Morphine Attenuates Apically-Directed Cytokine Secretion from Intestinal Epithelial Cells in Response to Enteric Pathogens


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Morphine on Epithelial Immune Responses to Enteric Pathogens


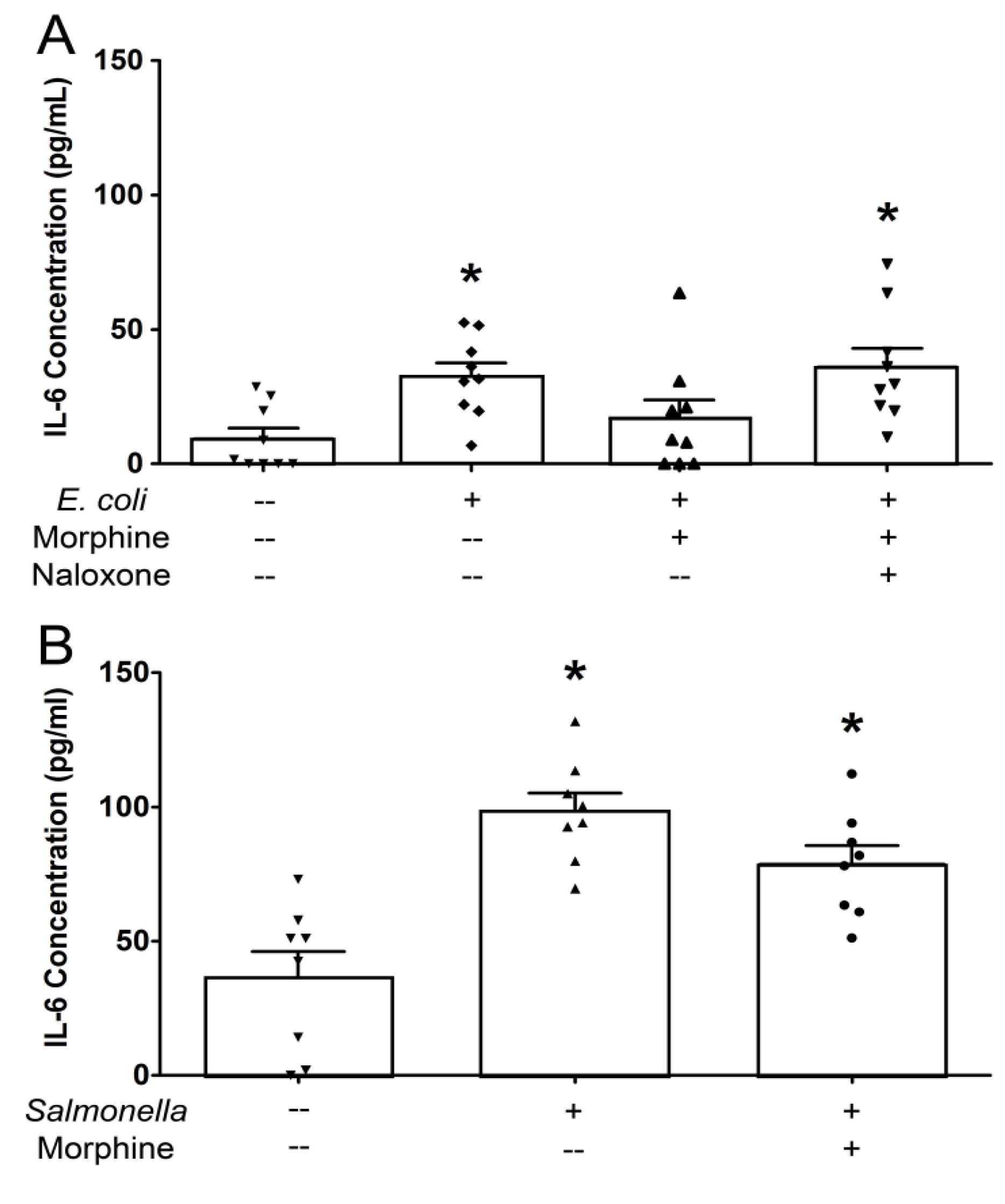{kind=link}
{kind=link}
| Control | E. coli | p value | Control | S. Typhimurium | p value | |
|---|---|---|---|---|---|---|
| Apical IL-8 concentration | 101.92 ± 4.54 | 425.60 ± 20.12 | 0.0015 | 143.72 ± 16.98 | 1129.74 ± 50.37 | <0.001 |
| Basolateral IL-8 concentration | 16.68 ± 2.78 | 1188.47 ± 45.02 | <0.001 | 48.14 ± 14.43 | 2516.64 ± 66.17 | <0.001 |
2.2. Effects of Morphine on Immune Responses by Epithelial Monolayers Exposed to Pathogen-Associated Molecular Patterns

3. Experimental Section
3.1. Reagents
3.2. IPEC-J2 Experiments
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Roy, S.; Ninkovic, J.; Banerjee, S.; Charboneau, R.G.; Das, S.; Dutta, R.; Kirchner, V.A.; Koodie, L.; Ma, J.; Meng, J.; et al. Opioid drug abuse and modulation of immune function: Consequences in the susceptibility to opportunistic infections. J. Neuroimmune Pharmacol. 2011, 6, 442–465. [Google Scholar] [CrossRef]
- Bidlack, J.M.; Khimich, M.; Parkhill, A.L.; Sumagin, S.; Sun, B.; Tipton, C.M. Opioid receptors and signaling on cells from the immune system. J. Neuroimmune Pharmacol. 2006, 1, 260–269. [Google Scholar] [CrossRef]
- Leslie, K.A.; Behme, R.; Clift, A.; Martin, S.; Grant, D.; Duff, J.H. Synergistic effects of tumour necrosis factor and morphine on gut barrier function. Can. J. Surg. 1994, 37, 143–147. [Google Scholar]
- Hilburger, M.E.; Adler, M.W.; Truant, A.L.; Meissler, J.J., Jr.; Satishchandran, V.; Rogers, T.J.; Eisenstein, T.K. Morphine induces sepsis in mice. J. Infect. Dis. 1997, 176, 183–188. [Google Scholar]
- Roy, S.; Charboneau, R.G.; Barke, R. A. Morphine synergizes with lipopolysaccharide in a chronic endotoxemia model. J. Neuroimmunol. 1999, 95, 107–114. [Google Scholar] [CrossRef]
- Ocasio, F.M.; Jiang, Y.; House, S.D.; Chang, S.L. Chronic morphine accelerates the progression of lipopolysaccharide-induced sepsis to septic shock. J. Neuroimmunol. 2004, 149, 90–100. [Google Scholar] [CrossRef]
- Runkel, N.S.; Moody, F.G.; Smith, G.S.; Rodriguez, L.F.; Chen, Y.; Larocco, M.T.; Miller, T.A. Alterations in rat intestinal transit by morphine promote bacterial translocation. Dig. Dis. Sci. 1993, 38, 1530–1536. [Google Scholar] [CrossRef]
