Katabarwa, M.N.; Unnasch, T.R. Validation of a remote sensing model to identify Simulium damnosum s.l. breeding sites in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2013, 7, e2342. [Google Scholar] [CrossRef]
- Smith, M.E.; Bilal, S.; Lakwo, T.L.; Habomugisha, P.; Tukahebwa, E.; Byamukama, E.; Katabarwa, M.; Richards, F.O.; Cupp, E.W.; Unnasch, T.R.; et al. Accelerating river blindness elimination by supplementing MDA with a vegetation “slash and clear” vector control strategy: A data-driven modeling analysis. Sci. Rep. 2019, 9, 15274. [Google Scholar] [CrossRef] [PubMed]
- Michael, E.; Smith, M.E.; Katabarwa, M.N.; Byamukama, E.; Griswold, E.; Habomugisha, P.; Lakwo, T.; Tukahebwa, E.; Miri, E.S.; Eigege, A.; et al. Substantiating freedom from parasitic infection by combining transmission model predictions with disease surveys. Nat. Commun. 2018, 9, 4324. [Google Scholar] [CrossRef] [PubMed]
- Bockarie, M.J.; Pedersen, E.M.; White, G.B.; Michael, E. Role of vector control in the global program to eliminate lymphatic filariasis. Annu. Rev. Entomol. 2009, 54, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Michael, E.; Smith, M.E.; Singh, B.K.; Katabarwa, M.N.; Byamukama, E.; Habomugisha, P.; Lakwo, T.; Tukahebwa, E.; Richards, F.O. Data-driven modelling and spatial complexity supports heterogeneity-based integrative management for eliminating Simulium neavei-transmitted river blindness. Sci. Rep. 2020, 10, 4235. [Google Scholar] [CrossRef]
- Hendy, A.; Sluydts, V.; Tushar, T.; De Witte, J.; Odonga, P.; Loum, D.; Nyaraga, M.; Lakwo, T.; Dujardin, J.C.; Post, R.; et al. Esperanza Window Traps for the collection of anthropophilic blackflies (Diptera: Simuliidae) in Uganda and Tanzania. PLoS Negl. Trop. Dis. 2017, 11, e0005688. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
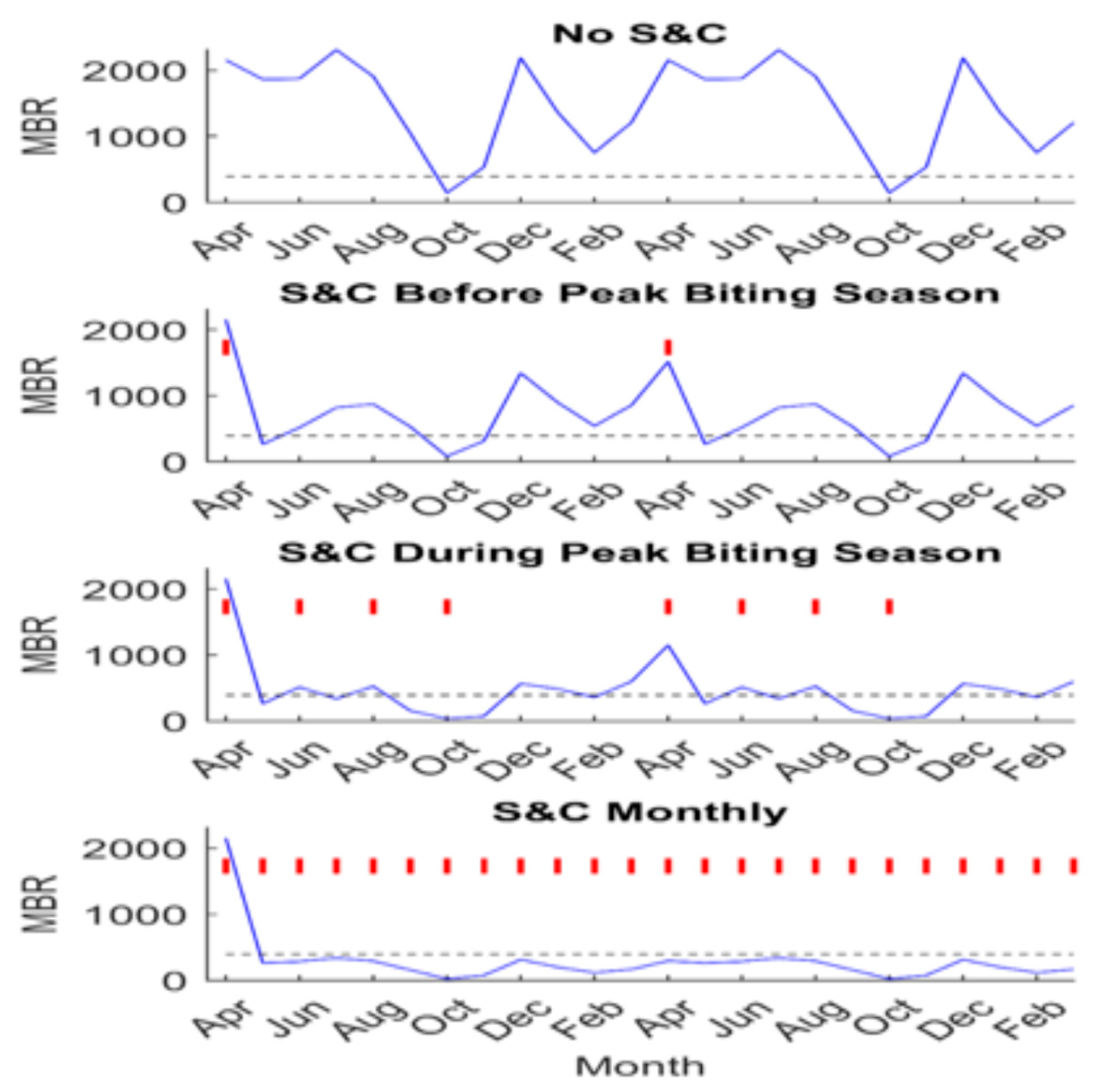{kind=link}
| Village (Baseline mf Prevalence (%), Model-Predicted mf Breakpoints at ABR and TBR (%)) | Skin mf Prevalence Threshold | ATP Threshold | ||||||
|---|---|---|---|---|---|---|---|---|