- Asakura, H.; Kawamoto, K.; Igimi, S.; Yamamoto, S.; Makino, S. Enhancement of mice susceptibility to infection with Listeria monocytogenes by the treatment of morphine. Microbiol. Immunol. 2006, 50, 543–547. [Google Scholar] [CrossRef]
- Cray, W.C., Jr.; Tokunaga, E.; Pierce, N.F. Successful colonization and immunization of adult rabbits by oral inoculation with Vibrio cholerae O1. Infect. Immun. 1983, 41, 735–741. [Google Scholar]
- Babrowski, T.; Holbrook, C.; Moss, J.; Gottlieb, L.; Valuckaite, V.; Zaborin, A.; Poroyko, V.; Liu, D.C.; Zaborina, O.; Alverdy, J.C. Pseudomonas aeruginosa virulence expression is directly activated by morphine and is capable of causing lethal gut-derived sepsis in mice during chronic morphine administration. Ann. Surg. 2012, 255, 386–393. [Google Scholar] [CrossRef]
- MacFarlane, A.S.; Peng, X.; Meissler, J.J., Jr.; Rogers, T.J.; Geller, E.B.; Adler, M.W.; Eisenstein, T.K. Morphine increases susceptibility to oral Salmonella typhimurium infection. J. Infect. Dis. 2000, 181, 1350–1358. [Google Scholar] [CrossRef]
- Pott, J.; Hornef, M. Innate immune signalling at the intestinal epithelium in homeostasis and disease. EMBO Rep. 2012, 13, 684–698. [Google Scholar] [CrossRef]
- Holzer, P. Opioid receptors in the gastrointestinal tract. Regul. Pept. 2009, 155, 11–17. [Google Scholar] [CrossRef]
- Martinez-Maqueda, D.; Miralles, B.; de Pascual-Teresa, S.; Reveron, I.; Munoz, R.; Recio, I. Food-derived peptides stimulate mucin secretion and gene expression in intestinal cells. J. Agric. Food Chem. 2012, 60, 8600–8605. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, M.; Son, D.O. Food-derived peptides and intestinal functions. Curr. Pharm. Des. 2007, 13, 885–895. [Google Scholar] [CrossRef]
- Quito, F.L.; Seybold, V.S.; Brown, D.R. Opiate binding sites in mucosa of pig small intestine. Life Sci. 1991, 49, 219–222. [Google Scholar]
- Neudeck, B.L.; Loeb, J.; Buck, J. Activation of the kappa-opioid receptor in Caco-2 cells decreases interleukin-8 secretion. Eur. J. Pharmacol. 2003, 467, 81–84. [Google Scholar] [CrossRef]
- Neudeck, B.L.; Loeb, J.M. Endomorphin-1 alters interleukin-8 secretion in Caco-2 cells via a receptor mediated process. Immunol. Lett. 2002, 84, 217–221. [Google Scholar] [CrossRef]
- Zoghbi, S.; Trompette, A.; Claustre, J.; El Homsi, M.; Garzon, J.; Jourdan, G.; Scoazec, J.Y.; Plaisancie, P. Beta-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a mu-opioid pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef]
- Nano, J.L.; Fournel, S.; Rampal, P. Characterization of delta-opioid receptors and effect of enkephalins on IRD 98 rat epithelial intestinal cell line. Pflugers. Arch. 2000, 439, 547–554. [Google Scholar] [CrossRef]
- Lang, M.E.; Davison, J.S.; Bates, S.L.; Meddings, J.B. Opioid receptors on guinea-pig intestinal crypt epithelial cells. J. Physiol. 1996, 497, 161–174. [Google Scholar]
- Nylund, G.; Pettersson, A.; Bengtsson, C.; Khorram-Manesh, A.; Nordgren, S.; Delbro, D.S. Functional expression of mu-opioid receptors in the human colon cancer cell line, HT-29, and their localization in human colon. Dig. Dis. Sci. 2008, 53, 461–466. [Google Scholar] [CrossRef]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef]
- Skjolaas, K.A.; Burkey, T.E.; Dritz, S.S.; Minton, J.E. Effects of Salmonella enterica serovars Typhimurium (ST) and Choleraesuis (SC) on chemokine and cytokine expression in swine ileum and jejunal epithelial cells. Vet. Immunol. Immunopathol. 2006, 111, 199–209. [Google Scholar] [CrossRef]
- Skjolaas, K.A.; Burkey, T.E.; Dritz, S.S.; Minton, J.E. Effects of Salmonella enterica serovar Typhimurium, or serovar Choleraesuis, Lactobacillus reuteri and Bacillus licheniformis on chemokine and cytokine expression in the swine jejunal epithelial cell line, IPEC-J2. Vet. Immunol. Immunopathol. 2007, 115, 299–308. [Google Scholar] [CrossRef]
- Cao, J.; Ren, G.; Gong, Y.; Dong, S.; Yin, Y.; Zhang, L. Bronchial epithelial cells release IL-6, CXCL1 and CXCL8 upon mast cell interaction. Cytokine 2011, 56, 823–831. [Google Scholar] [CrossRef]
- Djalilian, A.R.; Nagineni, C.N.; Mahesh, S.P.; Smith, J.A.; Nussenblatt, R.B.; Hooks, J.J. Inhibition of inflammatory cytokine production in human corneal cells by dexamethasone, but not cyclosporin. Cornea 2006, 25, 709–714. [Google Scholar] [CrossRef]
- Levine, S.J.; Larivee, P.; Logun, C.; Angus, C.W.; Shelhamer, J.H. Corticosteroids differentially regulate secretion of IL-6, IL-8, and G-CSF by a human bronchial epithelial cell line. Am. J. Physiol. 1993, 265, L360–368. [Google Scholar]
- McCormick, B.A.; Colgan, S.P.; Delp-Archer, C.; Miller, S.I.; Madara, J.L. Salmonella Typhimurium attachment to human intestinal epithelial monolayers: Transcellular signalling to subepithelial neutrophils. .J. Cell Biol. 1993, 123, 895–907. [Google Scholar] [CrossRef]
- Stevens, C.W.; Aravind, S.; Das, S.; Davis, R.L. Pharmacological characterization of LPS and opioid interactions at the Toll-like receptor 4. Br. J. Pharmacol. 2013, 168, 1421–1429. [Google Scholar] [CrossRef]