| No S&C | S&C before Peak Biting Season | S&C during Peak Biting Season | S&C Monthly | No S&C | S&C before Peak Biting Season | S&C during Peak Biting Season | S&C Monthly | |
| Model-predicted thresholds | ||||||||
| Palaure Pacunaci (100, 0.7, 0.08) | 34 (24–49) | 26 (16–45) | 25 (16–43) | 24 (16–41) | 28 (16–50) | 10 (2–23) | 8 (1–18) | 4 (1–12) |
| Masaloa (76, 0.9, 0.1) | 31 (19–49) | 19 (11–33) | 19 (10–31) | 18 (10–29) | 20 (10–34) | 7 (1–17) | 5 (1–14) | 1 (1–9) |
| Nyimanji (58, 0.8, 0.1) | 30 (18–47) | 19 (10–34) | 19 (10–33) | 18 (10–32) | 18 (8–33) | 7 (1–18) | 5 (1–14) | 1 (1–9) |
| Olimbuni/Aroga (24, 0.5, 0.1) | 28 (15–46) | 20 (9–38) | 19 (9–36) | 19 (9–34) | 17 (8–32) | 8 (1–18) | 5 (1–14) | 1 (1–9) |
| WHO thresholds | ||||||||
| Palaure Pacunaci (100, 0.7, 0.08) | 25 (15–45) | 24 (15–41) | 23 (15–40) | 22 (14–37) | 19 (9–45) | 16 (7–32) | 13 (4–26) | 9 (1–19) |
| Masaloa (76, 0.9, 0.1) | 20 (11–34) | 19 (10–32) | 19 (10–31) | 18 (10–29) | 13 (4–25) | 10 (1–21) | 8 (1–18) | 1 (1–12) |
| Nyimanji (58, 0.8, 0.1) | 19 (9–34) | 18 (9–31) | 17 (9–30) | 17 (9–29) | 11 (2–24) | 8 (1–20) | 6 (1–16) | 1 (1–10) |
| Olimbuni/Aroga (24, 0.5, 0.1) | 15 (5–30) | 14 (5–28) | 14 (5–26) | 14 (5–26) | 10 (1–22) | 7 (1–17) | 4 (1–13) | 1 (1–8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacob, B.; Michael, E.; Unnasch, T.R. Community-Directed Vector Control to Accelerate Onchocerciasis Elimination. Pathogens 2024, 13, 268. https://doi.org/10.3390/pathogens13030268
Jacob B, Michael E, Unnasch TR. Community-Directed Vector Control to Accelerate Onchocerciasis Elimination. Pathogens. 2024; 13(3):268. https://doi.org/10.3390/pathogens13030268
Chicago/Turabian StyleJacob, Benjamin, Edwin Michael, and Thomas R. Unnasch. 2024. "Community-Directed Vector Control to Accelerate Onchocerciasis Elimination" Pathogens 13, no. 3: 268. https://doi.org/10.3390/pathogens13030268
APA StyleJacob, B., Michael, E., & Unnasch, T. R. (2024). Community-Directed Vector Control to Accelerate Onchocerciasis Elimination. Pathogens, 13(3), 268. https://doi.org/10.3390/pathogens13030268