- Watkins, L.R.; Hutchinson, M.R.; Rice, K.C.; Maier, S.F. The “toll” of opioid-induced glial activation: Improving the clinical efficacy of opioids by targeting glia. Trends Pharmacol. Sci. 2009, 30, 581–591. [Google Scholar] [CrossRef]
- Nadeau, W.J.; Pistole, T.G.; McCormick, B.A. Polymorphonuclear leukocyte migration across model intestinal epithelia enhances Salmonella typhimurium killing via the epithelial derived cytokine, IL-6. Microbes. Infect. 2002, 4, 1379–1387. [Google Scholar] [CrossRef]
- Sitaraman, S.V.; Merlin, D.; Wang, L.; Wong, M.; Gewirtz, A.T.; Si-Tahar, M.; Madara, J.L. Neutrophil-epithelial crosstalk at the intestinal lumenal surface mediated by reciprocal secretion of adenosine and IL-6. J. Clin. Invest. 2001, 107, 861–869. [Google Scholar] [CrossRef]
- Schmidt, L.D.; Kohrt, L.J.; Brown, D.R. Comparison of growth phase on Salmonella enterica serovar Typhimurium invasion in an epithelial cell line (IPEC J2) and mucosal explants from porcine small intestine. Comp Immunol. Microbiol. Infect. Dis. 2008, 31, 63–69. [Google Scholar] [CrossRef]
- Chen, C.; Lyte, M.; Stevens, M.P.; Vulchanova, L.; Brown, D.R. Mucosally-directed adrenergic nerves and sympathomimetic drugs enhance non-intimate adherence of Escherichia coli O157:H7 to porcine cecum and colon. Eur. J. Pharmacol. 2006, 539, 116–124. [Google Scholar] [CrossRef]
- Tzipori, S.; Karch, H.; Wachsmuth, K.I.; Robins-Browne, R.M.; O’Brien, A.D.; Lior, H.; Cohen, M.L.; Smithers, J.; Levine, M.M. Role of a 60-megadalton plasmid and Shiga-like toxins in the pathogenesis of infection caused by enterohemorrhagic Escherichia coli O157:H7 in gnotobiotic piglets. Infect. Immun. 1987, 55, 3117–3125. [Google Scholar]
- Arce, C.; Ramirez-Boo, M.; Lucena, C.; Garrido, J.J. Innate immune activation of swine intestinal epithelial cell lines (IPEC-J2 and IPI-2I) in response to LPS from Salmonella typhimurium. Comp Immunol. Microbiol. Infect. Dis. 2010, 33, 161–174. [Google Scholar] [CrossRef]
- Bannon, C.; Davies, P.J.; Collett, A.; Warhurst, G. Potentiation of flagellin responses in gut epithelial cells by interferon-gamma is associated with STAT-independent regulation of MyD88 expression. Biochem. J. 2009, 423, 119–128. [Google Scholar]
- Philippe, D.; Dubuquoy, L.; Groux, H.; Brun, V.; Chuoï-Mariot, M.T.; Gaveriaux-Ruff, C.; Colombel, J.F.; Kieffer, B.L.; Desreumaux, P. Anti-inflammatory properties of the mu opioid receptor support its use in the treatment of colon inflammation. J. Clin. Invest. 2003, 111, 1329–1338. [Google Scholar] [CrossRef]
- Goldsmith, J.R.; Uronis, J.M.; Jobin, C. Mu opioid signaling protects against acute murine intestinal injury in a manner involving Stat3 signaling. Am. J. Pathol. 2011, 179, 673–683. [Google Scholar] [CrossRef]
- Goldsmith, J.R.; Perez-Chanona, E.; Yadav, P.N.; Whistler, J.; Roth, B.; Jobin, C. Intestinal epithelial cell-derived μ-opioid signaling protects against ischemia reperfusion injury through PI3K signaling. Am. J. Pathol. 2013, 182, 776–785. [Google Scholar] [CrossRef]
- Mignat, C.; Wille, U.; Ziegler, A. Affinity profiles of morphine, codeine, dihydrocodeine and their glucuronides at opioid receptor subtypes. Life Sci. 1995, 56, 79–799. [Google Scholar]
- Roy, S.; Cain, K.J.; Chapin, R.B.; Charboneau, R.G.; Barke, R.A. Morphine modulates NF kappa B activation in macrophages. Biochem. Biophys. Res. Comm. 1998, 245, 392–396. [Google Scholar] [CrossRef]
- Welters, I.D.; Menzebach, A.; Goumon, Y.; Cadet, P.; Menges, T.; Hughes, T.K.; Hempelmann, G.; Stefano, G.B. Morphine inhibits NF-kappaB nuclear binding in human neutrophils and monocytes by a nitric oxide-dependent mechanism. Anesthesiology 2000, 92, 1677–1684. [Google Scholar] [CrossRef]
- Ye, R.D. Regulation of nuclear factor κB activation by G-protein-coupled receptors. J. Leukoc. Biol. 2001, 70, 839–848. [Google Scholar]
- Chen, Y.L.; Law, P.Y.; Loh, H.H. Nuclear factor kappaB signaling in opioid functions and receptor gene expression. J. Neuroimmune Pharmacol. 2006, 1, 270–279. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brosnahan, A.J.; Jones, B.J.; Dvorak, C.M.; Brown, D.R. Morphine Attenuates Apically-Directed Cytokine Secretion from Intestinal Epithelial Cells in Response to Enteric Pathogens. Pathogens 2014, 3, 249-257. https://doi.org/10.3390/pathogens3020249
Brosnahan AJ, Jones BJ, Dvorak CM, Brown DR. Morphine Attenuates Apically-Directed Cytokine Secretion from Intestinal Epithelial Cells in Response to Enteric Pathogens. Pathogens. 2014; 3(2):249-257. https://doi.org/10.3390/pathogens3020249
Chicago/Turabian StyleBrosnahan, Amanda J., Bryan J. Jones, Cheryl M. Dvorak, and David R. Brown. 2014. "Morphine Attenuates Apically-Directed Cytokine Secretion from Intestinal Epithelial Cells in Response to Enteric Pathogens" Pathogens 3, no. 2: 249-257. https://doi.org/10.3390/pathogens3020249
APA StyleBrosnahan, A. J., Jones, B. J., Dvorak, C. M., & Brown, D. R. (2014). Morphine Attenuates Apically-Directed Cytokine Secretion from Intestinal Epithelial Cells in Response to Enteric Pathogens. Pathogens, 3(2), 249-257. https://doi.org/10.3390/pathogens3020249